
Trisunius Assing, 2011
|
publication ID |
https://doi.org/ 10.1206/0003-0090.460.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/FA4F87B1-E182-FF63-09D7-C0B6FE9BFBF9 |
|
treatment provided by |
Felipe |
|
scientific name |
Trisunius Assing |
| status |
|
Trisunius Assing View in CoL , New Subtribal Assignment
Figures 16 View FIGS , 22 View FIGS , 258 View FIGS , 326–348 View FIGS View FIGS View FIGS View FIGS View FIGS
Trisunius Assing, 2011: 196 View in CoL View Cited Treatment . Type species: Trisunius spathulatus Assing, 2011: 198 View in CoL View Cited Treatment , fixed by original designation.
— Assing, 2012a: 210 (partially revised key to species). — Assing, 2013: 1539 (checklist of species with general distribution). — Assing, 2014 (new records and species).
— Assing, 2015 (new records and species).
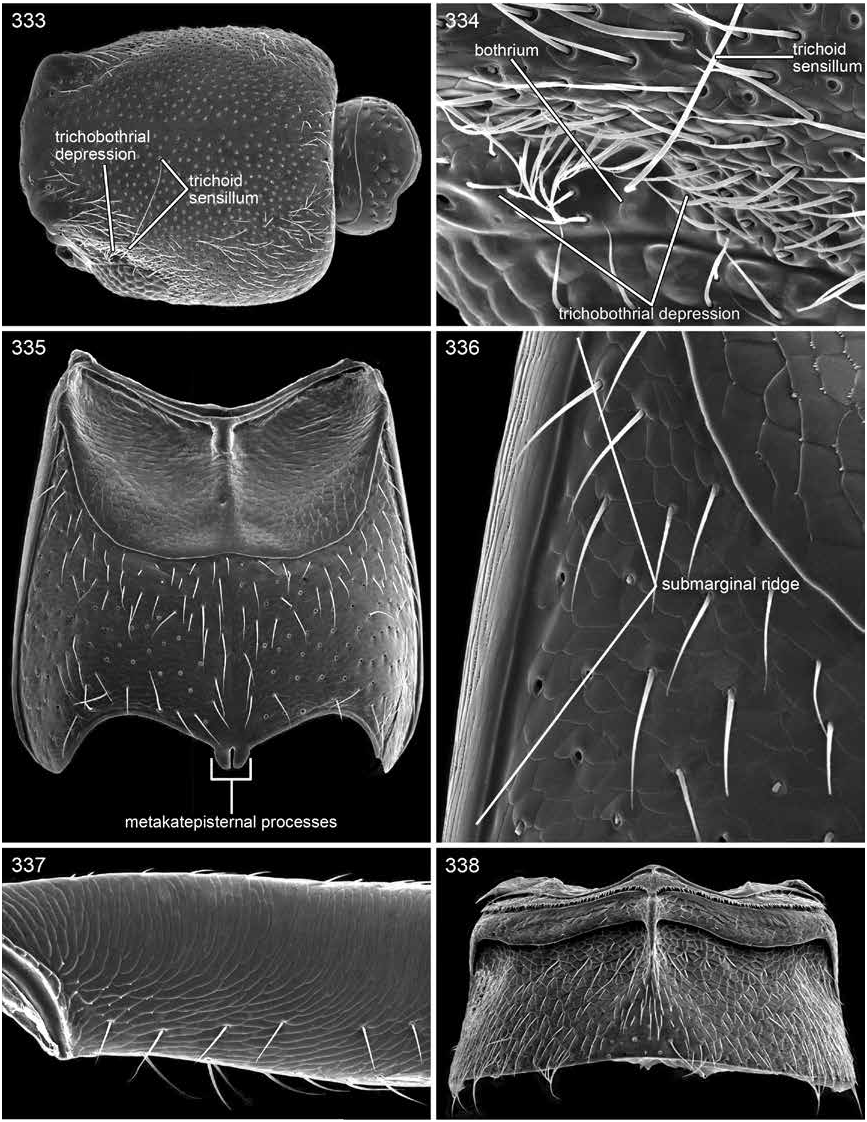
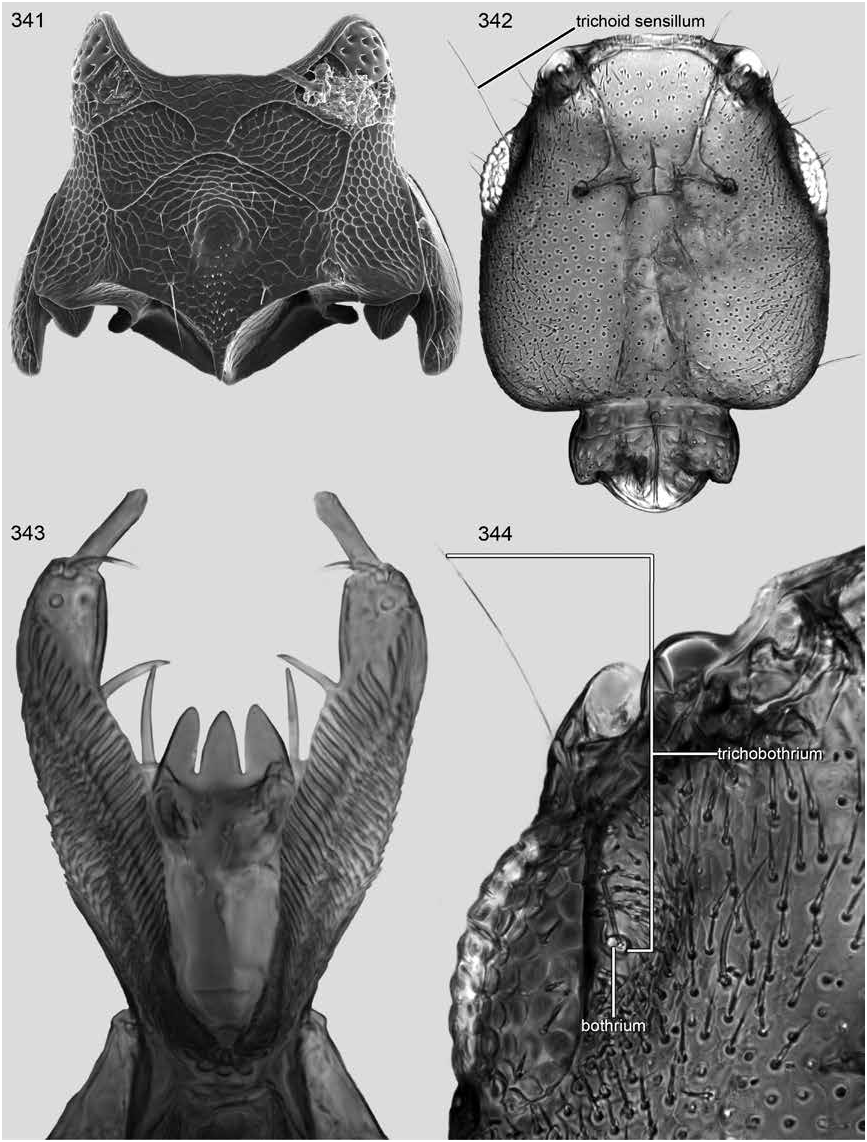
DIAGNOSIS: Trisunius is separated from Scopaeus by a wide neck (fig. 342), absence of a stridulum (figs. 335–337), apically rounded metakatepisternal process (fig. 335), and presence of the pericoxal ridge around the mesocoxal acetabulum (fig. 335). The wide neck of Trisunius will separate it from Hyperscopaeus (fig. 240); the neck of the former is about a third to two fifths as wide as the postocular width of head, the latter about one eighth to one sixth. Orus and Micranops are both separated from Trisunius by the position of the cephalic trichobothrium. The trichobothrium of Trisunius is in a trichobothrial depression near or just forward of the middle of the dorsal edge of the eye, and the sensillum is near the middle of the eye (figs. 344, 333); for Orus it is in a trichobothrial canal that begins at the dorsoposterior edge of the eye, which tapers posteriorly and the sensillum is adjacent to the posterior margin of the eye; for Micranops the trichobothrium is in a short trichobothrial cavity behind and separated from the eye.
DESCRIPTION: Body length 2.0–4.1 mm.
Head (fig. 342) with postocular lateral margin broadly and shallowly rounded; basal angles well developed and strongly rounded; basal margin shallowly emarginate medially and without median tumescence, groove, or sulcus; posteroventral surface without tubercles.
Neck (fig. 342) not petiolate; nuchal groove moderately deep; neck width across nuchal constriction a third to two fifths as wide as postocular width of head; nuchal ridge present dorsally and laterally (figs. 333, 342).
