
Cercosaura hypnoides, Doan, Tiffany M. & Lamar, William W., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.208567 |
|
publication LSID |
lsid:zoobank.org:pub:40C56FA5-1827-4F8F-B236-FCE0D3AFAB53 |
|
DOI |
https://doi.org/10.5281/zenodo.5657707 |
|
persistent identifier |
https://treatment.plazi.org/id/0AF96CEF-8879-4BD9-AF02-5E23730048ED |
|
taxon LSID |
lsid:zoobank.org:act:0AF96CEF-8879-4BD9-AF02-5E23730048ED |
|
treatment provided by |
Plazi |
|
scientific name |
Cercosaura hypnoides |
| status |
sp. nov. |
Cercosaura hypnoides new species
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 .
Holotype. UTA R- 60249 ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), an adult male, from Vereda de Portachuelo, in the vicinity of Manzanares, Municipality of Acacías, Department of Meta, Colombia; 4.1°N, 73.8°W; 1640 m ( Fig. 4 View FIGURE 4 ); collected on 17 August 1979 by William W. Lamar and Eduardo Thierry.
Paratypes. All paratypes from Vereda de Portachuelo, 1640 m, collected by William W. Lamar and Eduardo Thierry. UTA R- 60250, was found as an egg that hatched upon touch on 0 8 July 1980. All five other paratypes were collected as eggs that subsequently hatched in captivity. UTA R- 60248 ( Fig. 3 View FIGURE 3 ), collected on 0 8 July 1979; UTA R- 60251–60254, collected on 0 9 July 1980.
Diagnosis. (1) supraoculars three; (2) superciliaries 4–5, first expanded onto dorsal surface of head; (3) palpebral eye-disc made up of a single, undivided scale; (4) supralabials five; (5) infralabials five; (6) dorsal body scales hexagonal, with high rounded keel; (7) dorsal scales in transverse and oblique rows; (8) transverse rows of dorsals 28–31; (9) a continuous series of small lateral scales separating dorsals from ventrals, 4–7 scales wide; (10) transverse ventral rows 16–20; (11) longitudinal ventral rows six; (12) femoral pores per hind limb 8–12; (13) preanal pores absent; (14) subdigital lamellae on Toe IV 18–22; (15) limbs overlapping when adpressed against body; (16) dorsolateral stripe passing through dorsal surface of the eye and lateral stripe beginning on the supralabials, no vertebral stripe.
Cercosaura hypnoides is most similar to the species that were formerly considered to belong to the genus Pantodactylus ( Ruibal, 1952) , but may be distinguished from each of them by having lateral scales that are much smaller than the dorsal scales (versus dorsals and laterals approximately equal in size in C. parkeri (Ruibal) , C. quadrilineata Boettger , C. schreibersii Wiegmann , and C. steyeri ). Cercosaura hypnoides may also be distinguished from C. quadrilineata by having six longitudinal rows of ventral scales (versus 4). Cercosaura hypnoides can be further distinguished from C. parkeri (Ruibal) by the high femoral pore count of 8–12 (versus 2–6 for C. parkeri ). The new species can be distinguished from C. schreibersii Wiegmann by having three postocular scales (versus two) and by having 8–12 femoral pores versus 3–5. Cercosaura hypnoides can be distinguished from C. steyeri by having six longitudinal rows of ventral scales (versus 4 in C. steyeri ).
The new species can be distinguished from additional species of the genus Cercosaura by possessing hexagonal dorsal scales arranged in transverse and oblique series (versus quadrangular dorsals in transverse and longitudinal series for C. ocellata Wagler and by having only transverse series in C. argulus Peters , C. dicra (Uzzell) , C. eigenmanni (Griffin) , C. manicata O’Shaughnessy , C. nigroventris (Gorzula and Señaris) , C. phelpsorum (Lancini) , and C. vertebralis O’Shaughnessy ). The new species can be further separated from C. dicra and C. vertebralis by having three supraocular scales (versus four). Cercosaura hypnoides can be distinguished from C. dicra (8), C. manicata (8), and C. nigroventris (8) by having six longitudinal rows of ventral scales. The new species additionally differs from C. argulus , C. eigenmanni , C. ocellata , C. phelpsorum and C. vertebralis in its pattern of dorsolateral and lateral stripes.
