
Coryphomys buehleri Schaub, 1937
|
publication ID |
https://doi.org/10.1206/692.1 |
|
persistent identifier |
https://treatment.plazi.org/id/FA68571D-313E-FF84-FF14-33BC92958642 |
|
treatment provided by |
Tatiana |
|
scientific name |
Coryphomys buehleri Schaub, 1937 |
| status |
|
Coryphomys buehleri Schaub, 1937 View in CoL
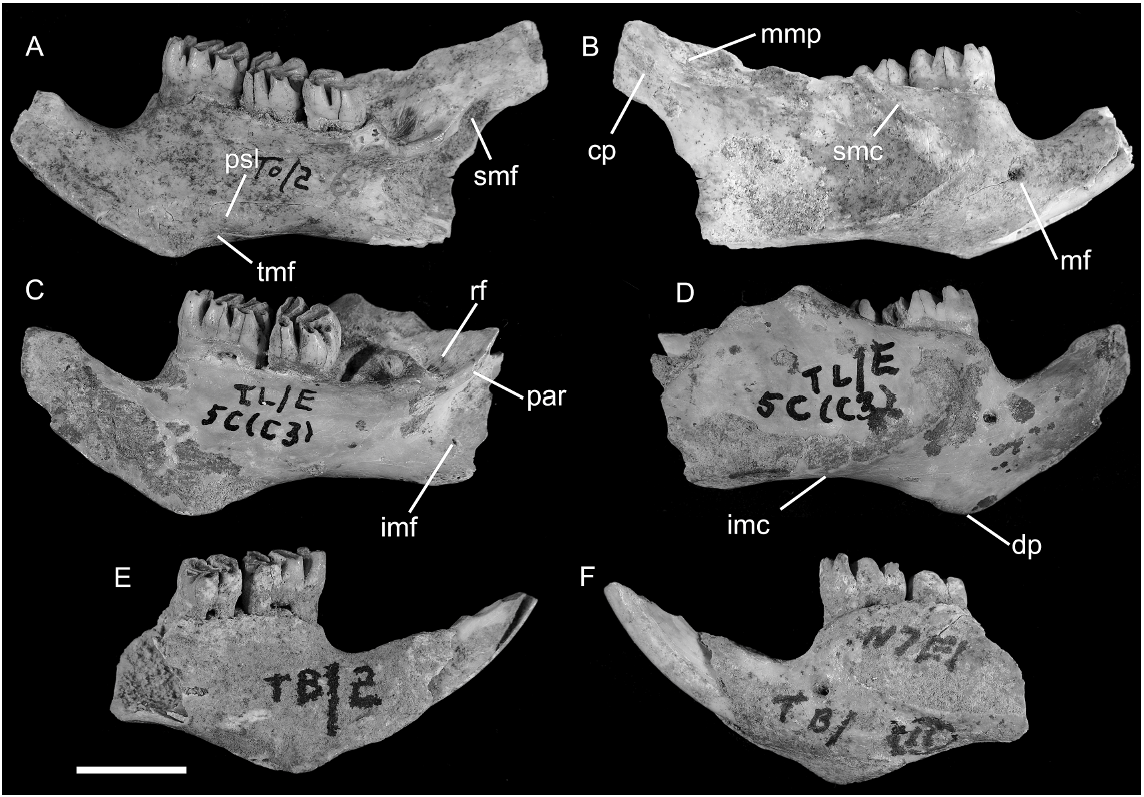
HOLOTYPE: Naturhistorisches Museum Basel, NMB A.P.1. A right dentary with m1–3 and root of incisor; preserving condylar process but with damage to coronoid and angular processes. Figured by Schaub (1937: figs.1 View Fig and 2 View Fig ). Not examined for this study.