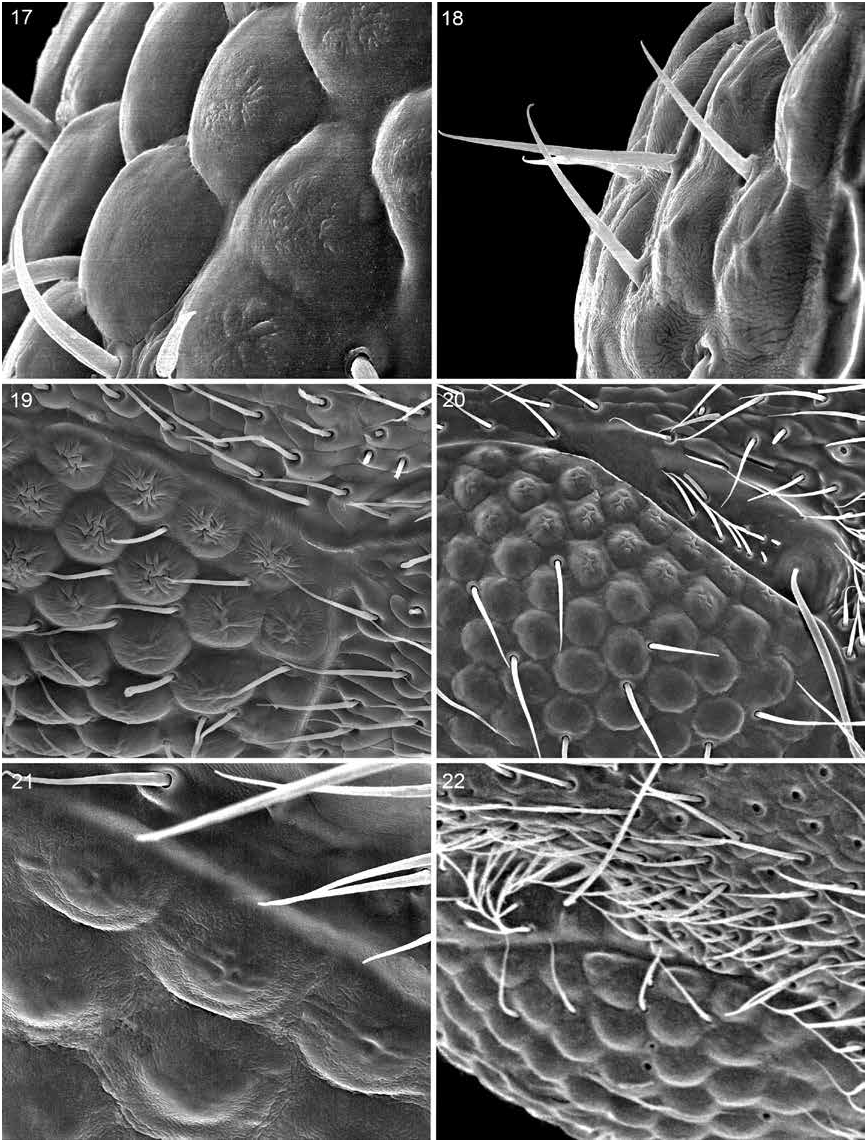
Dorsal surface of head with fine to moderately strong, moderately dense to dense punctation (fig. 333); midlongitudinal strip without or with less dense punctation; microsculpturing fine, strong, and dense to feeble to absent; surface with dull to polished luster; pubescence fine and moderately dense to dense; macrosetae moderately long, peripheral, and few.
Cephalic trichobothrium in trichobothrial depression adjacent dorsal margin of eye near or just forward of middle of eye (fig. 334); trichobothrial depression (fig. 334) with small cluster of setae in front of bothrium and with dense cluster of setae along dorsal margin.
Eyes with broadly rounded posterior margin.
