
Coryphomys musseri, Aplin, K. P. & Helgen, K. M., 2010
|
publication ID |
https://doi.org/ 10.1206/692.1 |
|
persistent identifier |
https://treatment.plazi.org/id/FA68571D-312D-FFBD-FC9B-330591D886FB |
|
treatment provided by |
Tatiana |
|
scientific name |
Coryphomys musseri |
| status |
sp. nov. |
Coryphomys musseri , sp. nov.
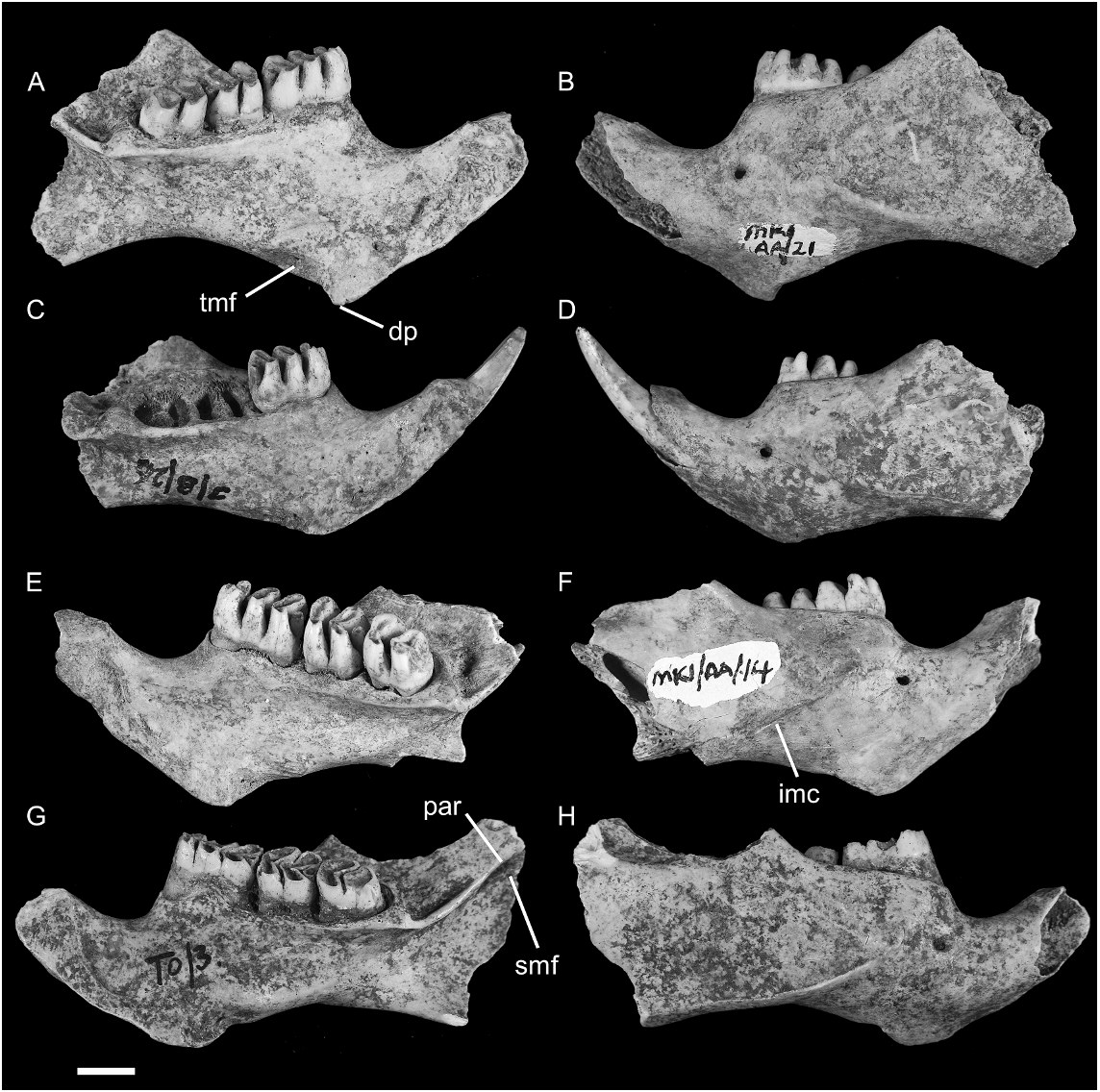
HOLOTYPE: ANWCP32 : Matja Kuru 1 ( MK1 /AA/21), an unburnt left dentary with m 1–3 in an early stage of wear ( figs. 11O View Fig , 16A–B View Fig ). The incisor is missing from its alveolus. The pterygoid, coronoid, and articular processes of the dentary are broken away. The specimen was excavated from Spit 21 of Square AA at the cave site of Matja Kuru 1 by S. O’Connor, and is understood to be early Holocene in age.
