
Austroconops Wirth and Lee
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2004)449<0001:AWALAL>2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5060649 |
|
persistent identifier |
https://treatment.plazi.org/id/F966E84E-FFAC-216E-FF15-FD4EFB3E06FA |
|
treatment provided by |
Felipe |
|
scientific name |
Austroconops Wirth and Lee |
| status |
|
Austroconops Wirth and Lee View in CoL View at ENA
Austroconops Wirth and Lee, 1958: 337 View in CoL . Type species: Austroconops mcmillani Wirth and Lee View in CoL , by original designation.
DIAGNOSIS: Male. The only extant or fossil Ceratopogonidae with flagellomere 13 ( fig. 1A, B View Fig ) with a subbasal constriction, with two welldeveloped radial cells, and rm parallel to R 1 ( fig. 1K View Fig ). Female. The only extant or fossil Ceratopogonidae with two welldeveloped, clearly open, radial cells and rm parallel to R 1 ( fig. 1L View Fig ). Egg. Only Ceratopogonidae with egg ( fig. 2A View Fig ) markedly elongate and remaining pale throughout larval development. Larva (all instars). The only Ceratopogonidae with markedly elongate antenna bearing an elongate blade and a prognathous head ( figs. 2H View Fig , 3C View Fig , 12A, B View Fig ). Pupa. The only Ceratopogonidae with the hindleg sheath not exposed from under lateral margin of wing sheath and with abdominal segments 2–8 with bifurcate setae ( fig. 4A View Fig ).
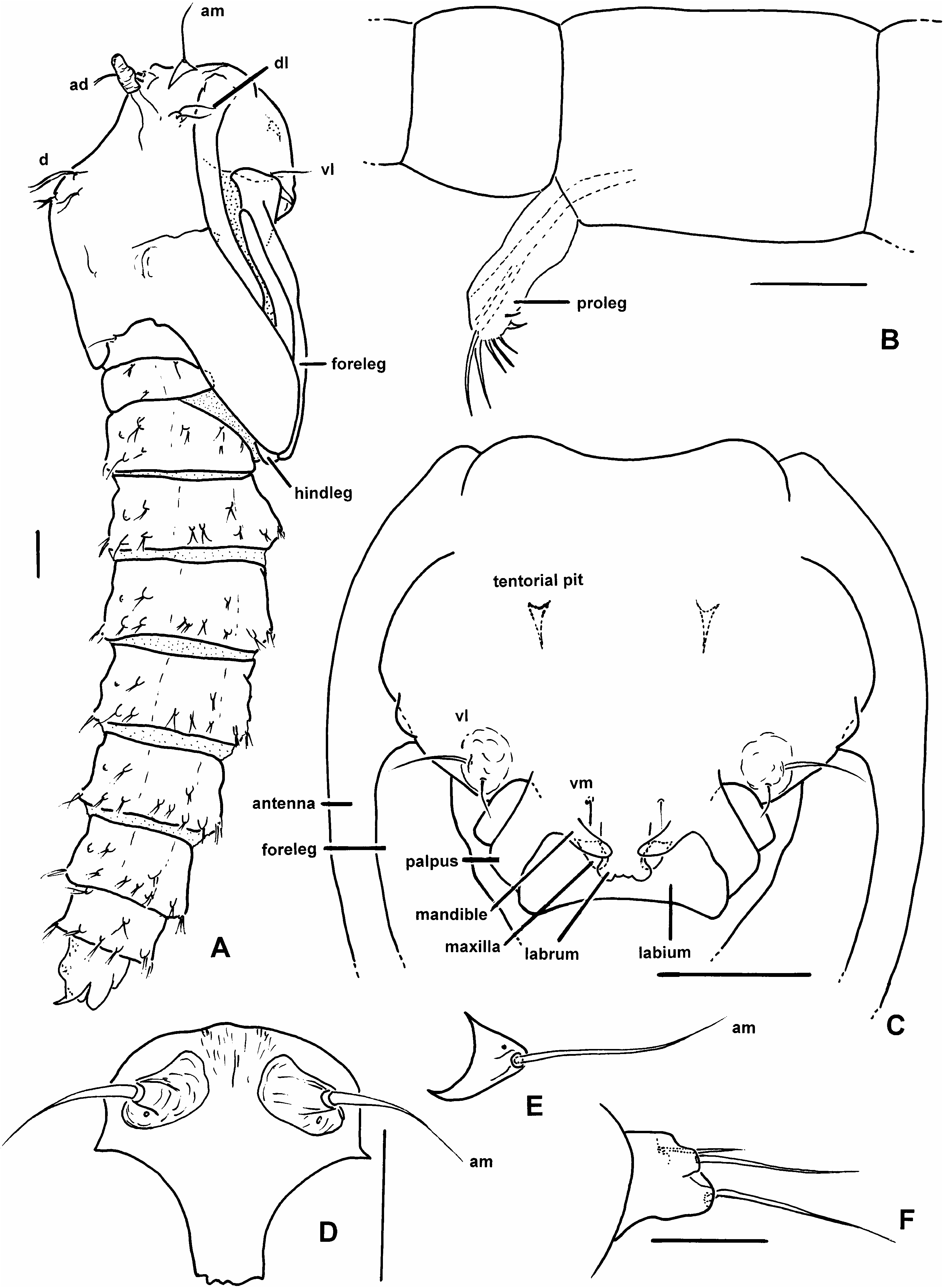
DESCRIPTION: Live adults of both sexes. With subcutaneous bluegreen tissue (likely fat body) externally evident in all areas (head, thorax, abdomen) with membrane or thin cuticle (including tip of halter). Wings completely overlapping at rest. Male with permanently erect antennal setae. Male adult. Head: Ommatidia narrowly separated dorsomedially, with single vertex seta. Antenna ( fig. 1A, B View Fig ) with welldeveloped plume of permanently erect setae, 13 separate flagellomeres, flagellomeres 12–13 more elongate than preceding flagellomeres, flagello mere 12 with or without subbasal constriction, flagellomere 13 with subbasal constriction, flagellomere 1 with two patches of short sensilla trichodea, without sensilla coeloconica. Mouthparts moderately short to moderately long. Mandible elongate (ending near apex of labrum), with several slender terminal spicules. Lacinia elongate, simple. Palpus ( fig. 1C, D View Fig ) with 4–5 segments, segment 3 ovoid to elongate, slender, with capitate sensilla scattered on mesal surface or perhaps in pit, at least extant species with membranous area between segment 3 and 4 + 5. Thorax: With three anterior pronotal apodemes. Scutum with scattered elongate setae. Scutellum rounded or angular in dorsal view. Anapleural suture elongate. Wing ( fig. 1K View Fig ): Without macrotrichia, fine microtrichia present on all membrane. Alula with fringe of macrotrichia. Costa extending to or beyond apex of R 3. Both radial cells present. rm parallel to R 1. M bifurcating distal to rm. Legs: Femora, tibiae slender. Legs lacking armature, except in some species with stout setae on some or all of first tarsomeres, some with pair of thick setae on apex of tarsomeres 1–4. Tarsal ratio (Ta 1 /Ta 2) of foreleg/hindleg = 1.4–1.9. Fore and midleg trochanter without pair of thick setae. Midleg tibia with or without apical spur. Hindtibia apex with two rows of spines. Hindleg first tarsomere with or without thick basal spine, with scattered setae ( fig. 1M, N View Fig ). Claws on fore, mid, hindleg equal in size, both equal on each leg, each claw simple or toothed, apically bifid (possibly simple in Lebanese amber fossils). Slender empodium. Genitalia: Apicolateral process absent to well developed. Gonocoxite short to moderately elongate. Gonostylus variable, apical spine present or absent. Parameres fused medially (known only in extant species). Aedeagus short, setose lobe with ventral plate (known only in extant species).