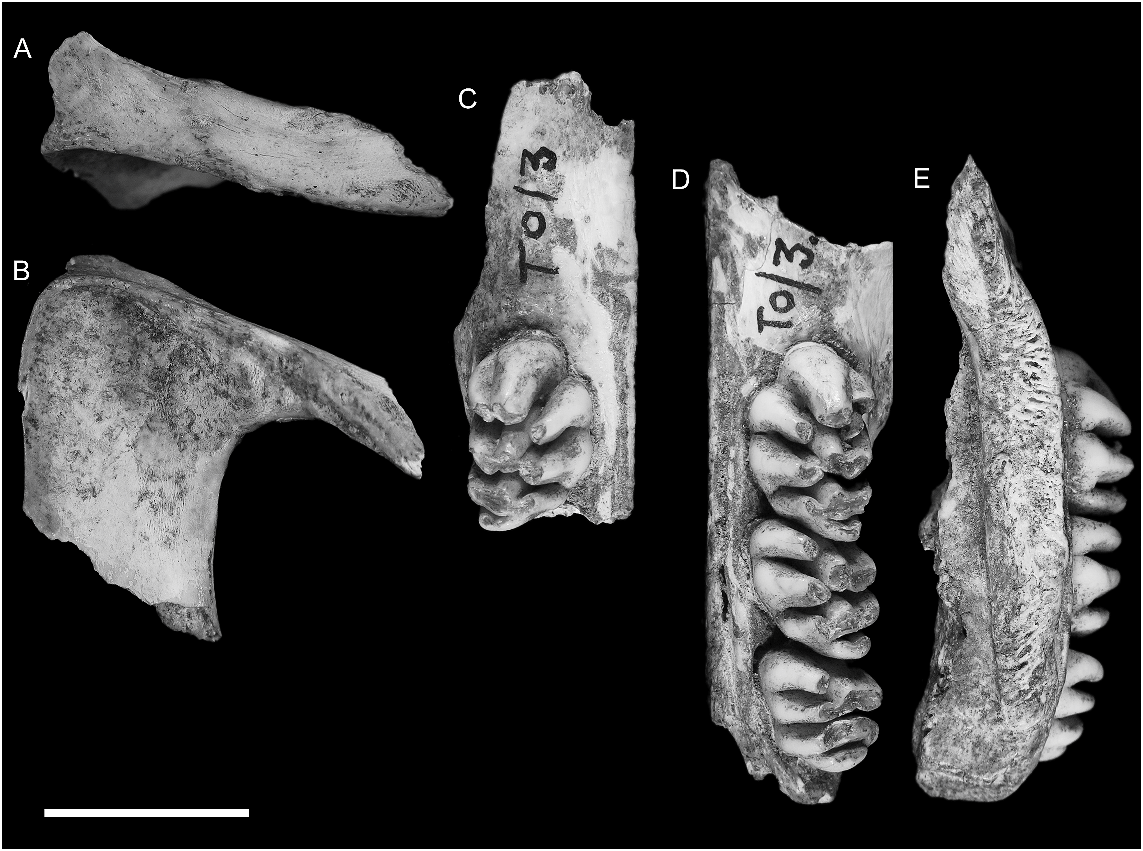
REFERRED MATERIAL: AMF 68831: Uai Bobo 1 (TO1/2), R Dentary with m1–3; AMF 68789: Bui Ceri Uato (TB/2), L Dentary with m1–2; AMF 68850: Lie Siri (TL/E 5C/C 3), R Dentary with m1–2; ANWCP 1: Jerimalai ( JM /B/19) L m2; ANWCP 2: Matja Kuru 1 ( MK 1/A/17), L M3; ANWCP 3: Matja Kuru 1 ( MK 1/AA/ 28), R M2; ANWCP 4: Matja Kuru 1 ( MK 1/ AA/25), L M3; AMF 68838: Lie Siri (TL/E/5C/C 3), L maxilla with M1; AMF 68751a: Uai Bobo 1 (TO1/3), L palatal fragment (partial maxilla and palatine) with M1–3; AMF 68751b: Uai Bobo 1 (TO1/3), R maxilla fragment with M1; AMF 68751c: Uai Bobo 1 (TO1/3), L zygomatic plate; ANWCP 5: Matja Kuru 1 ( MK 1/AA/10), R M1; ANWCP 6: Matja Kuru 2 ( MK 2/D/32), R m1; ANWCP 7: Matja Kuru 2 ( MK 2/D/ 41), L M3; AMF 68824: Uai Bobo 1 (TO1/3), R M1; ANWCP 8: Matja Kuru 1 ( MK 1/AA/ 10), L M2.