REFERRED SPECIMENS: AMF 68765: Uai Bobo 1 (TO2/G), L Dentary with m1–3; AMF 68851: Uai Bobo 1 (TO1/3), R Dentary with m1–3; AMF 68753: Uai Bobo 1 (TO1/3) R Dentary with m1–3; AMF 68812: Uai Bobo 1 (TO1/3), R Dentary with m1–3; ANWCP 9: Matja Kuru 2 ( MK 2/D/44), R Dentary with m1–2; AMF 68752: Uai Bobo 1 (TO1/3), L Dentary with i 1 m 1–2; AMF 68763: Uai Bobo 1 (TO1/4), L Dentary with m1–3; AMF 68789: Bui Ceri Uato (TB/2), L Dentary with m1–3; AMF 13 CM 10: Uai Bobo 1 (TO1/G), L Dentary with m1–3; AMF 68789: Uai Bobo 1 (TO1/G), R m1; ANWCP 10: Matja Kuru 1 ( MK 1/AA/14), R Dentary with m1–3; ANWCP 11: Matja Kuru 1 ( MK 1/AA/16), L Dentary with m1–3; ANWCP 12: Matja Kuru 1 ( MK 1/AA/20), R Dentary with m1–3; ANWCP 13: Matja Kuru 1 ( MK 1/AA/22), R Dentary with m1– 2; ANWCP 14: Matja Kuru 1 ( MK 1/AA/11), R M1; ANWCP 15: Matja Kuru 1 ( MK 1/AA/ 8), R Dentary with m1–3; ANWCP 16: Matja Kuru 1 ( MK 1/A/15), L m1; ANWCP 17: Matja Kuru 1 ( MK 1/AA/19), L m1; ANWCP 18: Matja Kuru 2 ( MK 2/D/43), R m1–3; ANWCP 19: Matja Kuru 1 ( MK 1/A/ 28), L M1; ANWCP 20: Matja Kuru 1 ( MK 1/ A/13), R Dentary with m1–2; ANWCP 21: Matja Kuru 1 ( MK 1/AA/12), R Dentary with m1–3; ANWCP 22: Matja Kuru 1 ( MK 1/AA/12), R m2; ANWCP 23: Matja Kuru 1 ( MK 1/A/29), L m2; ANWCP 24: Matja Kuru 1 ( MK 1/AA/18), R M1; ANWCP 25: Matja Kuru 1 ( MK 1/A/29), L M1; ANWCP 26: Matja Kuru 1 ( MK 1/A/31), R and L Maxillae with RM 2 and LM3; AMF 68770: Uai Bobo 1 (TO1/4), L Maxilla with M1–3; AMF 68756: Uai Bobo 1 (TO1/3), R Maxilla with M1; AMF 68768: Uai Bobo 1 (TO1/F), R Maxilla with M1; ANWCP 27: Jerimalai ( JM /B/26), L Dentary with i1 and m1; ANWCP 28: Matja Kuru 1 ( MK 1/AA/8), L m1 and L m2; ANWCP 29: Matja Kuru 1 ( MK 1/A/10), R m1; AMF 68831: Uai Bobo 1 (TO1/I), edentulous left dentary.
DIAGNOSIS: Differs from C. buehleri in having molars slightly smaller on average in actual dimensions and proportionally much smaller relative to osseous structures; M1–3 with smaller posterior cinguli; i1 with concave dorsolabial surface; m1 with anterocentral cuspid undivided in majority of specimens; m2–3 with smaller posterior cinguli; maxilla with anteriorly shallower medial palatal suture and longer antemolar region,
with superficial masseteric fossa located significantly further forward relative to M1; and dentary with more prominent digastric process, stronger inferior masseteric ridge, a larger fossa for deep portion of transverse mandibular muscle, and a postalveolar ridge that runs dorsal to superior mandibular foramen.
ETYMOLOGY: We take the greatest pleasure in naming this species after Guy Musser, in appreciation of his meticulous and inspirational studies of murine rodents.