Description of holotype. Adult male, with hemipenes everted, snout-vent length (SVL) 38.60 mm, tail 48.12 mm, partially regenerated, length of regenerated portion 8.03 mm; head scales smooth, glossy, without striations or rugosities; rostral scale wider than tall, meeting supralabials on either side at above the height of supralabials, in contact with frontonasal, nasals, and first supralabials; frontonasal wider than long, pentagonal with all corners rounded, in contact with nasals, prefrontals, and in point contact with loreal, shorter than frontal; prefrontals paired, pentagonal, in contact with frontal, anteriormost supraoculars, loreals, and anteriormost superciliaries; frontal longer than wide, hexagonal, in contact with first two pairs of supraoculars and frontoparietals; frontoparietals hexagonal, in contact with second and third pairs of supraoculars, parietals and interparietal; supraoculars three, first supraocular in contact with first three superciliaries, second supraocular in contact with supraciliaries 3 and 4, third supraocular in contact with fourth supraciliary, one postocular, and parietal; interparietal longer than wide, heptagonal, in contact with parietals and occipitals; parietals hexagonal, in contact with one postocular, a subequally large supratemporal, and one occipital; occipitals three, smaller than parietals, median smallest, extending further posteriorly than two lateral occipitals. Nasal longer than high, nostril situated in anterior third of scale, in contact with first and second supralabials, loreal, and frenocular; loreal quadrangular, not in contact with supralabials, contacting frenocular and first superciliary; frenocular quadrangular, dorsalmost corner in contact with first superciliary, in contact with second and third supralabials, preocular, and first subocular; four superciliaries, first expanded onto dorsal surface of head; one preocular, in contact with first superciliary and first subocular; palpebral eye-disc made up of a single transparent scale; suboculars five; postoculars three; temporals smooth, glossy, polygonal; supratemporals two, first largest; ear opening oblong, tympanum recessed; supralabials five, first four supralabials to angle of jaw; infralabials five. Mental wider than long, in contact with first infralabial and postmental posteriorly; postmental single, pentagonal, posterior suture angular with point directed posteriorly, in contact with first and second infralabials and first pair of genials; two pairs of genials, anterior pair in contact with second and third infralabials, second genials in contact with third and fourth infralabials; two pairs of chin shields, separated by irregular pregulars; gular scale rows eight; collar fold indistinct; lateral neck scales round, smooth.
Dorsals hexagonal, longer than wide, with posterior margins slightly curved, imbricate, with single high rounded keel, in 30 transverse rows, oblique rows also present; some paravertebral scales irregularly arranged; longitudinal dorsal scale rows 10 at fifth transverse ventral scale row, 11 at tenth transverse ventral scale row, 10 at fifteenth transverse ventral scale row; wide continuous lateral scale series, five to six scales wide, approximately half the size of dorsals, rounded squarish, smaller and more numerous lateral scales at limb insertion regions; no lateral fold. Ventral scales smooth, squarish with rounded posterior margins, imbricate, in 18 transverse rows; longitudinal ventral scale rows at midbody 6; anterior and posterior preanal plate scales paired. Scales on tail like those on body; dorsal and dorsolateral caudal scales hexagonal with keel, ventral and ventrolateral caudal scales square, smooth.