Female adult. Head: Ommatidia narrowly separated dorsomedially, with single vertex seta. Antenna with 13 separate flagellomeres, flagellomeres gradually increasing in length from flagellomeres 2–13 or with flagellomeres 9–13 more elongate than preceding flagellomeres, flagellomere 1 with two groups of short sensilla trichodea, without sensilla coeloconica. Mouthparts moderately elongate, further details not visible in fossils. Mandible ( fig. 1G, H View Fig ) and laciniae with fine teeth in extant species. Palpus ( fig. 1E, F View Fig , 23D View Fig ) with 4–5 segments, segment 3 ovoid to elongate, with capitate sensilla scattered on mesal surface, at least extant species with membranous area between segments 3 and 4 + 5. Thorax: With three anterior pronotal apodemes (known only in extant species). Scutum with scattered elongate setae. Scutellum angular in dorsal view. Anapleural suture elongate. Wing ( fig. 1L View Fig ): Without macrotrichia, fine microtrichia present on all membrane. Alula with or without fringe of macrotrichia. Costa extending to or beyond apex of R 3. Both radial cells present. M bifurcating distal to rm. rm parallel to R 1. Legs: Femora, tibiae slender. Legs lacking armature except in some with pair of stout setae on apex of tarsomeres 1–4 of all legs. TR of foreleg/hindleg = 1.7–2.2. Fore and midleg trochanter without pair of thick setae. Midleg tibia with or without apical spur. Hindtibia apex with two rows of spines. Hindleg first tarsomere with or without thick basal spine, with scattered slender