DESCRIPTION
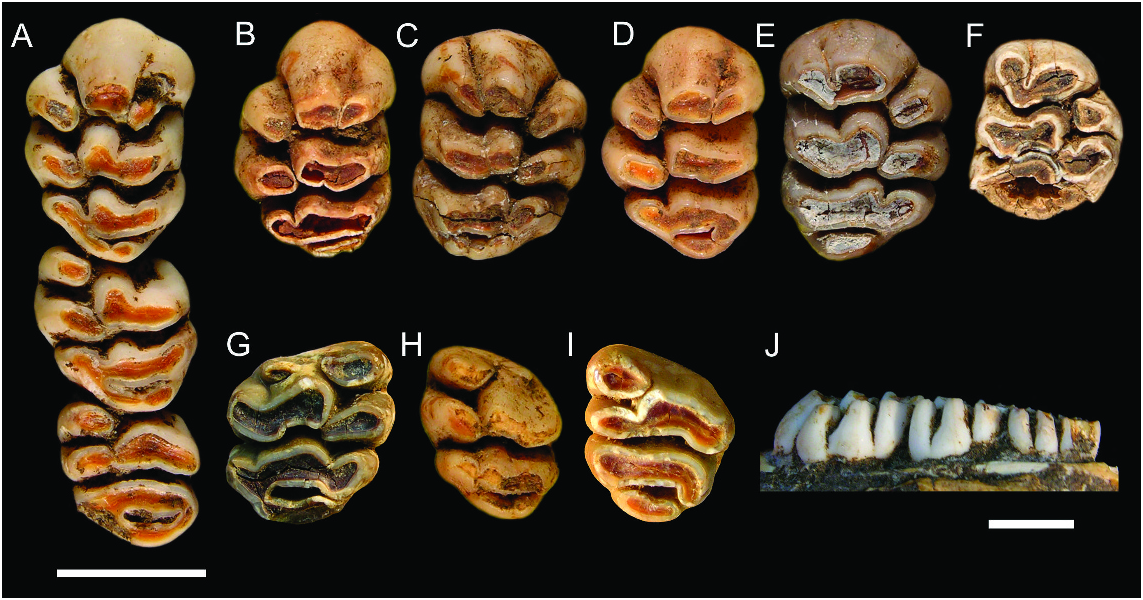
UPPER MOLARS ( fig. 10 View Fig , table 2): The upper molar cusp and root arrangements compare closely with those of C. buehleri . The root pattern of M2, as illustrated by ANWCP26, is shown to consist of a broad, kidney-shaped anterior root, supporting the anteroloph and with its concavity facing to the rear, a small circular root on the lingual side of the tooth, supporting cusps t4 and t7, and a larger circular posterolabial root, supporting cusps t8 and t9 and the associated posterior cingulum. The M3 of ANWCP26 has two roots and lacks the accessory labial root seen in C. buehleri .
M1: This tooth is represented by a total of six specimens ( fig. 10A–F View Fig ). The cuspal arrangement differs from that of C. buehleri in two respects: (1) each of the lingual cusps is positioned slightly further forward, creating a more transverse alignment of cusps in each lamina; and (2) the posterior cingulum is less bulky and more closely associated with the posterior surface of cusps t8–9.
As in C. buehleri , the posterior cingulum displays some variation in structure. On AMF 68770 ( fig. 10A View Fig ) it has a strong lingual connection to cusp t7 but is deeply separated from the rear of cusp t8 at the labial end; a small pimplelike cusp is present midway along the cingulum. On AMF 68756 ( fig. 10C View Fig ) the cingulum is an elevated ridge, deeply separated from each of cusps t7 and t8, and with three pimplelike cusps along its length and poor. Three other specimens have a simpler, lenticular posterior cingulum that is also deeply separated from each of cusps t7 and t8 ( fig. 10B, D–E View Fig ).
M2: Three examples are available ( figs. 10A, G–H View Fig ). Cusp t3 is entirely absent on AMF 68770 ( fig. 10A View Fig ), is represented by a small nubbin positioned near the crown base on another (ANWCP22; fig. 10H View Fig ), and is represented by a larger nubbin in the same position on the third (ANWCP24; fig. 10F View Fig ).
The posterior cingulum is a broader on M2 than on M1. In all three examples it shows a more elevated lingual connection to cusp t7 and a deeper labial separation from cusp t9. Two specimens have a pimple-like cusp at the labial end of the cingulum; the condition on the third is obscured by damage. Compared with C. buehleri , the posterior cingulum is less bulky and more closely approximated to the rear of cusp t8.
M3: Two specimens are available, both associated with an M2 ( figs. 10A, I View Fig ). The occlusal area of M3 appears to be smaller relative to M 2 in these specimens than in the few examples of C. buehleri . In part, this is due to the less bulky nature of the posterior cingulum, continuing the pattern observed on M1–2.
Neither M3 of C. musseri has any trace of cusp t3. The posterior lamina on AMF 68770 ( fig. 10A View Fig ) resembles that of C. buehleri with cusp t9 projecting backward to meet the elevated labial end of the posterior cingulum. On ANWCP26 ( fig. 10I View Fig ) cusp t9 is broadly united with cusp t8 and more deeply separated from the posterior cingulum.
LOWER INCISOR: Two dentaries preserve more or less complete lower incisors ( fig. 11 View Fig , table 2), but one of these is a young individual, with a correspondingly slender i1. Five others retain the base of a broken tooth. Although comparisons are difficult from a predominantly broken sample, the i1 appears to be slightly less robust than that of C. buehleri , due mainly to a more concave dorsolateral surface, and the medial surface appears to be flatter. The occlusal facet in the adult specimen is 10.52 mm in length and has a broadly concave posterior termination. The anterior margin of the facet curves to a medial point.
