
Parainvenusta englarorum Curini-Galletti
|
publication ID |
https://doi.org/ 10.5281/zenodo.196653 |
|
DOI |
https://doi.org/10.5281/zenodo.6209507 |
|
persistent identifier |
https://treatment.plazi.org/id/F87F0737-C76E-FFA2-FF3C-C01EB2AD03F8 |
|
treatment provided by |
Plazi |
|
scientific name |
Parainvenusta englarorum Curini-Galletti |
| status |
sp. nov. |
Parainvenusta englarorum Curini-Galletti sp. nov.
(Fig. 2)
Holotype: One specimen sagittally-sectioned ( SAM AHC 29915)
Type locality: Port Lincoln, South Australia. Sleaford Bay, Wreck beach (34°51’36.71”S; 135°43’41.19”E), fine sand, Jan. 2001
Additional material: one specimen sagittally sectioned (paratype: SMNH type-7950); four specimens sagittally sectioned (CZM 181-183); two specimens frontally-sectioned (CZM 184, 185); four whole mounts (CZM 186-189); two karyological slides (CZM 190, 191.). Sleaford Bay, Lone Pine beach (34°52’01.45”S; 135°42’44.98”E), medium-fine sand, Jan. 2001: three specimens sagittally-sectioned (CZM 192-194).
Etymology: the species is dedicated to Giuseppe and Saturna Englaro, for their long and strenuous struggle for the respect and actuation of the wills of their daughter Eluana (1970–2009), in irreversible vegetative state for 17 years, giving an exemplary lesson of laicity, love, and faith in legality.
Description. Animals slender; fixed mounts up to 6 mm long. Anterior end with few sensory bristles. Body entirely ciliated; cilia about 3 µm long. Epithelium with insunk nuclei. With numerous adhesive papillae in the tail; a few are scattered along the length of the body.
Different types of glands are present at the periphery of the body:
i) small, intensely stained eosinophylic glands (g 1 in Fig. 2 C), about 5 µm long, with the main body below the basal lamina, and whose necks pierce the overlaying epithelium;
ii) numerous ovoid rhabdoids (Figs 2A–C: r), 15–20 µm long, particularly dense in the caudal region;
iii) long paracnida , containing an eversible tube, distinctly granular at its proximal end. Two types of paracnida , differing in position and morphology, can be recognized. A few (6–10), very large (up to 60 µm long) and nearly straight, are placed radially around the genital area, and in most specimens form a rosette around the genital opening (Figs 2A, B: p1). Others (20 – 30 µm long), more numerous and more irregularly coiled, are distributed mostly in the caudal region (Figs 2A, B: p2), where they are intermingled with rhabdoid glands.
The brain is encapsulated, and abuts anteriorly the statocyst, which is about 20 µm across. The two neural cords originating ventrally from the brain are easily appreciable in sections of the anterior part of the body.
Subepithelial musculature thin, formed by an external, circular and an inner longitudinal layer. Numerous dorsoventral fibres are present, regularly arranged along the length of body. The gut extends from the cephalic area to the tail region.
Pharynx in the posterior 3rd of body. It is tubular, up to 500 µm long, and directed horizontally. It is provided with a thin layer of external longitudinal musculature and an inner, thicker, circular component. The pharynx is ciliated (cilia 2.5–3 µm long) apart from the distal tip, where long pharyngeal glands, which extend considerably anterior to the pharynx, discharge. With a short (20–30 µm long) oesophagous, lined with unciliated, nucleated epithelium.
Pharyngeal pocket lined with unciliated, nucleated epithelium; mouth ciliated.
Male genital system. With few testes (20–30), arranged in two irregular rows in front of the ovaries. The testes are placed laterally, abutting the vitellaria, and often intermingled with them.
Copulatory organ consisting of a globular bulb, connected to two seminal vesicles. The seminal vesicles are lined by a thin, flat epithelium, and extend laterally and posteriorly from the bulb, into which they enter dorsally. The bulb is 45– 50 µm across, lined externally by a flat, intranucleated epithelium. Below the epithelium, a strong muscular lining, 10–12 µm thick, is present. It consists of relatively few and very large muscle cells, arranged into an external longitudinal and an inner, thicker, circular layer. The bulb is posteriorly oriented, and is connected to the body by a relatively small, ventral area, crossed by muscular fibres which are connected to the body musculature. The bulb contains a cirrus, which originates from the fusion inside the bulb of the distalmost portions of the two seminal vesicles. No prostatic vesicle was observed, nor cell bodies that could be identified as prostatic glands. The cirrus is unarmed, long and very convoluted, so that its length is impossible to measure with any degree of confidence in sections. It is lined internally, at least in parts, by a basal lamina, without overlying epithelium, and it is surrounded by a thin coating of muscular cells, more noticeable distally. The cirrus opens ventrally into the atrium. The rest of the bulb appears filled with somewhat loose parenchymatic cells, with ‘empty’ spaces among cells.
Female genital system. Vitellaria at both sides of the body, in front of the pharynx. Two ovaries in front of the pharynx.