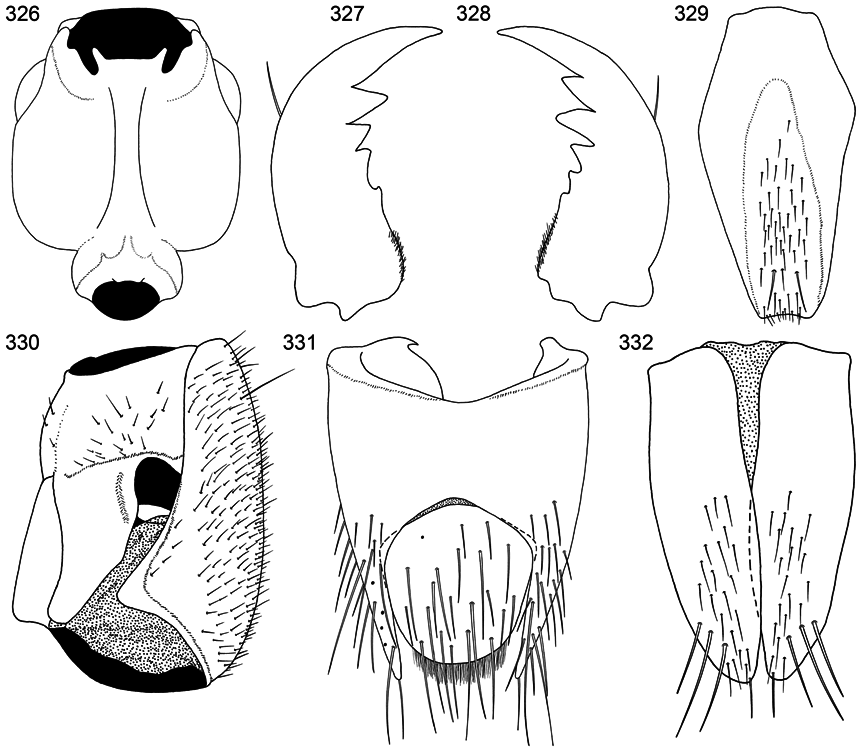
Gular sutures moderately widely separated and divergent posteriorly (fig. 326).
Mandibles: left mandible with three ( Assing, 2011: fig. 71) or four denticles (fig. 327); right mandible with three ( Assing, 2011: fig. 72) or five denticles (fig. 328).
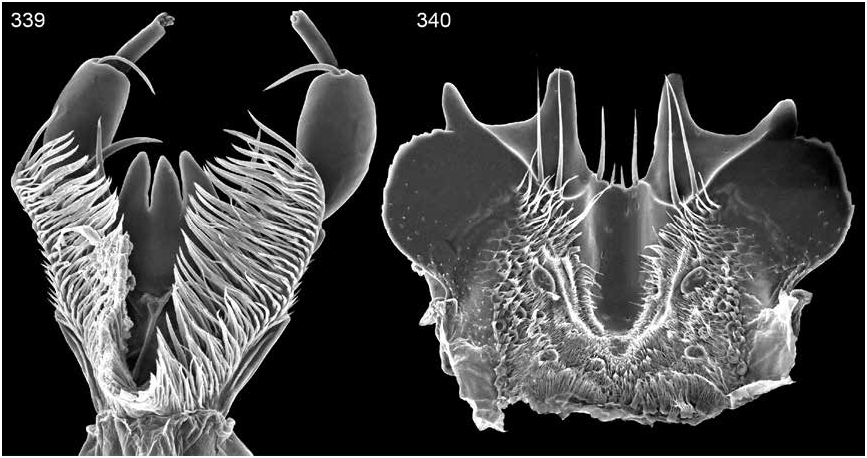
Labrum quadridentate (figs. 340).
Pronotum broadly and moderately convex; surface with moderately dense to dense punctation; microsculpturing well developed, poorly developed, or absent and uniform to present laterally and/or anteriorly; surface moderately shiny to polished; pubescence moderately dense to dense; macrosetae moderately long, peripheral, and few; midlongitudinal strip impunctate and without microsculpturing or with weak to feeble microsculpturing anteriorly; median groove absent or present posteriorly and moderately strong to weak; midlongitudinal ridge present and weak posteriorly or absent; posterior margin weakly emarginate to straight.
Prohypomeron without transverse hypomeronal ridge or ridge present, weakly developed, and incomplete; submarginal ridge present; lobe with or without setae or with a few (fig. 330).