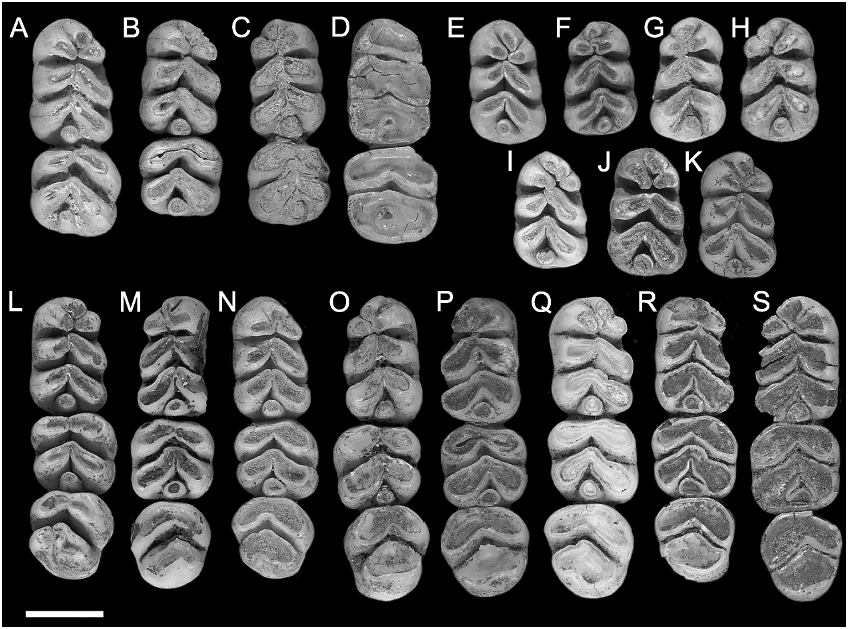
LOWER MOLARS ( fig. 11 View Fig , table 2): The complete lower molar series is retained in 12
TABLE 2
Measurements of Upper and Lower Dentition in Specimens Attributed to Coryphomys musseri
Measurements are molar lengths (L) and widths (W) and the breadth (IB) and depth (ID) of the lower incisor. Sample sizes (n), means, standard deviations (s.d.), and ranges are also shown.
3.31 4.76
3.29
5.01
IH 5.01 4.35 0.929 – 3.29 3
3.32
IB 2.88 0.666 1.91 – 4
W 5.69
M
3
5.1 4.98 4.92 5 5.14 0.263 – 4.8 14
L
3
5.61
M 5.61 5.23 4.8 5.18 0.331 4.5 – 14
W 5.78
M
2
5.32 5.68 5.12 5.59 5.32 4.91 5.2 5.32 0.236 4.91 – 20
L 5.77
2
) M
2 4.72 5.19 4.46 5.57 5.23 4.85 5.00 0.352 4.35 – 20 TABLE Continued W 1 5.05 – 23 (M 4.62 5 4.62 5 4.96 4.93 4.41 4.7 4.78 0.178 4.41
L 7.92
M
1
7.42 7.58 7.46 7.72 7.63 7.34 7.46 0.234 6.98 – 22
3
18.76
M
–
1
17.43 17.18 17.6 18.01 0.551 17.18 – 12
L R R L R L L R L
dentition No 3
Lower CP 32 13 CP CP 15 28 CP CP 14 23 CP 16 CP 21 CP Hooijer Mean.. d s Range N
dentaries, and six other specimens have two associated molars. Isolated lower molars are referred to this species on the basis of size and relative width. Molar root pattern appears identical to that described for C. buehleri except for the presence of a small accessory lingual root in two of 24 examples of the m1. Individual length and width measurements of m1 show slight overlap between C. musseri and C. buehleri (table 1– 2). Overlap is more extensive in dimensions of m2 and even more so for m3.
m1: This tooth is represented by 21 examples ( fig. 11A–S View Fig ), two of which are too heavily worn to show details of cuspal arrangements. The cuspal arrangement is essentially similar to that in C. buehleri but with the following minor differences: (1) the anterocentral cuspid is divided in two (10.5%) specimens (ANWCP20, fig. 11B View Fig ; and ANWCP29, fig. 11I View Fig ) (the cuspid is divided in 67% in C. buehleri , including the holotype and Hooijer’s specimen no. 2); (2) no specimen has small accessory cuspules attached to the rear of the anteroconid, though several (e.g., ANWCP13, fig. 11A View Fig ) have a buttresslike ridge extending forward from the base of the protoconid to produce a partial cingulum across the base of the anterolabial flexid (33% have accessory cuspules in C. buehleri ); (3) among the 16 examples where this relation can be scored, four (25%) show overlap of the hypoconid past the entoconid, 11 (69%) show the hypoconid and entoconid abutting without overlap, and one (6%) shows overlap of the entoconid in front of the hypoconid (60% of specimens show overlap of the hypoconid past the entoconid in C. buehleri ); and (4) the posterior cingulum is slightly smaller in all specimens.
Of 16 lesser worn examples of m1 that illustrate the relationship between the protoconid and metaconid, 11 (69%) show the two cusps abutting anteriorly without overlap, and five show the protoconid overlapping the front of the metaconid. The five specimens of C. buehleri (including Hooijer’s specimen no. 2) show two (40%) with overlap and three (60%) abutting without overlap.