REMARKS: Given the presence on Timor of various other equally large murines, it is possible that the murine femur referred by Schaub (1937: fig. 3 View Fig ) to Coryphomys buehleri may not belong to this taxon. Its identity will be reassessed following allocation of the abundant postcranial remains now available from the many excavated assemblages.
Only one of the three dentaries (no. 2) attributed to C. buehleri by Hooijer (1965) actually belongs to this species. Hooijer’s specimen no. 1 is referable to Mahoney’s Genus A, while his no. 3 belongs to the new species of Coryphomys described below. Ironically, Coryphomys buehleri , the only formally described member of the prehistoric Timorese murine fauna, is also one of the rarest of all the large-bodied taxa in the fossil samples.
Attribution of a maxillary dentition to this species is based on the recognition of a morphologically distinctive subset of Coryphomys specimens that are also slightly larger in all dental dimensions relative to those of the second species.
EXTENDED DESCRIPTION OF CORYPHOMYS BUEHLERI
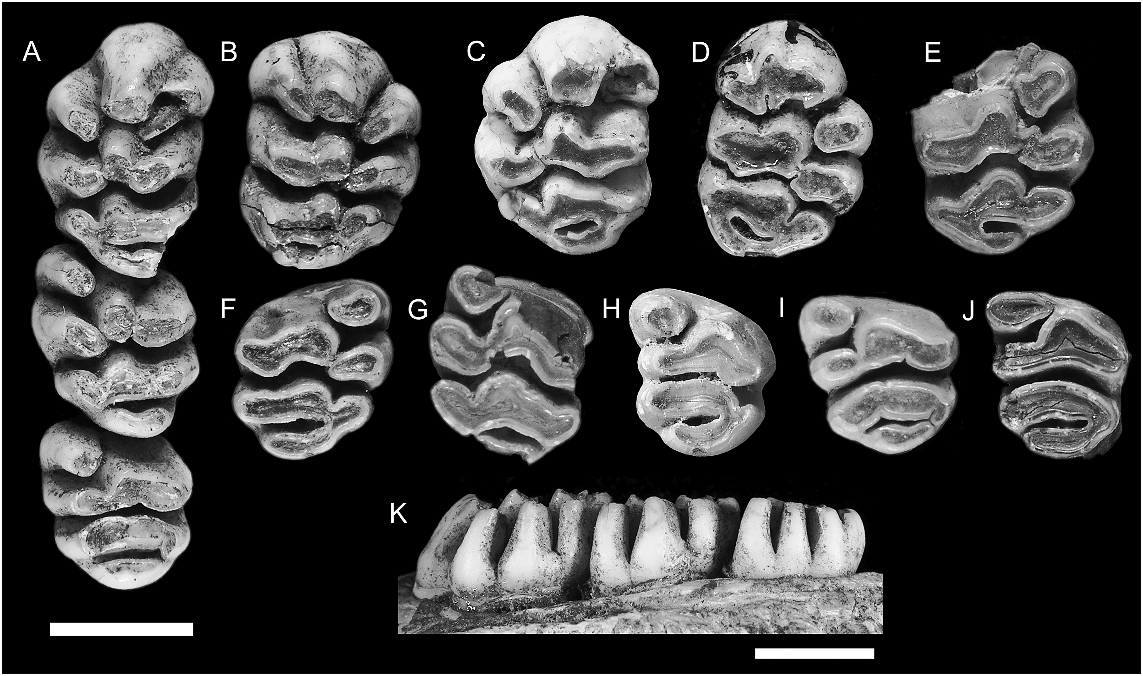
UPPER MOLARS ( fig. 4 View Fig ; table 1): The upper molars are moderately hypsodont but comprised of variably discrete and partially united cusps arranged in more or less transverse series. The occlusal plane of the molar row is tilted outward (i.e., lingual cusps are significantly lower than labial cusps) but there is little indication of helical torsion along the molar row.