Limbs pentadactyl; digits clawed; dorsal brachial scales polygonal, subequal in size, subimbricate, smooth; ventral brachial scales much smaller than dorsal scales, round, juxtaposed, smooth; dorsal and ventral antebrachial scales polygonal, subequal in size, smooth. Scales on dorsal surface of manus polygonal, smooth, subimbricate; scales on palmar surface of manus small, rounded, subimbricate, domelike; thenar scales two, smooth laterally, raised into domes medially; finger length formula IV> III> II> V> I; scales on dorsal surfaces of fingers smooth, quadrangular, covering dorsal half of digit, overhanging supradigital lamellae 3/4 on I, 6 on II, 8 on III, 10 on IV, 5 on V; subdigital lamellae 6 on I, 9/10 on II, 13/14 on III, 14/15 on IV, 8/9 on V. Scales on anterodorsal surface of thigh large, polygonal, smooth, subimbricate; scales on posterior surface of thigh small, rounded, subimbricate; scales on ventral surface of thigh large, rounded, flat, smooth; femoral pores 9/11; preanal pores absent; scales on anterior surface of crus polygonal, smooth, imbricate, decreasing in size distally; scales on anterodorsal surface of crus polygonal, subimbricate, some scales with a slight keel; scales on ventral surface of crus large, polygonal, smooth, flat, imbricate. Scales on dorsal surface of pes polygonal, smooth, subimbricate; scales on palmar surface of pes small, rounded, subimbricate, domelike; toe length formula IV> III> V> II> I; scales on dorsal surface of digits single, quadrangular, smooth, of varying sizes, overhanging supradigital lamellae 3 on I, 5 on II, 8/9 on III, 10 on IV, 8 on V; subdigital lamellae single distally, double proximally, 5/6 on I, 10/11 on II, 15/16 on III, 18/20 on IV, 14 on V; limbs overlapping when adpressed against the body.
Coloration in preservative. Dorsal surface of head dark brown. Lateral surface of head like dorsal surface, a distinct cream stripe initiates on the ventralmost portion of the first supralabial scale and continues through all supralabials onto body, after the first supralabial the stripe bends dorsally to the tops of supralabials 3 and 4, covering approximately half of the final subocular, then the stripe bends ventrally through the middle of the final supraocular in a straight line to the bottom of the auricular opening, onto the body; a second, fainter cream stripe originates at loreal and continues posteriorly through superciliaries, dorsalmost postocular, and supratemporals, continuing onto body. Ventral surface of head yellowish cream with small brown stippling on the chin shields and lateralmost pregulars. Gular region like head but posteriormost gular scales with denser brown stippling per scale, forming clumps.
Dorsal surface of body same color as head, keels tipped in light cream; the faint dorsolateral cream stripes continue longitudinally onto tail, the narrowest distance between the stripes is six dorsal scales wide near the forelimb insertions, the widest distance between the stripes is seven scales wide, approximately two-thirds of SVL. Lateral surface of body same coloration as dorsum, lateral stripe from head continues over the forelimb insertion, briefly stopping at the hindlimb insertion and continuing at the same height posterior to the hindlimb insertion onto the tail, the stripe becomes less distinct after the forelimb insertion. Ventral surface of body slightly paler brown than the dorsal and lateral surfaces, with cream coloring on the posteriormost surface of each scale, medialmost ventrals are slightly paler in color than more lateral scales.
Dorsal surface of forelimbs with mottling the same brown color of the dorsal body, ground color orangish cream, cream large irregular blotches; dorsal surfaces of manus yellowish cream with small brown stippling; ventral surfaces of forelimbs cream with dense brown mottling on lateral surfaces of antebrachial and diffuse brown mottling on manus. Dorsal surfaces of hindlimbs orangish cream with diffuse brown mottling; dorsal surfaces of pes similar but with less mottling; ventral surfaces hindlimbs similar to that of forelimbs.
Dorsal tail coloration made of three distinct stripes, dorsolateral stripes orangish-cream, much more distinct and wider than on body, dorsolateral stripes begin to converge and dark brown coloration fades to stippling, which disappears altogether at approximately one-third of the tail length, rendering the entire dorsal surface of the tail orangish-cream. Lateral surfaces of tail defined by three stripes, the top and bottom stripes dark brown and the middle stripe yellowish-cream; at approximately one-third of the tail length the bottom stripe disappears. Ventral surface of tail yellowish cream. All surfaces of the regenerated portion of the tail a slightly darker brownish-cream.