The posterior cingulum is discrete in all but the most heavily worn example of m1 (ANWCP9, fig. 11D View Fig ), where it is united by a common dentin pool to the hypoconid.
m2: The m2, represented by 20 new specimens (e.g., fig. 11A–D, L–S View Fig ), is similar in proportions and cuspal arrangement to this tooth in C. buehleri . The main points of distinction other than smaller average size are the smaller size of the posterior cingulum and the less frequent suggestion of a fused or incipient anterolabial cuspid. The latter structure is moderately well indicated in two (14%) of the total of 14 specimens with an appropriate degree of wear, weakly indicated in four others (29%), and not expressed at all in eight (57%). In C. buehleri this structure is moderately well indicated in two specimens (40%) and weakly indicated in three (60%), including Hooijer’s specimen no. 2. A groove on the posterior surface of the protoconid may also be less frequent in C. musseri , being present in two specimens only ( ANWCP18 , fig. 11J View Fig ; and ANWCP10 , fig. 11L View Fig ) .
The posterior cingulum in the most heavily worn example of m2 (ANWCP9, fig. 11D View Fig ) is united by a common dentin pool to both the hypoconid and entoconid.
m3: This tooth is represented by 15 specimens (e.g., fig. 11L–S View Fig ). Among nine specimens with lesser degrees of wear, the anterior margin of the protoconid bears a clear fold, suggestive of an amalgamated anterolabial cuspid, in one example ( AMF 68812, fig. 11Q View Fig ) (11%) ; it is angular in four others (44%), and smoothly rounded in the remaining four (44%).
The posterior lamina varies considerably in width and cuspal morphology. Only one specimen of nine (11%) matches the condition described for C. buehleri , wherein anterior and posterolingual grooves clearly mark the union of the hypoconid and entoconid. Six specimens (55%) have a posterolingual groove in the same position but lack any anterior groove. In two specimens (22%) the posterolophid is more evenly divided by a posterior groove, with a small posterior cingulum at the base. The cingulum is best developed and most discrete in ANWCP12 ( fig. 11Q View Fig ) ; in ANWCP21 (not shown) it is united to the posterolingual surface of the hypoconid. Another specimen ( AMF 68789, not shown) shows the unusual feature of a small but distinct posterolabial cusplet, defined by a narrow groove on the anterolabial face of the hypoconid .
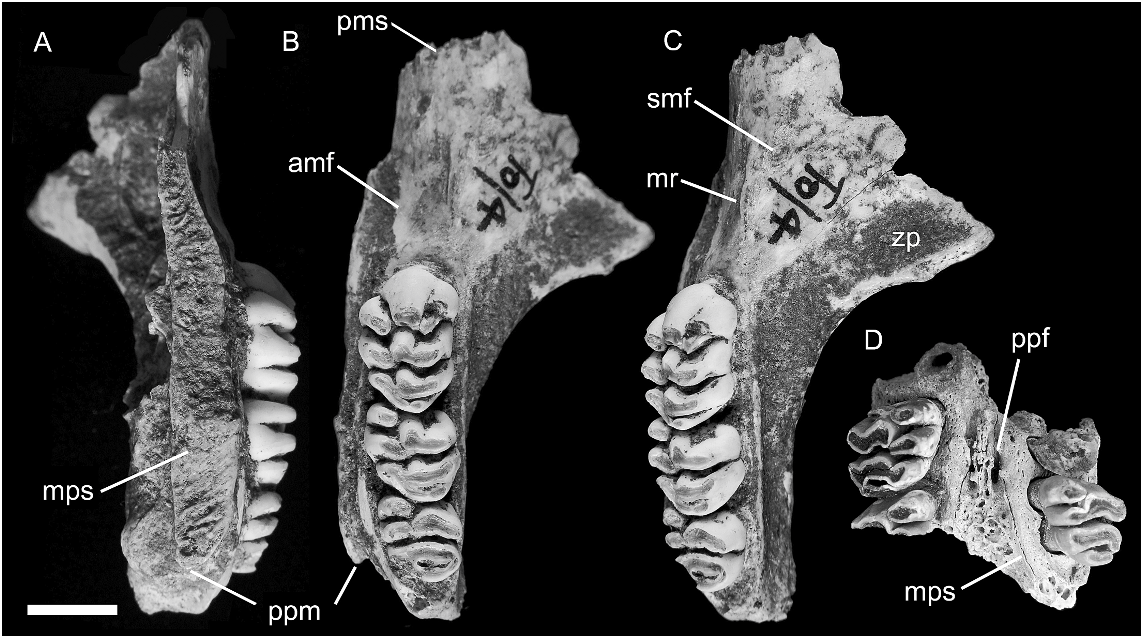
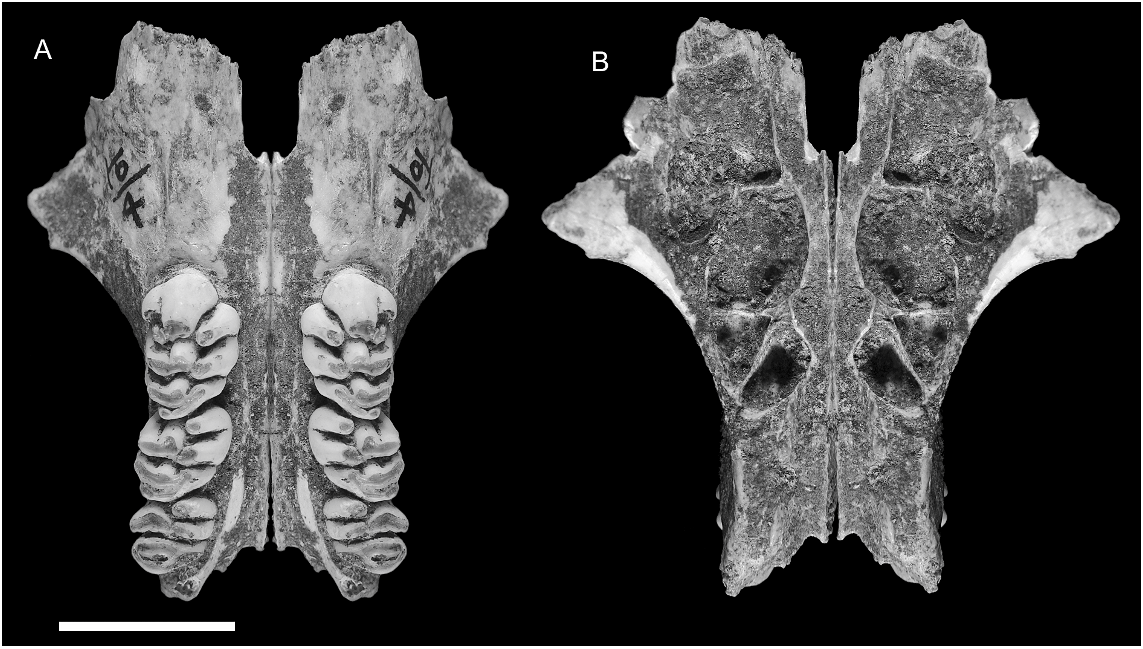