Oviducts ciliated; they run posteriorly, and open independently at the two sides of a small bursa. This is nearly spherical, with a lumen about 10 µm across, and, in most specimens, contains sperm. The bursa is lined by an unciliated cuboidal epithelium, and opens anteriorly into the dorsal wall of the atrium through a short, ciliated bursal canal, 10–15 µm long, which receives the female glands. Posteriorly, the bursa is connected to a long (50–60 µm in sections), Sshaped duct, surrounded by very strong circular muscles. The muscle ring closest to the bursa is the strongest, and may act as a sphincter. This duct is lined by an unciliated, nucleated epithelium, and opens into the posterior-most portion of the gut, thus forming a genito-intestinal connection. In most specimens, the posterior gut appears filled with sperm, suggesting the existence of a distinct resorbing bursa in this part of the intestine.
The common atrium is narrow, lined with an unciliated epithelium. The genital pore is surrounded by a sphincter.
Karyotype. The only plate suitable for study had n=5, with Chrom I isobrachial, and the rest of chromosome pairs heterobrachial. The smallest chromosome pair was about half the size of Chrom I.
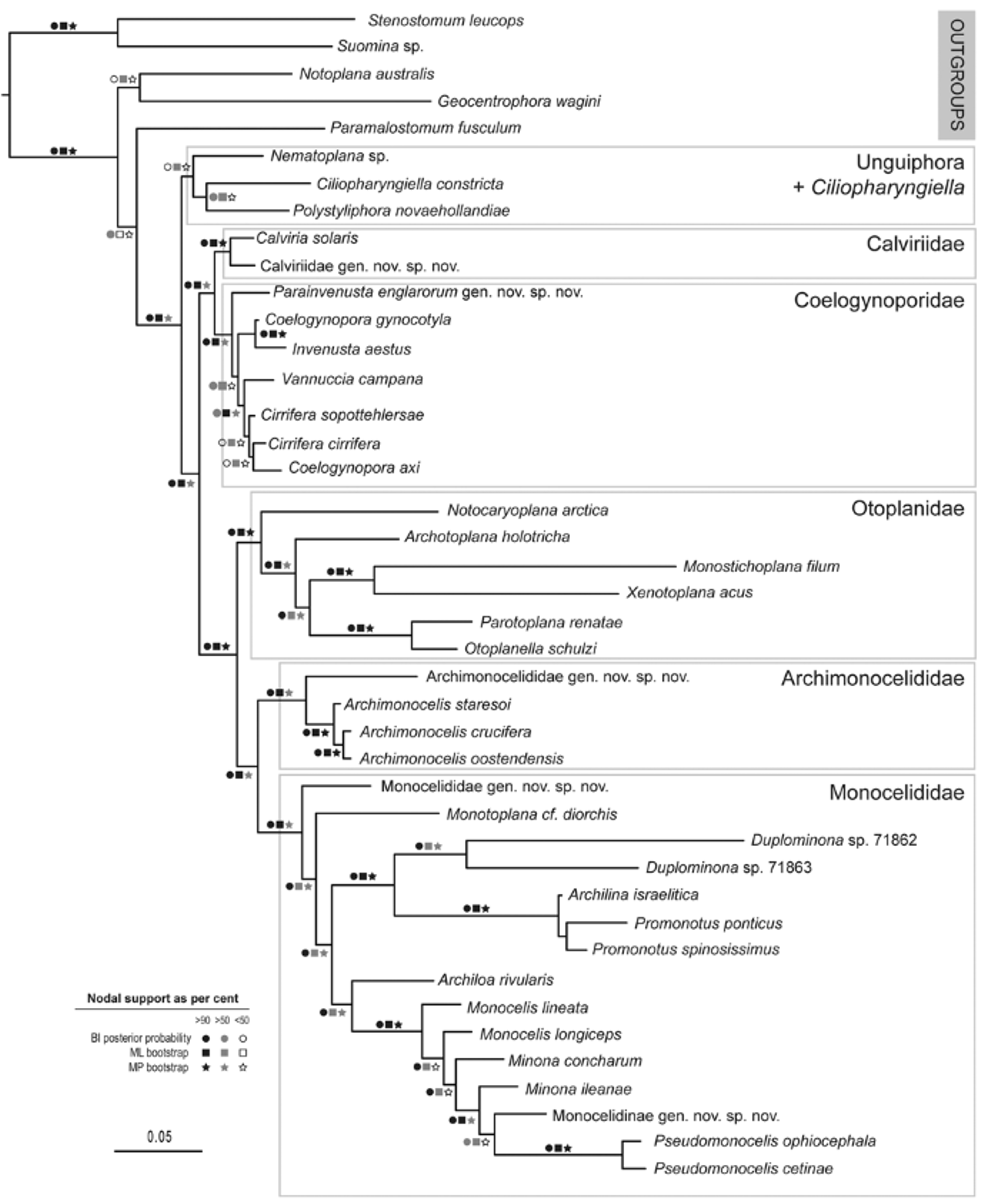
Discussion. The combination of characters found in the new species is unique, and, although clearly attributable to the Coelogynoporidae for a combination of characters (see discussion above), could not be fit into any extant genus. Indeed, albeit biased as this may be by incomplete taxonomic sample of coeloegynoporid genera, the species occupies a basal, isolated position in the molecular tree ( Figure 1 View FIGURE 1 ).