No maxillae without molars are confidently assigned to this species. The M1 has four roots. The anterior root, supporting the anterior lamina, is broad and kidney shaped with the concavity facing the rear. Two small circular roots on the lingual side of the tooth support cusps t4 and t7. A posterolabial root, intermediate in size, supports cusps t8 and t9 and the associated posterior cingulum. The root pattern for M2 cannot be determined from available specimens. Two isolated M3 show a broad anterior root supporting cusps t1 and t4–6, a somewhat narrower root supporting cusps t7–8 and a small labial rootlet above cusp t9 and partially united to the front of the posterior root.
M1: This tooth is represented by a total of five specimens ( fig. 4A–E View Fig ). They are broadly ovate in occlusal outline but have a weakly concave labial margin.
The anterior lamina consists of three discrete, columnar cusps. The central cusp t2 curves forward and downward from a bulbous base, narrowing toward an occlusal surface that is rounded anteriorly but flattened on each of the labial, lingual, and posterior margins. The labial cusp t3 is positioned level with cusp t2. It is firmly adpressed to the labial surface of cusp t2 but is fully encircled by enamel for approximately one third of its total height. Above that level, the two cusps are united but with welldefined anterior and posterior grooves that extend to the crown base. Cusp t3 also has a bulbous base and narrower apical portion. The occlusal surface is ovate, with the long axis oriented at approximately 45 ° to the coronal major axis. Cusp t1 is isolated from the cusp t2–3 complex by a deep cleft that
TABLE 1
Measurements of Upper and Lower Dentition in Specimens Attributed to Coryphomys buehleri
Measurements are molar lengths (L) and widths (W) and the breadth (IB) and depth (ID) of the lower incisor. Sample sizes, means, standard deviations, and ranges are also shown.
extends more than three quarters of the way to the crown base. The cusp is bulbous basally but narrows apically and folds against the anterolingual surface of cusp t5. A weak posterolingual groove on cusp t1 produces a kidney-shaped occlusal surface which is oriented at approximately 45 ° to the main coronal axis. The occlusal surface of cusp t1 lies in a plane slightly above that of cusps t1–2 and t5.
The middle lamina consists of a united but lobular cusp t5–6 complex and a discrete but adpressed cusp t4. Cusps t5 and t6 are approximately equal in bulk but differ in shape. The central cusp t5 is columnar and laterally compressed, and is smaller in occlusal area than cusp t2. Cusp t5 is firmly connected to cusp t6, but the cusp boundary is marked by strong posterior groove. Cusp t6 is an elongate, transversely oriented structure with a bulbous base, a rounded posterior surface, a flattened anterior surface and a well-developed posterolingually directed ridge. Cusp t4 is similar in basic shape and orientation to cusp t1, but differs in the presence of a well-developed anterolabial ridge that abuts against the corresponding ridge from cusp t4. As a result, the occlusal surface of cusp t4 is broadly arcuate (concave anteriorly) and continuous with the cusp t5–6 complex. The occlusal surface of cusp t4 lies in the same plane as cusp t1 and is thus slightly above that of cusps t5–6.
The posterior lamina consists of a transversely oriented t8–9 complex and a prominent t7 that is firmly connected to cusp t8. The central cusp t8 is similar in size and shape to cusp t5. The boundary between cusps t8 and t9 is well marked in relatively unworn specimens by an apical cleft but this is rapidly obliterated by wear. The lateral margin of cusp t9 supports a broad anterolabially directed ridge that ascends to the crown base. With progressive wear, this ridge produces an elongation of the occlusal surface of cusp t9. Cusp t7 is discrete apically from t8 but soon unites to this cusp with wear. In contrast, the fissure separating cusp t7 from cusp t4 extends almost to the crown base, thereby keeping the two cusps separate, even under advanced wear.