MAXILLA ( figs. 12–15 View Fig View Fig View Fig View Fig ): Partial maxillae of four individuals, two with attached portions of the palatine, together provide a fairly complete picture of palatal morphology in C. musseri . The most complete specimen (AMF 68770, figs. 12A–C View Fig , 13–15 View Fig View Fig View Fig ) illustrates the full length of the maxilla, from the suture with the premaxilla back to the posterior end of the alveolar portion, and retains all but the anterior portion of the midline palatal suture, including the palatine portion. The root of the zygomatic process is also preserved. The total length of the maxilla on AMF 68770 is reconstructed as 33.4 mm (essentially identical to C. buehleri ), with an M1–3 alveolar length of 17.3 mm (19.8 mm in C. buehleri ). This specimen is mirrored in figure 13 View Fig to illustrate the major features of the palate in comparison with C. buehleri .
Features shared with C. buehleri include: (1) the generally large size of the molar rows relative to the bony support structures and consequent narrowness of the palatal bridge that separates them; (2) the thickened nature of the palatal lamina (measuring 3.1 mm in depth at the medial suture between M1 and 3.9 mm between M2); (3) the short posterior palatal bridge (level with cusp t4 of M3); and (4) the morphology of the anterior palate which has a comparable midpalatal depression (antemolar palatal fossa) and lacks vascular grooving much beyond the anterior face of M1.
Key points of distinction between the two Coryphomys species (compared side by side in fig. 14 View Fig ) are the proportionally smaller size of the molars in C. musseri , relative to palatal structures, the greater degree of narrowing of the posterior nasopharyngeal margin (anterior margin of mesopterygoid fossa), and the proportionally longer antemolar palatal extent of the maxilla. Essentially, C. buehleri is a ‘‘megadont’’ version of C. musseri , with a degree of differentiation that greatly exceeds the bounds of normal intraspecific variation among murine rodents.
The anterior sutural contact with the premaxilla is located 13.1 mm forward of the anterior face of M1. Though damage to the anterior end of the maxilla on AMF 68770 and AMF 68756 has removed all trace of the anterior palatal foramen, it is clear that this was extremely short, probably much as in C. buehleri . The palate anterior to the molar row is considerably thinner in C. musseri than in C. buehleri , and this contrast is confirmed by a total of three specimens of C. musseri . The sutural contact between the palatine and maxilla, visible on the medial palatal suture of AMF 68770 ( fig. 12A View Fig ), is situated level with the embrasure between cusps t1 and t4 of M2, slightly forward of its relative position in C. buehleri .