Coloration in life. Dorsally a bronzy tan with sooty highlights on head, becoming paler on body at middorsal region; an intense creamy white stripe begins on eyelid and passes posteriorly to approximately midbody where it fades to nearly the ground color, this stripe is bordered dorsally by a narrow black stripe which begins just distal to eye and becomes faded and broken at midbody, the white stripe is bordered ventrally by a wide, shiny black stripe which begins at snout and extends posteriorly, passing just above extremities and narrowing at tail but continuing to tail tip, this wide stripe is, in turn, bordered ventrally by another intense pearly white stripe which begins just distal to rostral and passes through upper labials, just superior to insertion of forelimb, and gradually fading until its termination at insertion of hindlimb. This is bordered ventrally by another black stripe which begins at the first infralabial and passes squarely through the rictus, extending posteriorly through the forelimb and fading at the insertion of the hindlimb. Ventrally a greenish pearly white beneath the head and neck, black edging on the infralabials; ventral ground color gradually changes to a soft peach orange beneath the abdomen and it intensifies to a pinkish orange on hindlimbs, vent, and tail. Forelimb dark with pale spot on upper arm, elbow, and wrist; soft orange ventrally. Hindlimb brindled orange. Tail intense pinkish orange with black lateral stripe and another poorly defined broken black stripe from just distal to hindlimb, fading at mid-tail; a faint black dorsal line is evident in the sacral region. Iris deep brown, pupil ringed in gold. Tongue pink with dark gray forks.
Hemipenis. The organ was almost completely everted during preservation. It is broad and weakly symmetrically bilobed. The sulcus spermaticus runs a medial course until it bifurcates and runs onto the apical surfaces of the lobes. The organ has 14 oblique plicae (flounces), 12 of which occur on the lobes. The plicae bear comblike rows of spicules.
Variation. All paratypes are juveniles SVL 22.54–23.65 mm; complete original tails of juveniles 35.88–41.50 mm. The paratypes are very similar to the holotype with the following minor exceptions. In some specimens the frontal only makes contact with the first supraocular (versus the first two supraoculars in the holotype); in such cases the frontoparietal makes contact with all three supraoculars; in one specimen the frontal is irregularly octagonal. In UTA R-60252 the frenocular does not make contact with the first superciliary. In UTA R-60251 on the right side the loreal is much larger, making contact with the supralabial and preocular, separating the frenocular from the nasal and first superciliary; there are five superciliaries on the right side. The left side of this specimen resembles the holotype except the loreal makes contact with the preocular. In UTA R-60250 there is one pair of genials in contact with the first three infralabials. In UTA R-60250 and UTA R-60251 the posterior cloacal plate is made up of four scales.
Meristic variation in the paratypes includes: gular scale rows 5–7; transverse dorsal rows 28–31; longitudinal dorsal rows at 5th ventral row 8–11, at 10th ventral row 11–14, at 15th ventral row 10–14; lateral scale row 4–7 scales wide; transverse ventral rows 16–20; supradigital lamellae on Finger IV 9–11; subdigital lamellae on Finger IV 14–17; femoral pores 8–12; supradigital lamellae on Toe IV 10–12; subdigital lamellae on Toe IV 18–22.
Coloration in the juveniles in preservative is quite similar to the holotype. Overall they have paler pigmentation. The dorsal surface is orangish-brown instead of dark brown. The lateralmost stripe is much wider and covers almost the entire surface of all supraoculars, the stripe is thickest beneath the eye; the second stripe is more distinct than in the holotype, especially posterior to the eye. Ventral surface of the head is like that of holotype but with no stippling; stippling is present in the gular region in some specimens and on the ventral body in all specimens, but not as dense as in the holotype. The dorsal body stripes are much more distinct throughout length of body and tail; there are four to six scales between stripes.
There is color information for two of the paratypes in life (UTA R-60248 and 60250). In general the paratypes are more brightly colored than the holotype. However, in UTA R-60250 the dorsal ground color is dark gray on head and back, with black on sides. In both paratypes the dorsolateral stripe is pale yellow, becoming more orange distally. The lateral stripe is pale yellow. In UTA R-60248 the stripes are bordered in black and ventral and limb surfaces are similar to the holotype. In UTA R- 60250 the tongue is dark gray with white tips whereas in UTA R- 60248 the tongue is pink with yellow tips and a blue patch at the union of the two forks. In UTA R-60248 the umbilical scar is plainly visible.