or stout setae. Claws on fore, mid, hindleg equal in size, both equal on each leg, each claw simple or toothed ( fig. 1I, J View Fig ). Empodium slender. Genitalia: Two large, one markedly smaller spermathecae. Sternite 9 continuous medially. Segment 10 with pair of setae. Cerci short to moderately elongate.
Egg ( fig. 2A View Fig ): Very light yellowish brown, appearing nearly white during development of larva. Elongate and slender. With 9–10 longitudinal rows of short, minute tubercles, each tubercle slender at base, expanded distally ( fig. 2B View Fig ). Anterior end more rounded than posterior end. Eggshell opening a single dorsal slit along most of length.
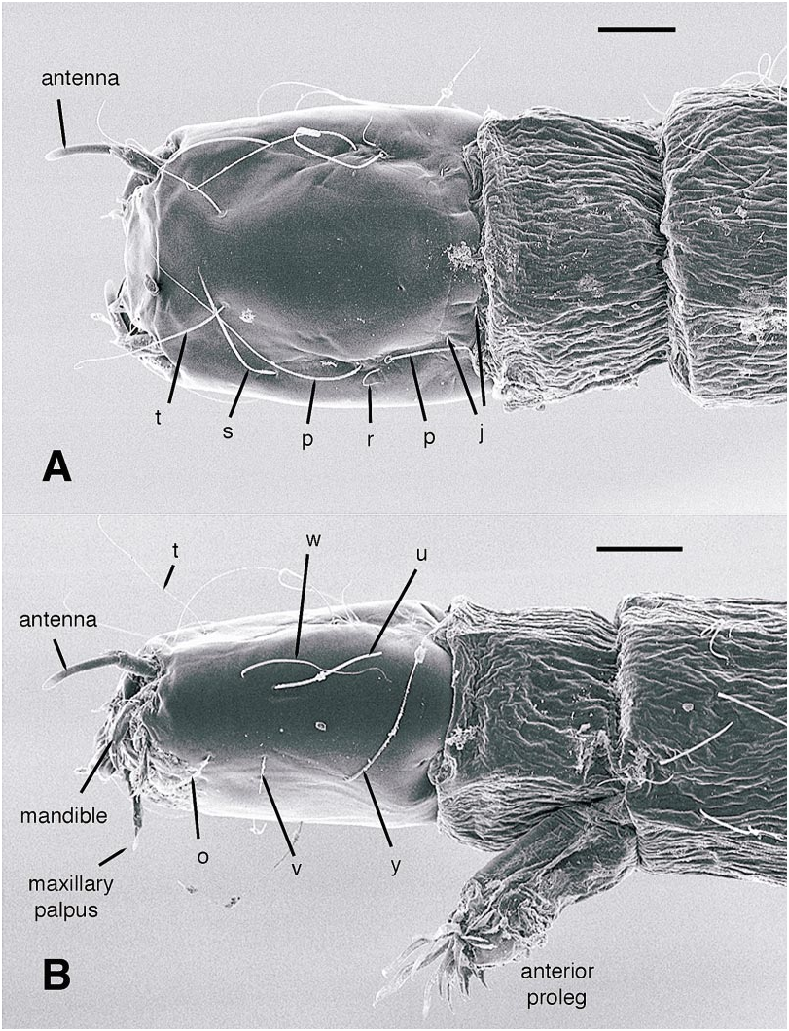
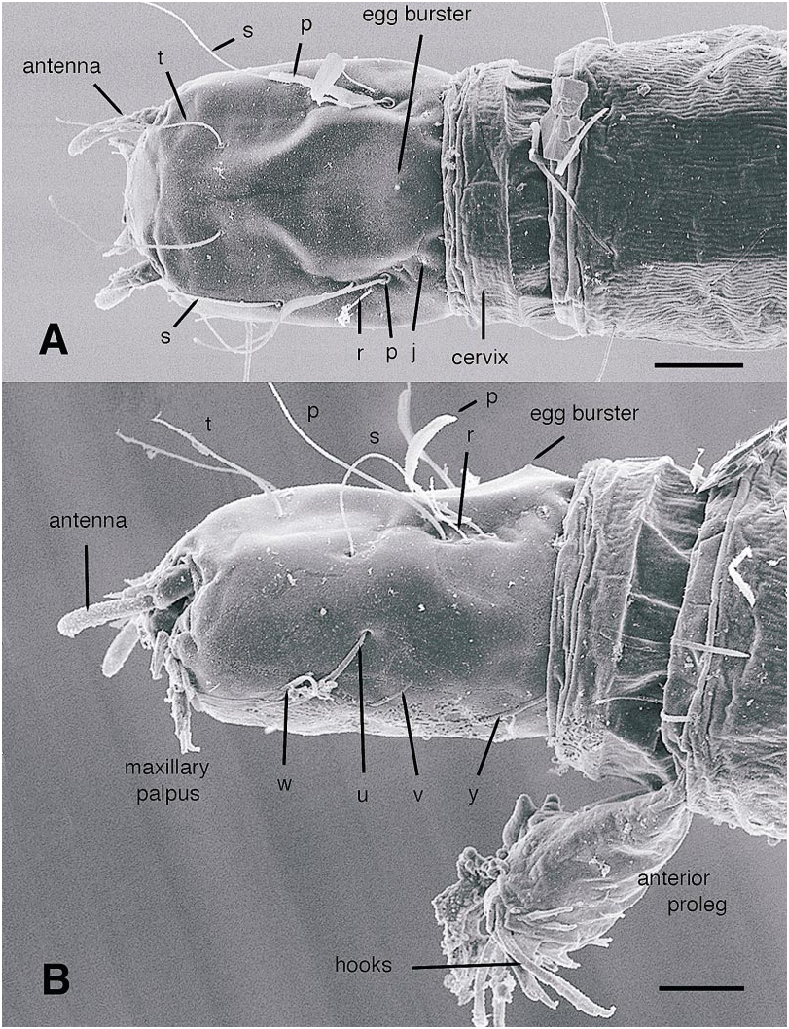
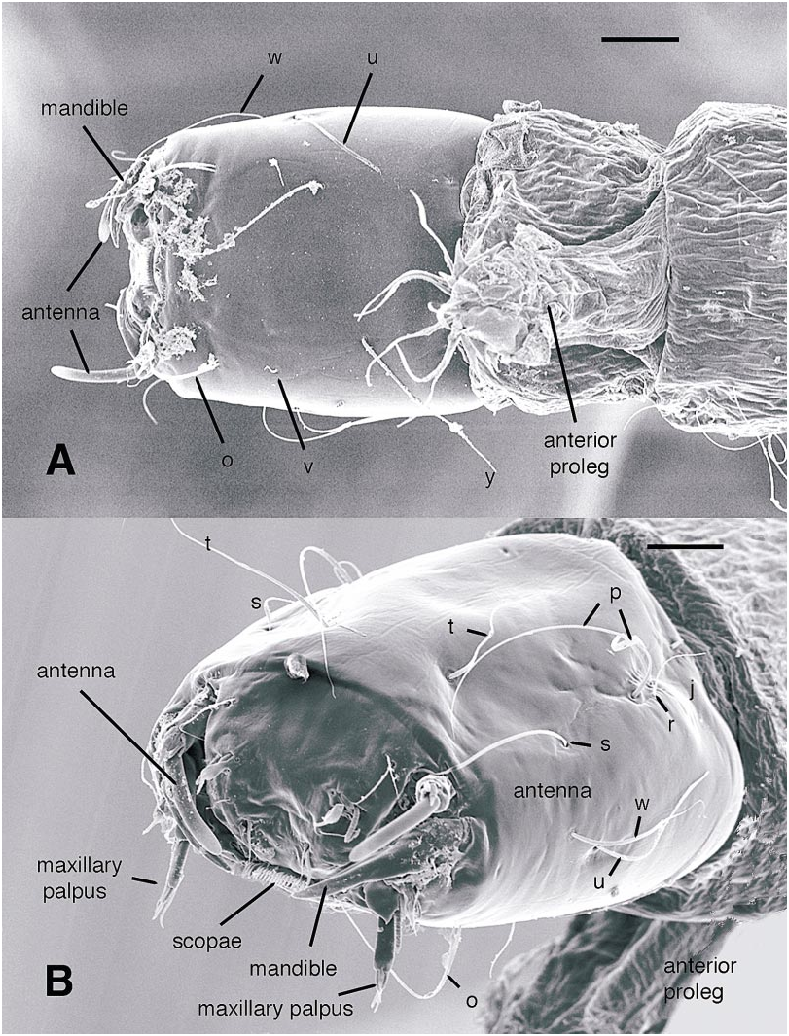
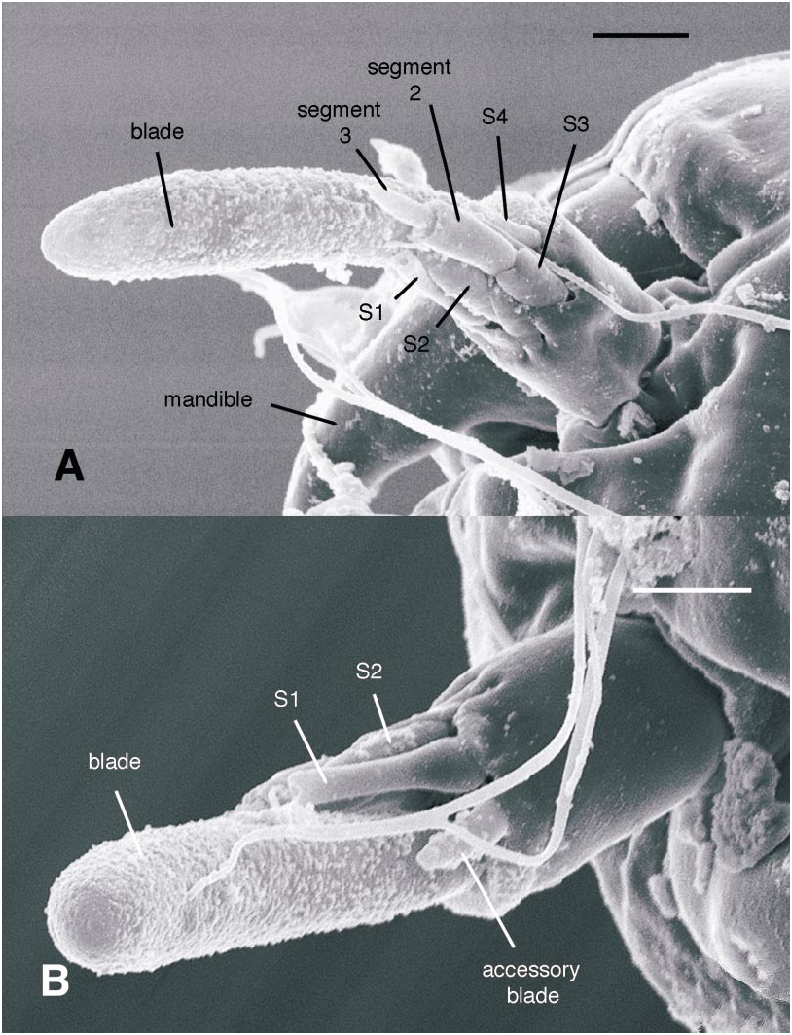
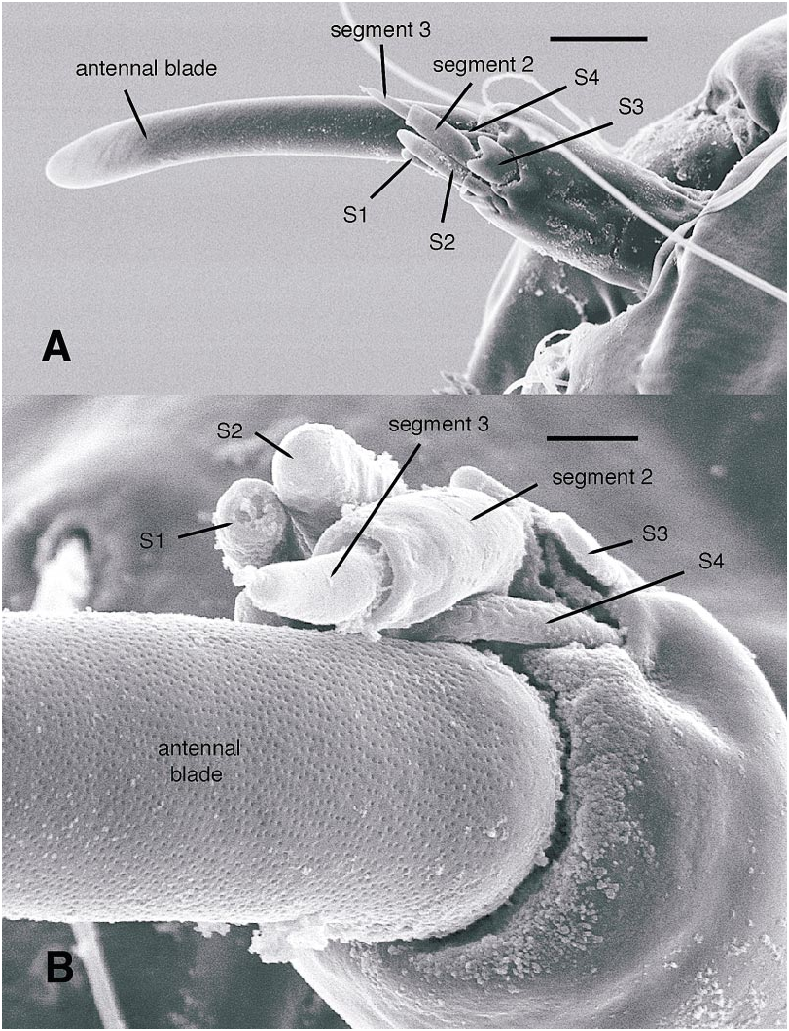
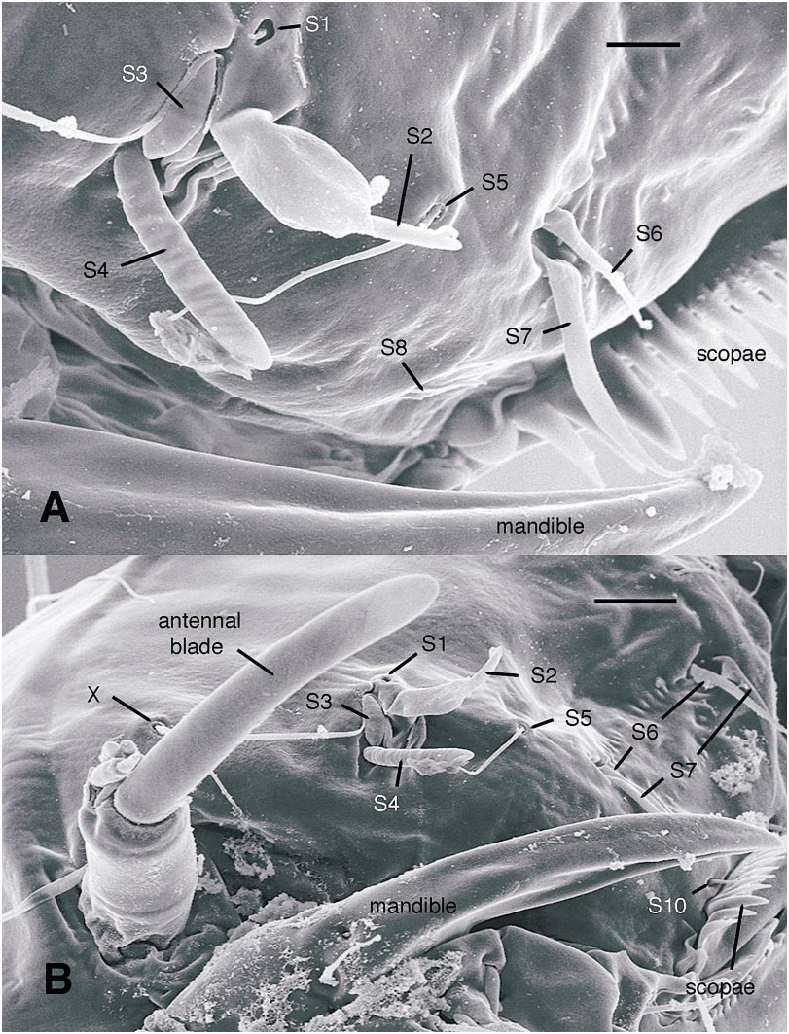
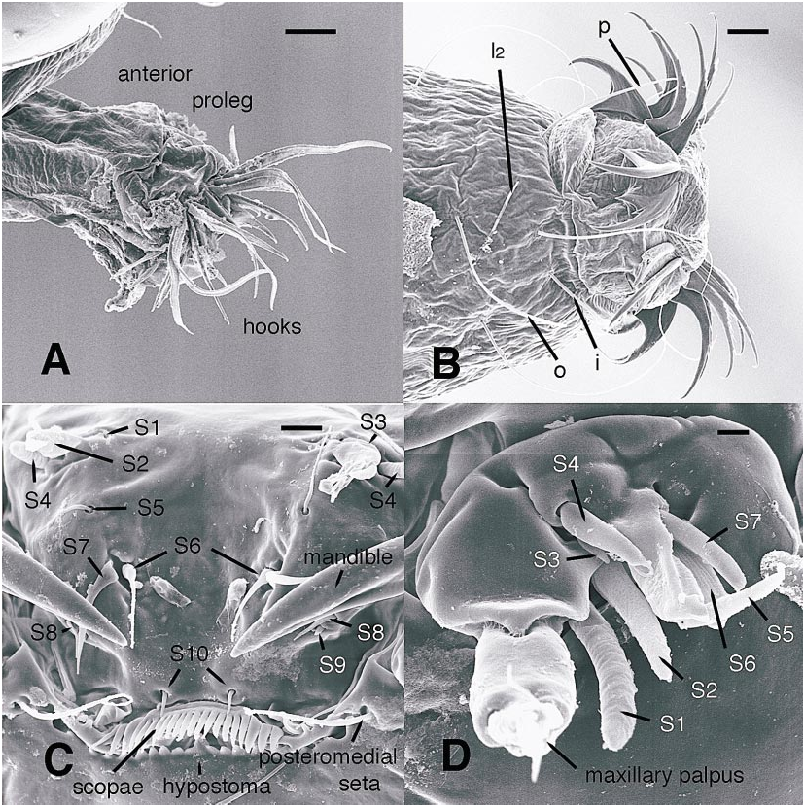
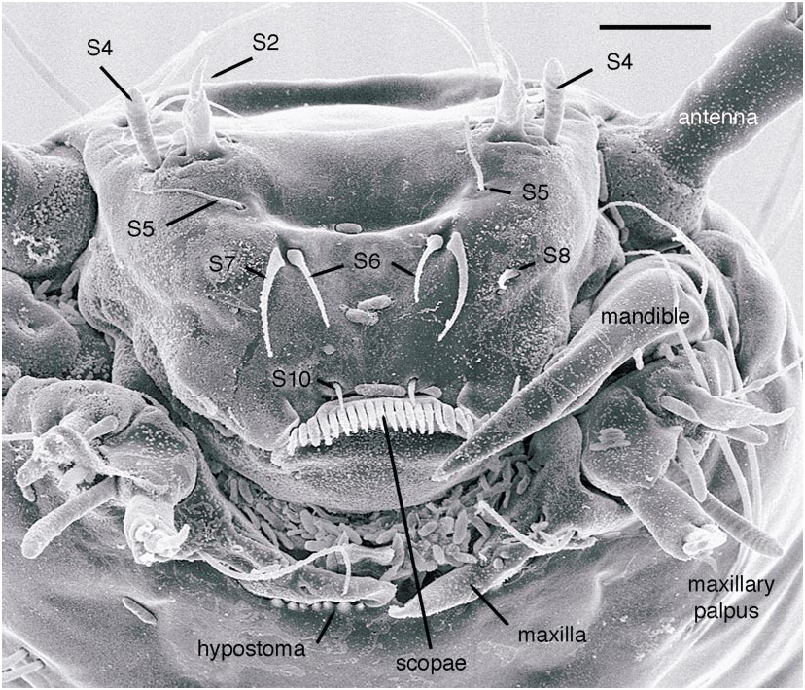
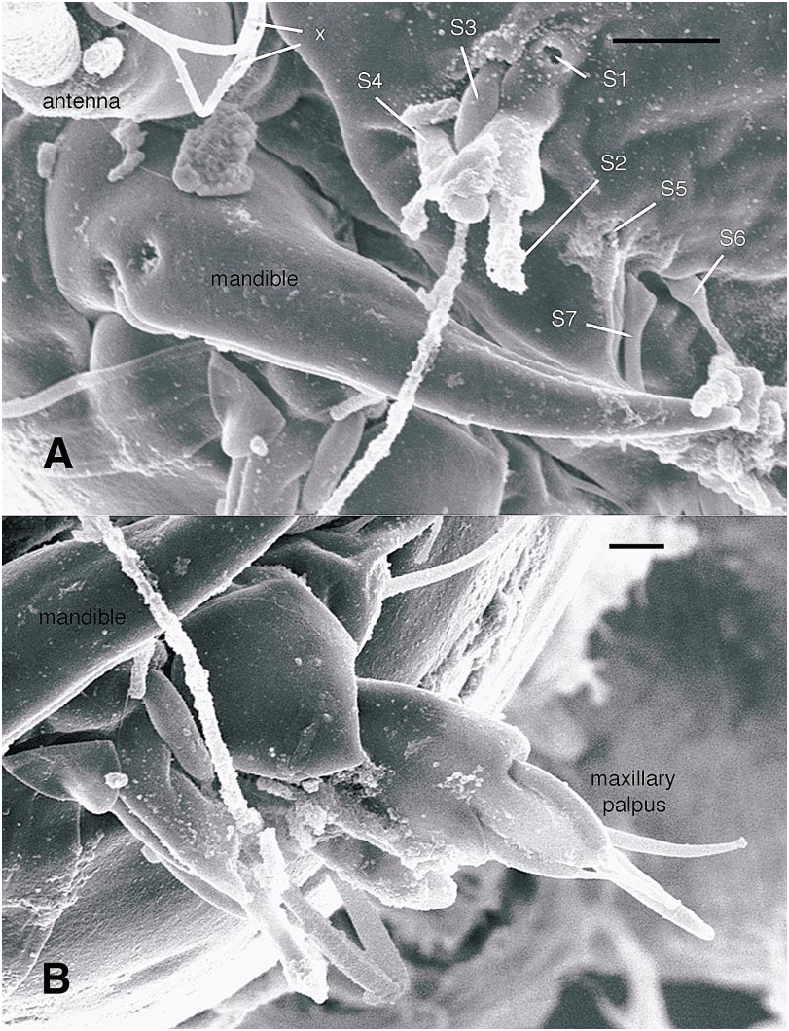
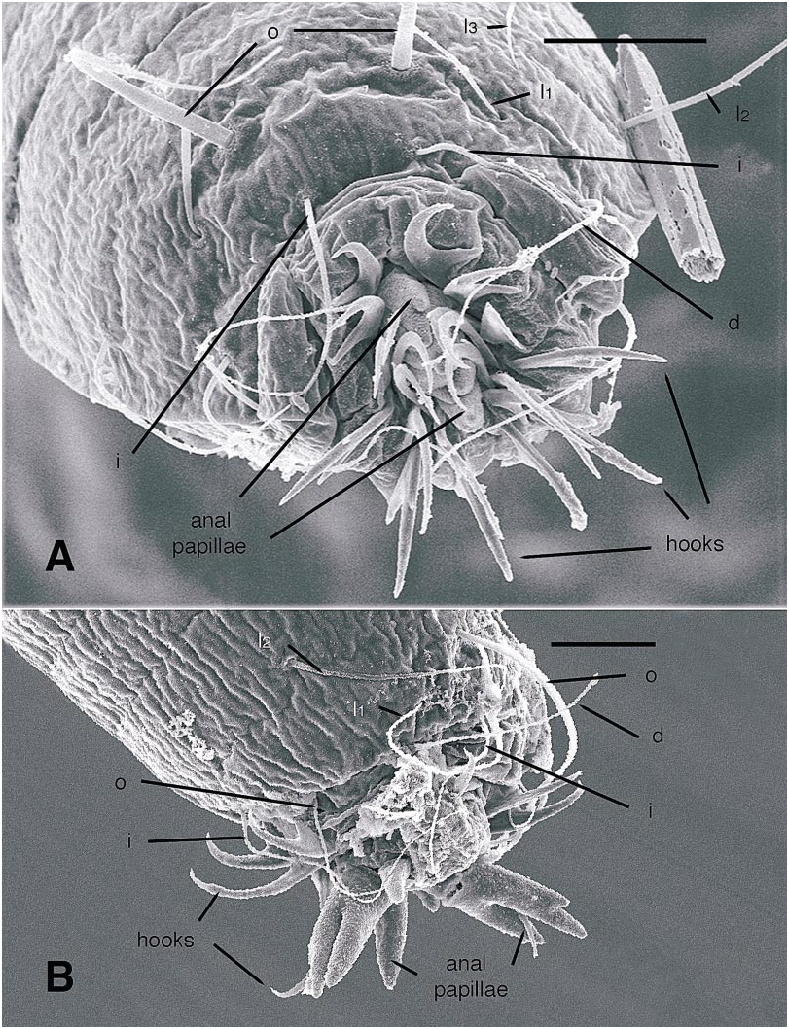
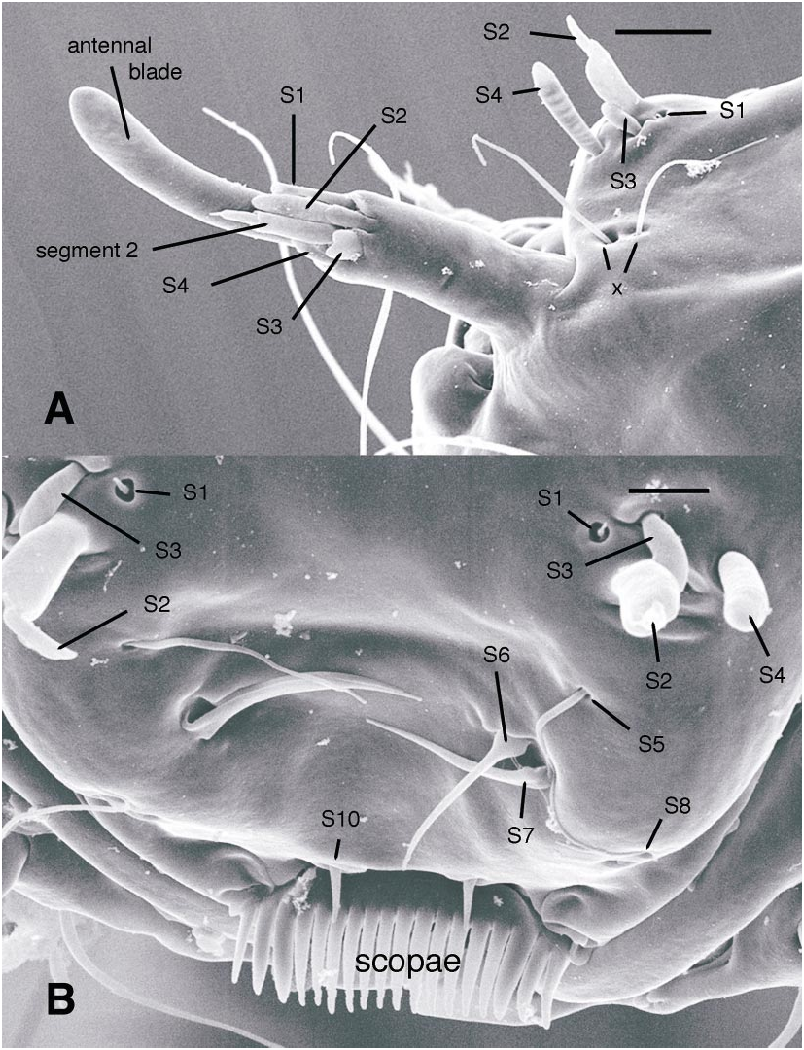
All larval instars: Head capsule ( figs. 2H, K View Fig , 3A, C, G View Fig , 7A, B View Fig , 8A, B View Fig , 12A, B View Fig , 13A, B View Fig ): Nearly square in dorsoventrally viewed outline; light brown, with only pigmentation present: dark epipharyngeal bar, premandible, apex of mandible; eye in later instars (some seconds, all thirds, fourths) a single, roughly circular to oval spot. Ecdysial suture extended anteriorly