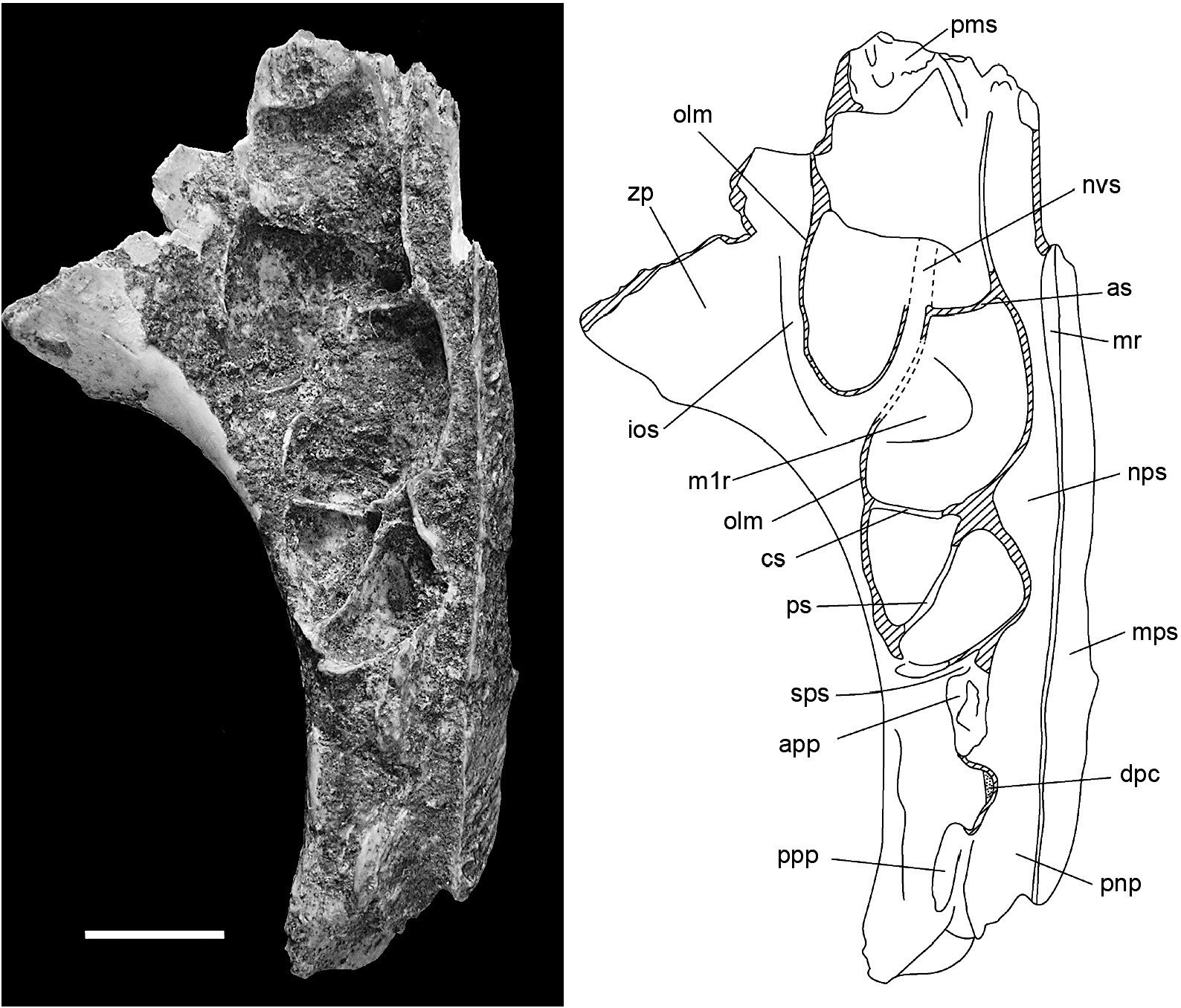
Though only the root of the zygomatic plate is preserved on AMF 68770 ( fig. 12B, C View Fig ), enough survives to show that it was relatively small and lightly built, especially considering the moderate wear of the molars in this individual. It measures 9.5 mm in length and its posterior margin is level with the anterior face of M1. The anterior root is closely occluded by the rostral lamina of the maxilla, creating a slitlike anteroventral aperture for the infraorbital fissure. As in C. buehleri , the posterior root of the zygomatic plate is thinned to accommodate the lateral expansion of the posterior rostral skeleton, reflecting enlargement of the maxillary sinus complex ( figs. 13 View Fig , 15 View Fig ).
A shallow depression situated just behind and ventral to the anterior root of the zygomatic arch on AMF 68770 is identified as the superficial masseteric tubercle ( fig. 12C View Fig ). The posterior margin of the tubercle lies 6.5 mm forward of the anterior face of M1, more than twice the distance in C. buehleri . A sharply defined masseteric ridge, aligned with the lingual cusp series of M1, originates 3.3 mm forward of M1 and terminates medial to the tubercle. A nutrient foramen cannot be identified in this region of the maxilla on AMF 68770, but a small foramen might be obscured by encrusted sediment. A second maxillary fragment AMF 68756 has a nutrient foramen located just forward of M1 and lateral to the masseteric ridge.