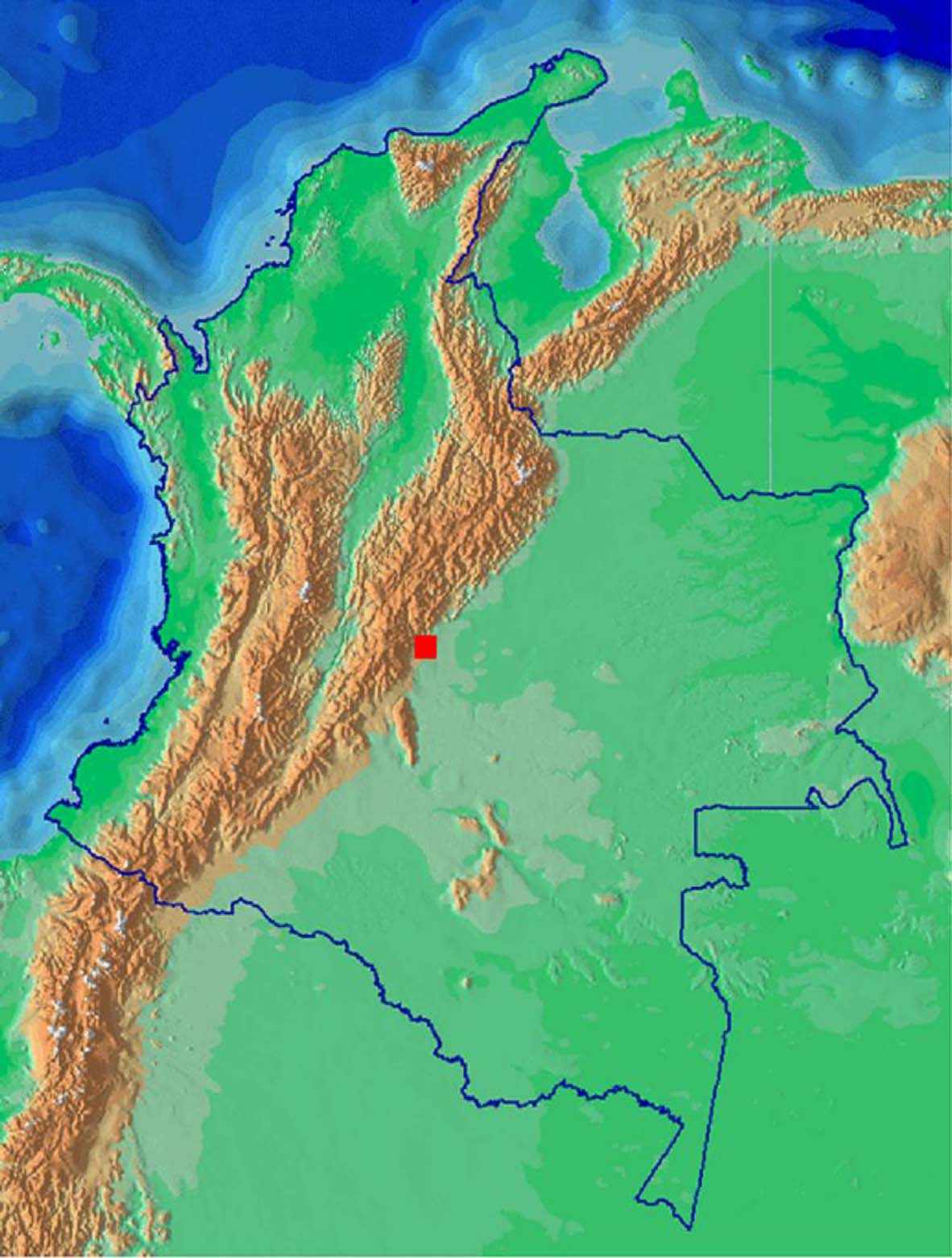
Distribution and natural history. Cercosaura hypnoides is known only from the type locality on the Vereda Portachuelo ( Fig. 5 View FIGURE 5 ). Vereda Portachuelo is a ridge of the Cordillera Oriental of the Andes Mountains at 1560–1800 m elevation, at Kms 13–15 along the road between Guayabetal and Manzanares, Municipality of Acacías, Department of Meta, Colombia. Type locality is 1640 m. The habitat is premontane pluvial forest ( Espinal, 1977), receiving over 4000 mm annual rainfall. The annual temperature range of the type locality is 18–24 ˚C. There is steep, rugged terrain with numerous rocky streams, and relatively tall undisturbed forest that has an abundance of mosses, lichens, aroids, orchids, bromeliads, and vines, creating dense vegetation. Plant associations include trees and shrubs of the genera Vismia (lanzo, carate), Albizia (pisquín), Cecropia (yarumo), Trema (zurrumbo), Heliocarpus (balso blanco), Solanum (lulo), Cedrella (cedro), Ochroma (balso), Miconia (tunos), Saurauia (dulumoco), Insertia, Inga (guano), and Pollalesta . The area contains many man-made clearings and small pastures. Owing to the high precipitation, runoff is fairly constant and soils are washed regularly. Nighttime temperatures drop rather sharply, causing fog; high humidity produces a significant chill factor.