slightly beyond level of sensillum s, possibly longer but difficult to discern; area of thinner cuticle broad in area of sensilla j (anterior one), p, r. Ventral suture not present. Setae all simple (not bifurcate), arrangement as in figures 2H, K View Fig , 3C, G View Fig , 7A, B View Fig , 8A View Fig , 12A, B View Fig , 13A, B View Fig ; following sensilla present: x (both setae present), t (or possibly an anteriorly placed q), s, p (two long setae, posterior one expanded distally), r (a short peg), j (anterior one a short peg on head capsule; other a more elongate seta on anterior margin of cervix), o (both an equally long seta), w, u, m, v, and y. Antenna ( figs. 9A, B View Fig , 14A, B View Fig ) elongate, with three segments; with well developed, short, basal segment, segments 2–3 short, slender, segment 3 tapering to slender, elongate apex; first segment otherwise bearing greatly elongate, multiporous blade and five other sensilla: accessory blade, sensilla 1–4; sensillum 1 long, slender, with apical pit with small peg; sensillum 2 moderately elongate; sensillum 3 short, bilobed; sensillum 4 moderately elongate, with pores. Labrum ( figs. 3A View Fig , 15A, B View Fig , 17C View Fig , 19 View Fig ) wide, with well developed torma, with well developed, dark, articulating, apically bent premandible; torma abutting lateral margin of stout, black, transverse epipharyngeal bar; epipharyngeal bar forming large, ventrally directed, internal, median lobe, posterodorsally directed bilobed apodeme and lateral apodeme abutting