The alveolar body of the maxilla is deeper than in C. buehleri , and the lateral surface maintains a near vertical orientation above the entire molar row. The position of the posterior palatal foramen is obscured by cemented sediment on AMF 68770. On ANWCP26 ( fig. 12D View Fig ), a posterior palatal fragment with conjoined portions of both maxillae and palatines, the posterior palatal foramen is level with cusp t7 of M2. This specimen also shows that the palatal surface of the palatine bone is perforated by numerous vascular canals, suggestive of a highly vascularized anterior portion of the soft palate.
The well-preserved dorsal surface of AMF 68770 ( figs. 13 View Fig and 15 View Fig ) illustrates the full lateral development of the maxillary sinus complex, including its extension onto the dorsal surface of the root of the zygomatic arch. It also shows the entire course of the neurovascular sulcus for the nasal branch of the infraorbital artery (and associated nerve). The following minor differences are seen in comparison with C. buehleri : the neurovascular sulcus in C. musseri appears to traverse the maxillary sinus complex somewhat more medially and posteriorly, suggesting an origin farther back within the intraorbital sulcus; and the medial rim of the anterior nasopharyngeal sulcus in C. musseri is lower anteriorly, and less extensive posteriorly.
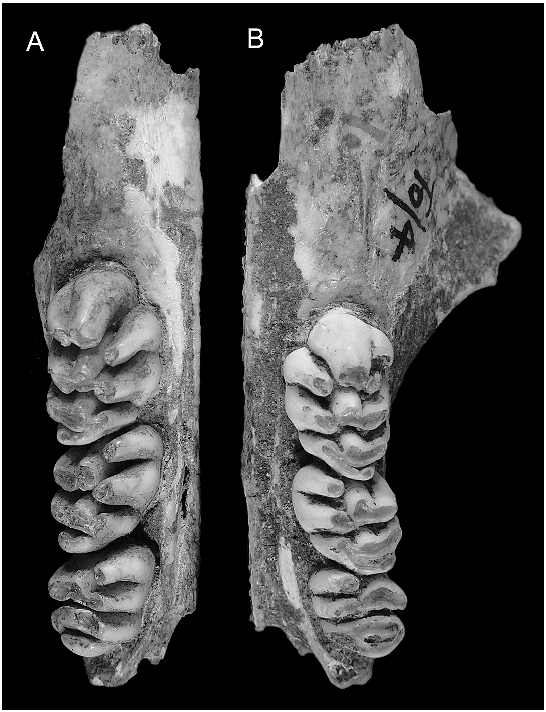
DENTARY ( fig. 16 View Fig ): All 15 dentaries attributed to this taxon are damaged and no specimen preserves either the coronoid or angular processes or the articular condyle. The holotype (ANWCP32, fig. 16A–B View Fig ) and three other specimens at contrasting wear stages illustrate the changes in mandibular form that occur through life ( fig. 16 View Fig ).
The dentary is similar in size and general morphology to that of C. buehleri . Comparison of specimens of equal wear stages reveals the following points of contrast: (1) the molar row is smaller in proportion to the dentary; (2) the inferior masseteric crest is more prominent; (3) the digastric process is more prominent and has a more pronounced posterior rim; (4) the fossa for the deep portion of the transverse mandibular muscle is substantially larger; and (5) the postalveolar ridge runs dorsal to the superior mandibular foramen and extends further onto the neck of the condylar process.
In combination, these features reflect a more powerfully developed masticatory musculature in C. musseri , compared with the larger-toothed C. buehleri . Enlargement of the fossa for the deep portion of the transverse mandibular muscle is particularly noteworthy. In laboratory rats this muscle is particularly active during adduction of the mandible ( Weijs and Dantuma, 1975), with muscular contraction causing spreading of the anterior and superior borders of the symphysis and relieving compression on the symphyseal cartilage ( Beecher, 1979).
One specimen (ANWCP12, not shown) with m1–3 retained has a clearly defined alveolus positioned behind the m3. This is as broad as the posterior lamina of m3 and presumably held a peglike tooth on a substantial root. Supernumerary fourth molars are rare in rodents generally, and seem to be especially so in Murinae (summarized by Johnson, 1952).
One edentulous dentary (AMF 68831, not shown) is referred to Coryphomys on the basis of the well-developed retromolar fossa, and tentatively assigned to C. musseri on the basis of inferred molar dimensions. The deep ramal body and strong development of the alveoli for m3 suggest an aged individual. The m 1 in this specimen bore a small accessory root, midway along the lingual side of the tooth, in addition to a large anterior root with its posterolabial extension and a broad posterior root. Each of the m2 and m3 was supported by transversely widened anterior and posterior roots. The lingual margin of each alveolus is vertical, but the labial margins are splayed distally to provide enhanced anchorage. The anterior alveolus of each of m2 and m3 is bilobed in its deepest expression. Partially separate labial and lingual anterior roots are also observed in the shallower alveoli of a young individual of C. musseri (ANWCP27, fig. 16C–D View Fig ) that has lost m2–3.
| R |
Departamento de Geologia, Universidad de Chile |
| MK |
National Museum of Kenya |
| CM |
Chongqing Museum |
| RM |
McGill University, Redpath Museum |
| JM |
Jura Museum, Eichstatt |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.