As recalled by the etymology, Parainvenusta englarorum gen. nov. sp. nov. superficially resembles species of Invenusta , which possess similar strikingly long paracnida , and have an unarmed copulatory organ. Two species of Invenusta are known: I. aestus Sopott-Ehlers, 1976 (type species) from the eastern Atlantic (Canary Islands and French Atlantic coast) and I. paracnida (Karling, 1966) from western U.S.A. (Bodega Bay, California, and S. Juan islands, Washington). Both species have long paracnida , spread all over the body, and a copulatory organ provided with a muscular penis papilla, with the two seminal vesicles external to the bulb. Both lack a bursa, and the oviducts enter a ciliated diverticulum of the atrium. However, the two species appear to differ to an extent that their congenerity may be questioned. Among the main differences, the paracnida of I. aestus have the eversible part provided with barbs, while they are granular basally (and thus similar to the new species) in I. paracnida . The pharynx is short and oriented vertically, and the copulatory organ is anteriorly oriented, in I. aestus , while the pharynx is tubular and horizontal and the copulatory organ vertical in I. paracnida . Furthermore, in I. paracnida a long, convoluted genito-intestinal duct is present, while this is absent in I. aestus (Karling, 1966, Sopott-Ehlers, 1976). The structure of the female genital system in I. paracnida – consisting of a diverticulum of the atrium which receives the oviducts, and a long, winding genitointestinal connection - is indeed comparable to that of the new species. However, in P. englarorum , the ciliated duct, here interpreted as a bursal canal, leads to a bursa which receives the outlets of the oviducts, and is histologically different from the rest of the female system.
Furthermore, the copulatory organ is of the simplex type in Invenusta , i.e. it is formed by a single muscular wall which surrounds the last portion of the ejaculatory duct (Karling, 1956; Litvaitis et al., 1996). In contrast, in P. englarorum the cirrus bulb contains a (presumably) eversible cirrus, surrounded by a fluid matrix. The presence of two muscular walls, an outer and inner one, this latter corresponding to the last portion of the ejaculatory duct, fits the definition of duplex-type copulatory bulb (Karling, 1956).
Finally, the position of P. englarorum and I. aestus in the molecular tree ( Fig. 1 View FIGURE 1 ) does not support a close phylogenetic relationship among the two taxa.
Within the Coelogynoporidae , a duplex– type copulatory organ is present in the genera Cirrifera Sopott, 1972 , Vannuccia Marcus, 1948 , and Stilivannuccia Faubel & Rohde, 1998 . In these taxa, however, the cirrus is provided with numerous spines; and accessory stylets may be present. In the genus Cirrifera , the seminal vesicle is entirely extracapsular, and a prostatic vesicle is present. Furthermore, species of Cirrifera have a vertically oriented pharynx, and lack a bursa and true paracnida (Jouk et al., 2007) . In Vannuccia and Stilivannuccia , the seminal vesicle is anterior to the copulatory bulb, an intra-bulb prostatic vesicle is generally present, as well as a long female duct leading to the bursa.
A cirrus is also present in Macroatrium setosum Riser, 1981 from N.W. Atlantic (New England); although its morphology is not known in details, it is spiny, broad, and cup-shaped, and thus very different from that of the new species. M. setosum lacks paracnida , and is provided with a comparatively very large atrium and a seminal bursa roughly of the same size as the atrium (Riser, 1981).
Similarly to Parainvenusta englarorum , Pseudovannuccia hirutai (Tajika, 1981) is characterized by an unarmed copulatory organ. However, in this species the copulatory organ is of the simplex type (Faubel & Rohde, 1998), with a highly developed, strongly muscular penis papilla. Furthermore, the seminal vesicle is entirely within the copulatory bulb and anterior to the gonopore (Tajika, 1981).
Given the morphological distinctiveness of the new species, its inclusion in any of the genera above would render that genus exceedingly heterogenous; and a new genus is thus proposed for it.
FIGURE 2. Parainvenusta englarorum nov. sp. nov. gen. A: general appearance; B: Postpharyngeal genital organs, based on observations on living squeezed specimens and frontal sections; C: sagittal reconstruction of the postpharyngeal genital organs; D,E: genital area from frontal (D) and sagittal (E) sections. Scale bar: 25 µm. Abbreviations: a: atrium; b: bursa; bc: bursal canal; br: brain; ci: cirrus; co: copulatory organ; fg: female glands; g1: gland; gic: genito-intestinal connection; gl: gut lumen; gp: genital pore; mc: muscular lining of the cirrus bulb; mr: muscle ring; od: oviduct; ov: ovary; p1: paracnida (genital area); p2: paracnida (caudal area); ph: pharynx; r: rhabdoid gland; st: statocyst; sv: seminal vesicle; t: testis; vi: vitellaria.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.