Syntopic amphibians included Allobates sp., Atelopus minutulus Ruiz-Carranza, Hernández-Camacho & Ardila-Robayo , Centrolene (undescribed), Hyalinobatrachium orientale (Rivero) , Hyloscirtus denticulentus (Duellman) , Gastrotheca nicefori Gaige , Rhaebo glaberrimus (Günther) , Rheobates palmatus (Werner) , Rhinella sp., and various Pristimantis species. Reptiles included Anolis lamari Williams , Pseudogonatodes peruvianus Huey & Dixon , Ptychoglossus bicolor (Werner) , Bothrops atrox (Linneaus) , Chironius monticola Roze , Dipsas (undescribed), Liophis taeniurus (Tschudi) , Mastigodryas boddaertii (Sentzen) , Micrurus (undescribed), Ninia atrata (Hallowell) , Oxyrhopus leucomelas (Werner) , and Synophis lasallei (Maria) . The type locality, Portachuelo, is the type locality of Atelopus minutulus and Anolis lamari , which demonstrates the importance of this site for novel herpetofaunal species.
The holotype was collected by day beneath a rotting log on a steep treeless slope with a southeastern exposure. There was heavy moss growth all around.
All the paratypes were collected as eggs. A total of 30 eggs were found during a two day period (08–09 July) along an open slope with western exposure at Vereda de Portachuelo. On 0 8 July a total of 14 eggs (3 empty) were located in heavy moss usually at the edge of rotted logs and frequently in nests of stinging ants (genus Odontomachus ). On two occasions the eggs were plainly visible on logs or exposed moss. All were in distinct stages of development, some recently deposited and others ready to hatch. On 0 9 July in 5 sites on the same western exposed slope a total of 16 eggs were found. These were either single, double (in one case two freshly laid small eggs), or in one case a total of 11 eggs were found in a communal nest in a pocket of earth beneath a log. Two of the latter had hatched and the remaining nine were of various sizes. All unhatched eggs were maintained in a plastic bag filled with moss from the collection site. Two of the paratypes emerged the same day that the eggs were collected. The other four paratypes (UTA R-60251–60254) hatched 10–18 days after collection. Dates of hatching ranged from 8–27 July. The egg sizes that hatched measured from 13.0 x 8.0 mm to 15.0 x 10.0 mm and were dirty tan in color. Dimensions of the others that had not hatched (taken 27 July 1980) as follows: 11.0 x 14.0 mm; 9.5 x 14.0 mm; 11.0 x 9.0 mm; 12.0 x 9.0 mm; 11.0 x 7.5 mm; 11.0 x 8.0 mm; 11.0 x 8.0 mm; 9.0 x 12.0 mm; 9.0 x 11.4 mm. In addition, one egg, which measured 11.0 x 15.0 mm, was opened on 27 July 1980 and was found to contain a fully developed, motile embryo.
Etymology. The specific epithet is taken from the New Latin adjective hypnum, in turn derived from Greek hupnon (a type of lichen, + -oid) meaning “of the moss in reference to the luxuriant moss growth that characterized the lizard’s habitat and egg deposition sites.




| UTA |
University of Texas at Arlington |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cercosaura hypnoides
| Doan, Tiffany M. & Lamar, William W. 2012 |
Pantodactylus (
| Ruibal 1952 |