The posterior cingulum is an elevated transverse ridge that forms the posterior surface of the tooth. The cingulum shows a variable pattern of connections. On both sides of AMF 68751 ( fig. 4A–B View Fig ) it is firmly connected labially to the point of union of cusps t8 and t9, but remains well separated from the posterior surface of cusp t7 as it rises toward the crown base, creating a lingually open cingular shelf. On AMF 68838 ( fig. 4C View Fig ) the labial end of the cingulum is united high to the anterior surface of cusp t9, thereby enclosing a distinctive cingular fossette. AMF 68824 ( fig. 4D View Fig ), with more advanced wear, also has a fully enclosed fossette with broad cingular connection at both labial and lingual ends. Two small pimples on the elevated portion of the ridge create a weakly bicuspid occlusal outline on AMF 68751 ( fig. 4A–B View Fig ).
M2: Three specimens are available ( fig. 4A, F–G View Fig ), each at a different wear stage. The less worn example, part of the complete molar series in AMF 68751 ( fig. 4A View Fig ), is described first. In both size and cuspal arrangement the M2 closely resembles M1, save for the absence of elevated cusps t2–3. Cusp t1 is more vertically oriented than on M1 but is similarly folded against the anterolingual surface of cusp t5. Its posterior surface is rounded and the occlusal surface is correspondingly ovate rather than kidney shaped. The occlusal surface of cusp t1 lies in a plane slightly above that of cusp t5. Cusp t3 is represented by a small nubbin positioned near the crown base in the fold that marks union of cusps t5 and t6.
Cusps t5 and t6 of the middle lamina are firmly united but cuspal limits are marked by strong anterior and posterior grooves. The central cusp t5 is columnar and laterally compressed, and is slightly larger than the serial homolog on M1. Cusp t6 is larger in occlusal area than cusp t5. It is broader and more transversely oriented than the serial homolog on M1 but otherwise similar in form. Cusp t4 is similar in basic shape and orientation to cusp t1 but is larger in occlusal area. It lacks the anterolabial ridge seen on cusp t4 of M1 and this has a simpler, ovate occlusal outline. The occlusal surface of cusp t4 lies slightly above that of cusps t5–6, but the contrast is less marked than on M1.
The posterior lamina is an almost exact duplicate of this structure on M1 except that cusp t7 is slightly larger in occlusal area. The posterior cingulum is connected at its lingual end to the posterior surface of cusp t7. The labial end is deeply separated from cusp t9 by a cleft that ascends almost to the crown base.
ANWCP3 ( fig. 4F View Fig ) is in a more advanced state of wear. It shows comparable features save for a more elevated connection between the labial end of the posterior cingulum and the posterior surface of cusp t9. Cusps t7–t9 and the posterior cingulum are united by a common dentin pool in this specimen, while cusps t4 and t1 each remains discrete from cusp t5. ANWCP8 ( fig. 4G View Fig ) is even more heavily worn. Cusps on both laminae are united into common dentin pools but cusp t9 remains isolated from the labial end of the posterior cingulum.
M3: Four specimens are available ( fig. 4A, H–J View Fig ). The least worn examples of this tooth (AMF 68751, fig. 4A View Fig ; ANWCP4, fig. 4I View Fig ) are only slightly shorter and narrower than M2, but they differ in numerous morphological features. Cusp t1 is more conical in form but bears a short ridge on its labial surface, directed to the front of cusp t5. Cusps t5 and t6 are firmly united into a transversely oriented lamina with a common dentin pool. There is no delimiting anterior groove, and no cusp nubbin or fossette in the position of cusp t3. However, a posterior groove indicates the relative contribution of the two cusps, with cusp t6 being slightly the larger. Cusp t4 is smaller and more transversely oriented than it serial homologs on M1–2; the occlusal plane is only slightly above that of cusp t5 and the two cusps are united by a common dentin pool.
Cusps t7 and t8 are united into a broad, transversely oriented structure that is gently convex anteriorly. The boundary between the two equal-sized cusps is marked by a slight indentation of the anterior margin and a corresponding flexion of the occlusal surface. Cusp t9 is a smaller columnar cusp that is firmly applied to the labial surface of cusp t8 but projects posterior to this cusp.