torma; labrum with 10 sensilla: sensilla 1–4 an anterodorsal group, with sensillum 1 a peg in pit, sensillum 2 a styloconicum on cuticular projection, sensilla 3, 4 lobeshaped with 4 longer than 3, sensillum 5 an elongate seta, sensilla 6, 7 basally stout basiconica, sensilla 8, 9 laterally placed, sensillum 8 short seta, sensillum 9 a short peg, sensillum 10 a stout seta; ventral margin with well developed scopae, 18–21 moreorless uniform teeth (three lateral teeth more stout) in undivided row. Mandible ( figs. 2I View Fig , 3E View Fig , 10A View Fig ) curved, apical half tapering to sharp, darkly pigmented, point, with dorsal and ventral grooves; with large to very small lobe (not visible in some specimens) on inner surface; with subbasal seta and minute peg in pit on outer margin; with well developed apodeme attached to dorsal margin of mandible, extending into posterior half of head capsule. Maxilla ( figs. 10B View Fig , 16B View Fig , 17D View Fig ) well developed, wide; palpus elongate; large lobelike, apically tapering plate dorsal to base of palpus; with posteromedial elongate seta; palpus with 4 apical or subapical sensilla, two lateral sensilla each with elongate slender nib; group of 7 thick sensilla lateral to palpus: sensillum 1 extending to about half length of palpus; sensillum 2 with rounded apex; sensillum 3 short, arising near base of sensillum 4; sensillum 4 with elongate apical nib; sensillum 5 on elongate cuticular extension; sensillum 6 with rounded apex, ribbed; sensillum 7 with rounded apex; lateral