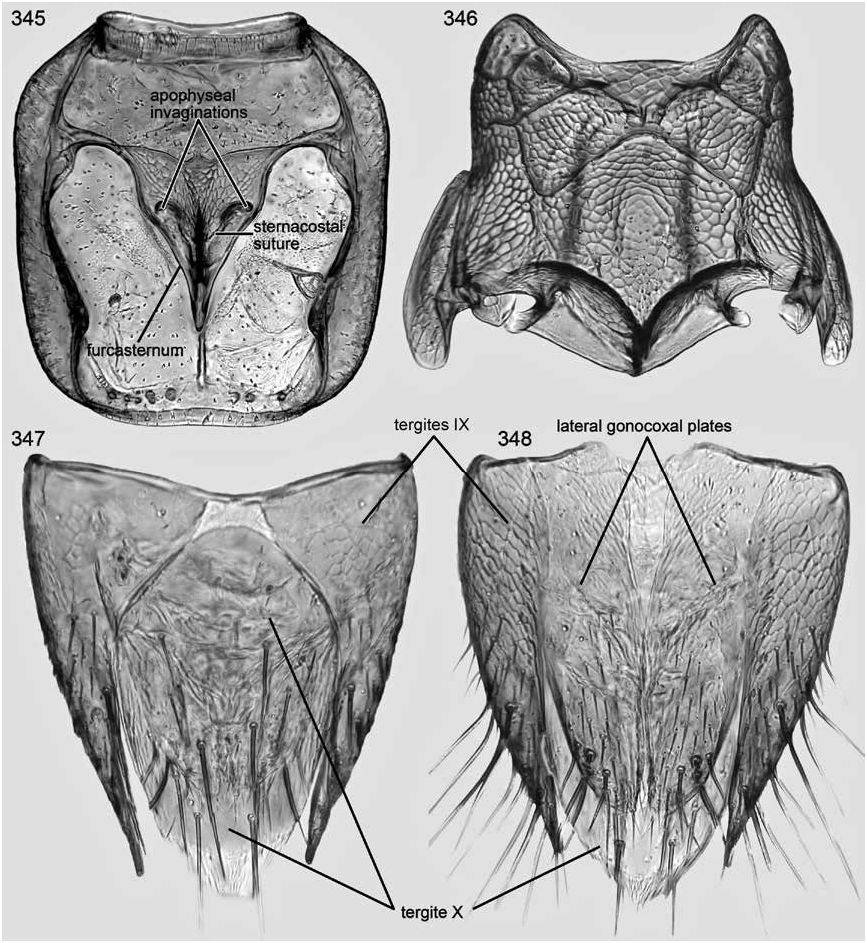
Notosternal suture present (fig. 330, 345).
Elytra longer to shorter than pronotum; surface with moderately dense punctation and pubescence; posterior margin with row of setae.
Mesoventrite with large, rounded, deep, median, basisternal depression (fig. 341); prepectal ridges separated medially; mesotransversal ridge broadly curved; mesanapleural ridge with anterior segment present, posterior segment absent.
Mesofurcasternum without (fig. 346) internal, median apophysis on posteromedial margin.
Mesofemur without stridular plectrum (figs. 335–337).
Mesocoxal acetabulum margined by pericoxal ridge (fig. 335) or present only laterally.
Metaventrite without stridular file (fig. 336). Metakatepisternal process short and apically rounded (fig. 335).
Sternite II with median point on posterior margin (fig. 338).
Sternite III with well developed median carina; transverse basal ridge moderately sinuate and with long median point.
Male. Sternite VII with or without slightly to moderately modified surface and with or without shallow emargination of posterior margin (see Assing, 2011: figs. 6, 13, 23, 30, 37, 43, 48, 54, 63, 77).
Sternite VIII with emargination of posterior margin of variable width and depth (see Assing, 2011: figs. 7, 14, 24, 31, 38, 44, 49, 55, 64, 78).
Tergite IX (fig. 331) asymmetrical; left anteroventral side larger, more broadly rounded, and wrapping ventromedially more than right; posterior emargination more or less U-shaped; middorsal base fused. Sternite IX (fig. 329) elongate and moderately wide.
Tergite X elliptical; anterior margin without median point, strongly rounded medially, then sloped and slightly rounded to lateral margin (figs. 331).
Aedeagus with dorsal surface of median lobe midlongitudinally divided.
Female. Sternite VII unmodified.
Sternite VIII with unmodified surface and posterior margin.
Abdominal sternite VIII with rounded posterior margin.
Tergite IX (fig. 347) symmetrical; left and right anteroventral sides of approximately equal size; posterior emargination wide, deep posteriorly, narrow anteriorly, and extending to anterior margin of tergite; middorsal base medially divided.
Tergite X (fig. 347) with anterior margin strongly rounded medially, then sloped and slightly rounded to lateral margin.