The posterior cingulum forms a robust, transversely oriented lamina. It is united high on the crown to the posterior surfaces of cusps t7 and t9 and has a rounded and bulbous posterior surface. The occlusal surface consists of two facets, the lingual facet paralleling that on cusp t7.
The most heavily worn example of M3 (ANWCP2, fig. 4J View Fig ) has the posterior cingulum joined by a common dentin pool to t9 but still discrete from cusp t7 at the lingual end.
LOWER INCISOR: The incisor is retained in AMF 68789 ( fig. 9F View Fig ) and lacks only the tip. The body of the tooth is approximately Dshaped in cross-section and has a width-todepth ratio of 0.645 (table 1). A smoothly curved strip of enamel is present along the ventral and lower lateral surfaces of the tooth, extending just over half way up the lateral surface of the tooth and terminating at the widest point of the tooth. Enamel is absent from the medial surface, save for a 0.34 mm rim corresponding to the thickness of the enamel layer. Pale orange pigment is present medially in a 1.6 mm wide band. Otherwise the enamel is unpigmented. The enamel-free dorsal half of the incisor bears a weak dorsolateral groove. The medial surface is very slightly concave. The occlusal facet is incomplete but terminates posteriorly in a distinct step.
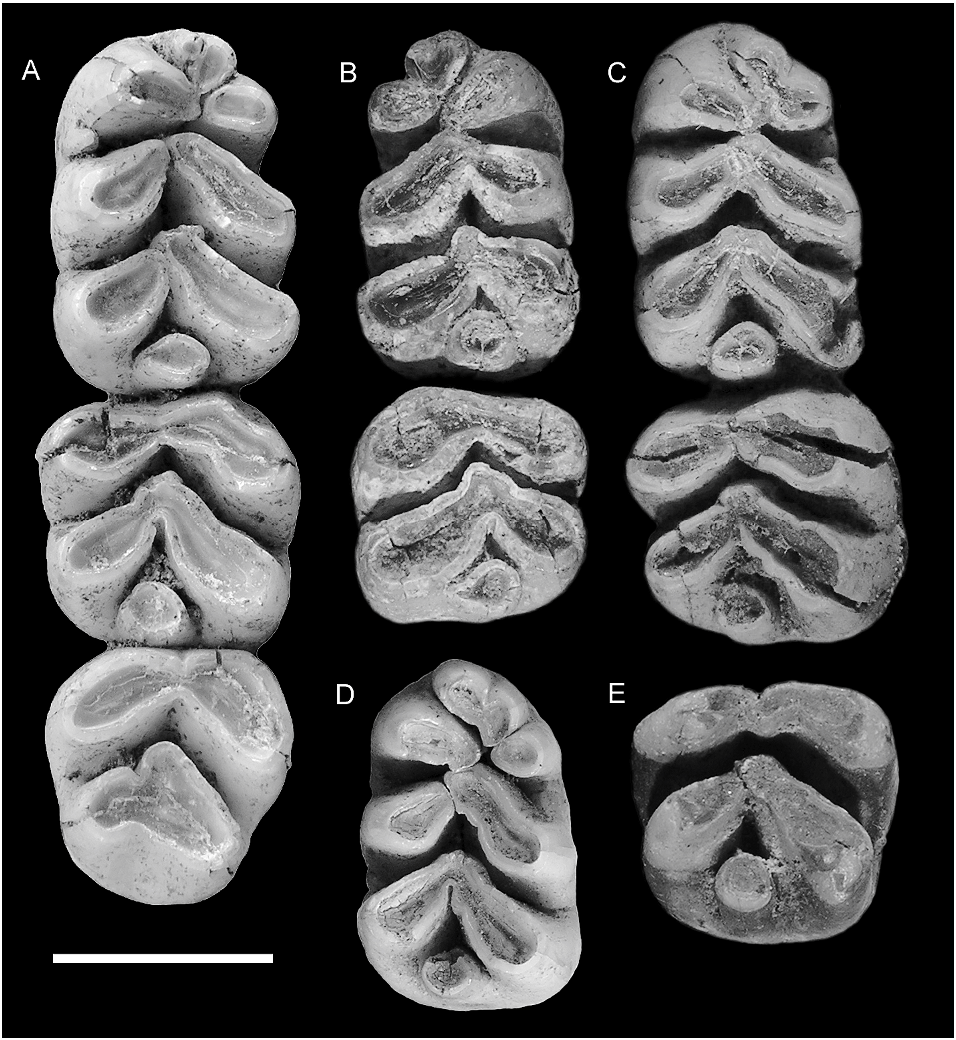
LOWER MOLARS ( fig. 5 View Fig , table 1): The lower molars are also moderately hypsodont. The enamel is smooth on all surfaces, irrespective of intensity of wear. The occlusal plane of the molar row has a moderate degree of helical torsion, changing from transversely horizontal on m1 to lingually inclined (i.e., lingual cusps slightly lower than labial cusps) on m3.