seta elongate. Pharyngeal complex ( figs. 2J View Fig , 3F View Fig ) well developed, epipharynx with two lateral arms, apparently lacking combs (present but difficult to discern in some other Ceratopogonidae ). Hypopharynx with lateral arms articulating with lateral apices of lateral arms of epipharynx. Thorax, abdomen: Cuticle unpigmented, transparent, thin. Prothorax secondarily divided, with well developed cervix; with elongate proleg ( figs. 2C–G View Fig , 4B View Fig , 7B View Fig , 12B View Fig , 17A View Fig , 18 View Fig ), with apical hooks, 5–6 (per half) anterior terminal hooks elongate, posterior hooks short; proleg capable in life of being withdrawn into prothorax (posterior to cervix). Segment nine ( figs. 3B, H View Fig , 11A, B View Fig ) with wellseparated (with bases not closely ap proximated) setae: dorsal setae o, i, l 1, d, l 2, l 3, l 4, ventral setae i, o, l 1, l 2, v; posterior proleg ( figs. 2C–G View Fig , 3B View Fig , 11A, B View Fig , 17B View Fig ) a single posterior structure with about 15–20 well developed hooks, dorsal hooks with broader bases than ventral, more slender hooks; ventral hooks with spicules; proleg capable of being extruded or withdrawn into body cavity. Four anal papillae, each apically bifur cate. Midgut white, with annulations (obscured by fat body in some third and fourthinstar larvae); anterior margin situated at midlength of fourth true abdominal segment (at anterior margin of apparent segment 8 in larvae with secondarily divided segments). With two Malpighian tubules.