DISTRIBUTION AND HABITAT: At this writing all the named species are from the Asian mainland. Ten of the 22 described species are from China. The genus is also known from Nepal, India, Thailand, and Vietnam. In China the genus is known from Yunnan, Hubei, and Shaanxi. Unnamed species from Vietnam, Malaysia, Nepal, the Philippines, and South Africa were examined.
Reflecting the intensity of collecting, most of the Chinese species are represented by many specimens; only one is known by just the holotype and two others by the holotype and one paratype. By contrast, most of the species from Nepal and Thailand are known by only the holotype and for India by a few specimens. Since species are found in Hubei and Shaanxi provinces perhaps other more northern and eastern species are yet to be discovered. Based on the occurrence of described species along with the data for a few unnamed species, it is plausible that many more species will be described and that the known geographic range of Trisunius will be significantly enlarged. The genus probably occurs throughout central and southern China, Southeast Asia , the Indian subcontinent, Maritime Southeast Asia , Africa , and perhaps Australia .
According to label data published by Assing (2011 – 2015) specimens have been collected primarily in forest litter, deep humus, wet debris near a waterfall at 850 m elevation, near snowfields, and occasionally in open grassland or shrub habitats. Species have been collected at elevations of 270 to 3600 m. The genus appears to be primarily montane. A specimen of T. spathulatus was collected at 850 m, but the species was generally found at elevations between 2000 and 2600 m. Most species and specimens were collected at elevations between 2000 and 2900 m. Individuals of 11 species were collected below 2000 m. High mountain species collected above 3000 m include T. perpusillus (3100 m), T. monticola (3470 m), and T. alesi (3600 m). Unnamed Vietnamese specimens were collected from near a forest stream at 270 m and from a drying stream at 1150 m and one from Borneo was found at 1500 m.
DISCUSSION: When Assing (2011) described Trisunius he included 10 species, only one of which was already known. To date Assing has described 20 of the 22 species; two were described in other genera. When Assing (2011: 214) proposed the genus he transferred the Indian species, Medon monticola Cameron, 1931 , to Trisunius and a year later added Lathrobium perpusillus Coiffait, 1982 ( Assing, 2012a: 210) .
Subtribal assignment. Assing (2011) included Trisunius in the inadequately defined Medonina . That subtribe is effectively an accumulation of leftover genera with a squarish pronotum that cannot be included in other subtribes and that may or may not be related. Trisunius has a supraocular trichobothrium, one of the principal defining features of the Scopaeina . However, some impalpable feature also suggests Scopaeina . When I first saw a specimen of the type species in Berlin I immediately thought “ Orus ” then looked for the trichobothrium. Since most of the species of the Scopaeina , most notably the speciose Scopaeus , have a narrow, petiolate neck, many workers simply do not consider the Scopaeina to be home to species with a wide neck. Not only does Trisunius share the paraocular cephalic trichobothrium with other scopaeines, but it also bears the tripartite ligular lobe, trilobed abdominal sternite II, and it lacks a pronotal marginal ridge.
FLIGHT: Of the 22 species presently described nearly half are reported to have shortened elytra with weakly developed humeral angles, wings that are reduced or “completely” reduced, and a palisade fringe of tergite VII that is present, rudimentary, or absent. Four species, T. cultellatus , T. discrepans , T. ligulatus , and T. spathulatus , have a palisade fringe and are considered dimorphic for elytral and wing length ( Assing, 2011: 198, 202, 203, 206). Furthermore, the eyes are reduced for those species. Assing (2011: 198) wrote, not only do dimorphic species have modifications of the elytra and wings, but the head of the micropterous and macropterous individuals differ. The modified head of the two forms was most clearly illustrated for T. cultellatus ; the head of the micropterous morph is weakly dilated behind the eyes, whereas the head of the macropterous form is subparallel to narrowed posteriorly ( Assing, 2011: 203, cf. figs. 18, 19). Three species, T. alesi , T. rastratus , and T. smetanai , are reported to have a narrow or rudimentary palisade fringe; these species may have yet undiscovered flying individuals ( Assing, 2012a; 2013; 2014: 446). On the other hand, because they lack a palisade fringe, three species, T. borailicus , T. manasluensis , and T. perpusillus , flying individuals may be uncommon; for each the wings are completely reduced, however, each is known only by the holotype ( Assing, 2012a: 211, 214; 2013: 1535).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Trisunius Assing
| Herman, Lee 2023 |