No dentaries without molars are confidently assigned to this species; hence, information on molar root patterns is based on careful examination of teeth in situ. The m1 has two roots. The posterior root has similar dimensions on the labial and lingual sides and appears to bear a posterior groove; it probably occupies a symmetrical and possibly weakly bilobed alveolus. In contrast, the anterior root appears much broader labially than lingually, and the alveolus is presumably comma shaped with an anterior ‘‘head’’ and a posterolabial ‘‘tail.’’ The m2 appears to have two broadly united, transverse roots, one supporting each of the two principal chevrons. The posterior root, as exposed on AMF 68789, is solidly united to the root apices and shows an increase in width with depth. This firm anchoring of the molars presumably accounts for the high rate of retention of molars in preserved dentaries of this taxon. The alveoli of m3 are exposed on AMF 68850. The anterior alveolus is broad and contained a fused root that terminated in short separate nubbins. The posterior alveolus is narrower and more circular.
m1: This tooth is represented by four new examples ( fig. 5A–D View Fig ). They have a rectangular outline that is broadest across the posterior lamina and narrows slightly to the front. The anteroconid is relatively large and equals the two posterior lophids in occlusal area. The principal cuspids are weakly united, even in heavily worn examples. The occlusal surfaces of the labial and lingual rows of cuspids are elevated externally and slope down to a central occlusal valley. The external surfaces of the labial cuspids are vertical, whereas those of the lingual cuspids are slightly bowed in less heavily worn examples.
The anteroconid shows three principal cuspids arranged in a somewhat variable pattern. The anterolingual cuspid is consistently the largest and most discrete of the three. It has an ovate occlusal outline, with the labial end adpressed against the front of the protoconid and a long axis that swings approximately 30 ° forward of transverse. The anterolabial cuspid is transversely aligned with the posterolabial end of the anterolingual cuspid but it is considerably smaller and has a more rounded occlusal outline. In all specimens the two cuspids are deeply separated almost to their bases. The anterocentral cuspid is more variable in size, morphology, and relations. In AMF 68850 ( fig. 5C View Fig ) it is a simple rounded cuspid that is broadly connected to the anterior surface of the anterolabial cuspid. All other examples show a bicuspid anterocentral cuspid. In ANWCP6 ( fig. 5E View Fig ) it is divided into two subequal components by an anterolabial groove; the posterolabial end is adpressed against the anterolabial cuspid. AMF 68831 ( fig. 5a View Fig ) shows a larger labial component and a smaller, pimplelike lingual component attached high on the anterolingual side; the combined anterocentral cuspid in this example is equally discrete from each of the anterolabial and anterolingual cuspids. AMF 68789 ( fig. 5B View Fig ) has the anterocentral cuspid divided into a larger lingual component and a smaller labial component, the combined cuspids having a specific attachment to the anterolabial cuspid. In all cases, the occlusal surface of the anterocentral cusp is elevated anteriorly and slopes down to the rear, thereby blocking the longitudinal occlusal valley of m1. The type specimen, as described and illustrated by Schaub (1937: fig. 2 View Fig ), has a divided anterocentral cuspid. Hooijer’s (1965: pl. I) specimen no. 2, herein referred to C. buehleri , possesses an undivided cuspid.