Firstinstar larva: Head capsule ( figs. 7A, B View Fig , 8A View Fig ): With dorsal, darkly pigmented egg burster ( figs. 2H View Fig , 7A, B View Fig , 8B View Fig ). Without eyespot. Mandible ( fig. 2I View Fig ) with large triangular tooth on inner margin (not evident in some specimens). Hypostoma lacking teeth ( figs. 2K View Fig , 8A View Fig ) Abdomen: With or without segments 1–8 secondarily divided (so abdomen appears to have either 9 or 17 segments) ( fig. 2C, D View Fig ). Abdominal segment 9 ( fig. 11A, B View Fig ) with at least dorsal setae o, i, d, l 1, 1 2, 1 3, ventral setae o, i present; others not visible but may be present; dorsal seta o notably thicker, longer than other setae on segment. Hemolymph unpigmented.
Secondinstar larva: Head capsule: With or without eyespot. Mandible likely (not clearly visible) with moderately sized triangular tooth on inner margin. Hypostoma with well developed row of teeth. Abdomen: With segments 1–8 secondarily divided (so abdomen appears to have 17 segments) ( fig. 2E View Fig ). Abdominal segment 9 with uncertain number of setae; dorsal seta o at least slightly thicker than other setae on segment. Hemolymph unpigmented or very pale pink.
Thirdinstar larva: Head capsule: With eyespot. Mandible with small bump on inner margin. Hypostoma ( fig. 19 View Fig ) with well developed row of teeth, central tooth largest. Abdomen: With segments 1–8 secondarily divided (so abdomen appears to have 17 segments) ( fig. 2F View Fig ). Abdominal segment 9 with 7 dorsal, 5 ventral setae distributed as in figures 3H View Fig , 17B View Fig ; dorsal seta o at least slightly thicker than other setae on segment. Hemolymph unpigmented or pink. Some fat body visible in more mature larvae.
Fourthinstar larva: Head capsule: With eyespot. Mandible ( fig. 3E View Fig ) with small bump on inner margin. Hypostoma ( figs. 3G View Fig , 16A View Fig ) with well developed row of teeth, central tooth largest. Abdomen: With segments 1–8 secondarily divided (so abdomen appears to have 17 segments) ( fig. 2G View Fig ). Abdominal segment 9 with 7 dorsal, 5 ventral setae distributed as in figures 3H View Fig , 17B View Fig ; dorsal seta o equal in diameter to other long setae on segment. Hemolymph pink or reddish. Fat body present.
Pupa: Only pupa known is of A. mcmillani , described below.
DISTRIBUTION AND BIONOMICS
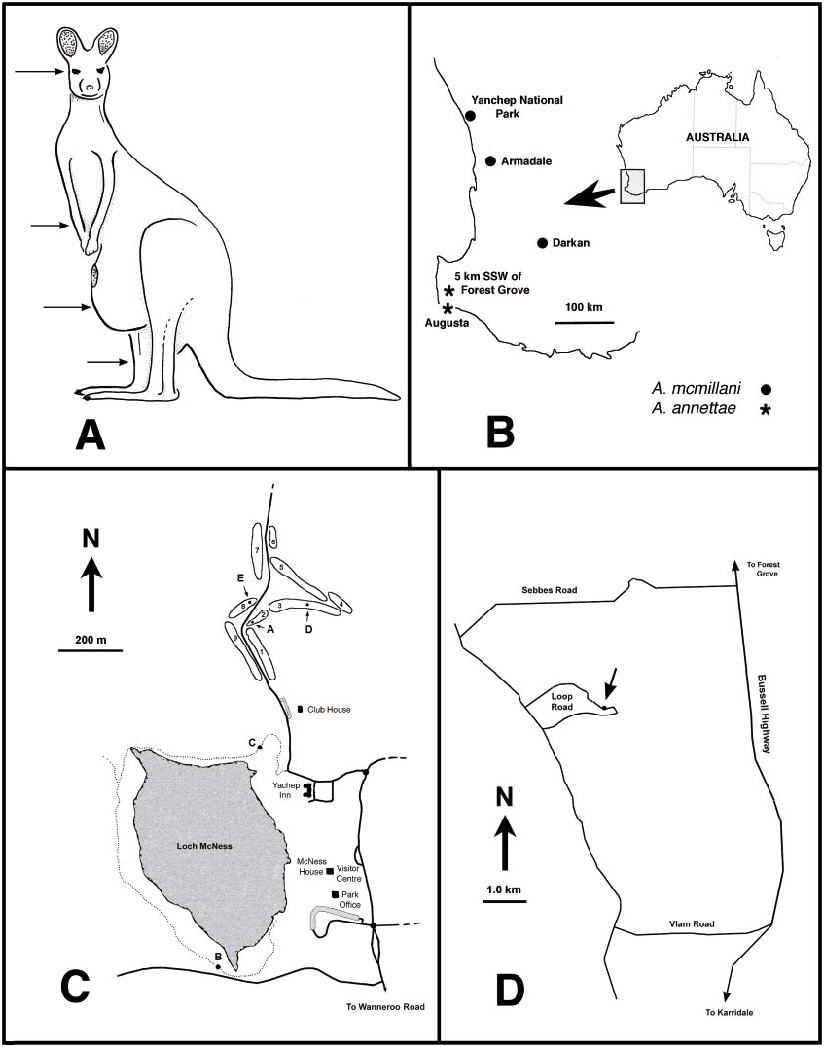
The only two extant species are restricted to southwestern Australia ( fig. 22B View Fig ), but Cretaceous fossils are known from France, Spain, Siberia, Lebanon, and Myanmar ( Borkent, 2000a; Szadziewski, in press; Szadziewski and Arillo, 2001), proving that the genus was once much more broadly distributed.
The only species in which females have been observed to bite are those of A. mcmillani , which feed on kangaroos and humans. However, the finely serrate mandible and retrorse lacinial teeth of the adult females of A. annettae strongly indicate that these too feed on vertebrates ( Borkent, 1995: 129– 132). The morphology of the claws of the female of A. annettae additionally may indicate that these feed on birds (see discussion below under that species). The mouthparts of fossil Austroconops are not visible ( Borkent, 2000a), and therefore it is uncertain what they fed upon. The presence, however, of vertebrate bloodfeeding in A. mcmillani and the phylogenetic position of the genus as an early lineage within the family where vertebrate bloodfeeding is plesiotypic strongly suggest that all extinct species fed on vertebrates ( Borkent, 1995, 2000a). Details of male swarming in A. mcmillani are given below.
Firstinstar larvae of A. annettae and A. mcmillani were both successfully reared to fourthinstar larvae (and A. mcmillani to the pupal stage) in very wet soil, with regular (generally every second day) additions of nematodes and a fecal infusion. All instars of the aquatic larvae were clearly attracted to fresh drops of fecal infusion as shown by the concentrations of larvae directly under these drops and they were rarely seen to feed on nematodes. This phenomenon, and the presence of well developed, finely toothed scopae ( figs. 3A View Fig , 19 View Fig ) suggest that they are likely associated with feces or concentrated decomposing vegetation in nature (producing an abundance of microorganisms). Some second, nearly all third, and all fourthinstar larvae had pink or red hemolymph, indicating the presence of hemoglobin, which additionally suggests that they may be associated with an oxygen deficient wet habitat. Larvae cannot swim but use a combination of their anterior and posterior prolegs and, especially in later instars, a relatively slow serpentine body motion to move through wet substrate. Further details of behavior and habitat are provided below for each species.
Borkent et al. (1987) suggested that the bluegreen pigmentation of live adult A. mcmillani may have indicated that the then unknown larvae were feeding on algae because this coloration has been observed in some Ceratopogonidae such as some Culicoides species of the schulzei species group and some Dasyhelea Kieffer species which feed on algae as larvae. Our observations here show that the relationship between algal feeding and adult color is not substantiated for at least A. mcmillani as larvae matured to adulthood without feeding on algae. Austroconops annettae was also successfully reared to the fourthinstar without algae.
TAXONOMIC DISCUSSION
We consider the elongate, pale egg of Austroconops species unique within the family. Eggs laid by other Ceratopogonidae are initially pale but soon turn dark. However, of 103 extant genera, eggs have been described for only 18 genera and of those, 4 genera are known only as eggs described from within the female abdomen (so their final color cannot be determined).
We identified the four larval instars of both species based on the following evidence. Firstinstar larvae are easily identified by their darkly pigmented egg burster ( figs. 2H View Fig , 7A, B View Fig , 8B View Fig ). Because of the small number of measured second and fourthinstars of reared Austroconops larvae it was initially difficult to identify the instars 2–4. The fourthinstar of A. mcmillani was confidently identified because one of these molted to the pupal stage. Although the head capsule of this individual was not retrieved from the mud and therefore not measured, observations before it pupated showed that it was clearly close to, or within, the range of measurements reported here. Head capsule length increments of all instars followed Dyar’s Law, increasing by a factor of 1.32, 1.31, and 1.34 for A. mcmillani and 1.33, 1.32, and 1.25 for A. annettae . The latter value, the factor of change for third to fourthinstar, may be due to the small sample size of fourthinstar A. annettae or perhaps to less than optimal feeding conditions (and hence smaller individuals). These values are similar to those reported for species of Culicoides , which are about 1.4 ( Kettle and Lawson, 1952; Kettle and Elson, 1975).
There are significant statistical differences in egg characteristics (table 3) and larval head capsule lengths (table 4) of the different instars between A. mcmillani and A. annettae . Considering, however, that these immatures were obtained from eggs laid by a very few females, and that the larvae were reared under laboratory conditions, the size differences noted here may be artifacts. Otherwise, eggs and larvae of the two species could not be distinguished from one another.
Larvae of Austroconops , when compared to other Ceratopogonidae , are missing head capsule sensilla z, k (sensory pit), and q. Also, there are two additional dorsolateral setae on larval abdominal segment 9 ( fig. 3H View Fig ), here labeled as l 3 and l 4, which have not been reported in other Ceratopogonidae .
The larval antennal sensilla of Ceratopogonidae have never been described in sufficient detail to determine most homologies within and outside the family. Larvae of Austroconops species have a welldefined basal segment bearing 6 apical sensilla and a further second and third segment ( figs. 9A, B View Fig , 14A, B View Fig , 20A View Fig ). Based on position, relatively large size, and a uniformly porous surface, the most elongate, porous sensillum is homologous to the ‘‘large lobe’’ present in Culicoides ( Murphree and Mullen, 1991) and most other Ceratopogoninae ( Borkent and Bissett, 1990; Borkent and Craig, 1999). Based on position and some details of structure, the following sensilla are likely homologous to sensilla in Chironomidae : most elongate, porous sensillum = blade; mesal short sensillum = accessory blade; short bilobed sensillum labeled sensillum 3 here = fused style and peg sensillum. The remaining three short sensilla do not have readily apparent homologies with Chironomidae ; they are labeled here as sensilla 1, 2, and 4. Sensillum 1 has an open apex bearing a tiny peg. Sensillum 4 has a porous surface.
The labrum of Austroconops shows a number of similarities to other Ceratopogonidae . Sensilla 1–4 are clearly present as a group on the labra of most other Ceratopogonidae ( Hribar and Mullen, 1991, labeled as ‘‘sensillum styloconicum’’). In many Chironomidae the two sensilla SIVA amd SIVB on the anterior portion of the labrum are morphologically very similar to S2 and S4, and these are likely homologous. Chironomidae S1, just dorsal to the labral lamellae, probably is homologous to our S10, and the scopae present in Austroconops and Ceratopogoninae are probably homologous to the labral lamellae (Wiederholm, 1983). Further detailed comparisons require further study of these structures in numerous taxa (both SEM and study of nerves).
Borkent et al. (1987) reported an ‘‘Mshaped apodeme’’ in the female genitalia of A. mcmillani . In further study we are puzzled as to the nature of this structure, which varies in both A. mcmillani and A. annettae from a faint Mshaped sclerotization to the presence of a pair of laterally positioned, pigmented sclerites. Some other Ceratopogonidae also have sclerotized structures anterior to the fused or separated sternite 9 (e.g., some Leptoconops , some Culicoides , Alluaudomyia Kieffer , and others). Careful histological study is needed to better interpret this feature.
Cladistic analysis below indicates that, among extant taxa, Austroconops is the sister group of Leptoconops and that together they form the sister group to all remaining extant Ceratopogonidae . Eight species of Austroconops are now known: two of these are extant and six are Cretaceous fossils:
A. annettae View in CoL , n.sp. Borkent, this publication. Australia (Western Australia)
A. borkenti Szadziewski and Schlüter, 1992: 78 View in CoL . France. Upper Cretaceous
A. fossilis Szadziewski, 1996: 38 View in CoL . Lebanon. Low er Cretaceous
A. gladius Borkent, 2000a: 378 View in CoL . Lebanon. Lower Cretaceous.
A. gondwanicus Szadziewski, 1996: 38 View in CoL . Lebanon. Lower Cretaceous
A. mcmillani Wirth and Lee, 1958: 337 View in CoL . Australia (Western Australia)
A. megaspinus Borkent, 2000a: 381 View in CoL . Lebanon. Lower Cretaceous.
A. sibericus Szadziewski, 1996: 40 . Russia. Upper Cretaceous
The genus Jordanoconops Szadziewski was proposed as a monotypic genus to include a Lower Cretaceous Jordanian amber fossil, Jordanoconops weitschati Szadziewski ( Szadziewski, 2000) . It may actually be a member of Austroconops (see below under







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Austroconops Wirth and Lee
| BORKENT, ART & CRAIG, DOUGLAS A. 2004 |
A. gladius
| Borkent, A. 2000: 378 |
A. megaspinus
| Borkent, A. 2000: 381 |
A. fossilis
| Szadziewski, R. 1996: 38 |
A. gondwanicus
| Szadziewski, R. 1996: 38 |
A. sibericus
| Szadziewski, R. 1996: 40 |