Two specimens show small accessory cuspules attached to the anteroconid. AMF 68831 ( fig. 5A View Fig ) has a small cuspule attached to the posterolingual base of the anterolingual cuspid. The other ( ANWCP2 , fig. 5C View Fig ) has a basal cuspule attached to the posterior surface of the anterolabial cuspid ; a weak buttresslike ridge from the base of the protoconid joins this structure to produce a partial cingulum across the base of the anterolabial flexid. These cuspules appear to be absent from the holotype and Hooijer’s specimen no. 2, judging from the respective illustration and figure.
The anterior lamina of m1 is made up of two elongate cuspids arranged in a forwardfacing chevron. The labial protoconid is elongate and has an occlusal surface that is variably rectilinear or weakly dumbbell shaped, the latter condition produced by shallow concavities on both the anterolabial and the posterolingual surfaces. The lingual metaconid is D-shaped, flattened anteriorly, curved posteriorly, and smaller in occlusal area than the protoconid. In two specimens ( AMF 68831, fig. 5A View Fig ; AMF 68850, fig. 5C View Fig ) the metaconid abuts the anterior end of the protoconid without overlap. In the other two ( AMF 68789, fig. 5B View Fig ; ANWCP6 , fig. 5D View Fig ), the protoconid overlaps the front of the metaconid, so that the latter abuts the lingual face of the protoconid. In both conditions the two cuspids remain narrowly separate almost to the crown base. Hooijer’s (1965: pl. I) specimen no. 2, herein referred to C. buehleri , has the two cuspids abutting without overlap. The anterolophid chevron has an anterior angle of approximately 130 ° and a posterior angle of approximately 70 °.
The posterior lamina of m1 repeats the basic structure of the anterior lamina but with minor differences in cusp size and orientation. The labial hypoconid is virtually identical in size and orientation to the protoconid, even to the presence of a low anterolabial buttress that partially encloses the posterolabial flexid. The lingual entoconid is slightly larger in occlusal area than the metaconid. Centrally, the hypoconid and entoconid are more firmly adpressed and the two cuspids show a transversely continuous dentin pool in more heavily worn examples. The hypoconid and entoconid abut without overlap on one specimen (ANWCP6, fig. 5D View Fig ), but in the other three the hypoconid overlaps the front of the entoconid. They abut in Hooijer’s (1965: pl. I) specimen no. 2, herein referred to C. buehleri . The posterior chevron has an anterior angle of approximately 115 ° and a posterior angle of approximately 90 °. There is no contact between the anterior and posterior laminae above the level of the crown base.
The posterior cingulum is an isolated, round to oval-shaped cusp. It is positioned low and centrally at the rear of the tooth and, unlike the primary cusps, stands vertical rather than sloping forward.
m2: This tooth is represented by four new specimens ( fig. 5A–C, E View Fig ) one of which (ANWCP1, fig. 5E View Fig ) is essentially unworn. All examples are slightly wider than long and the anterior lamina is slightly wider than the posterior lamina.
The anterior lamina is essentially transverse in orientation, with a broad and shallow anterior concavity and deeper and more broadly U-shaped posterior groove. In lightly worn specimens (ANWCP1, fig. 5E View Fig ; AMF 68831, fig. 5A View Fig ) the protoconid and metaconid contributions are defined by a short, vertical fissure located approximately two-thirds across from the labial margin. With additional wear the protoconid and metaconid rapidly merge into a common dentin pool. The metaconid is teardrop shaped in occlusal outline. The protoconid is more variable in shape. On three specimens it has a distinct anterolabial groove that may indicate the position of an incorporated anterolabial cuspid. One specimen lacks the anterolabial groove, as does Hooijer’s (1965: pl. I) specimen no. 2, herein referred to C. buehleri . The posterior surface of the protoconid is gently rounded in two specimens and in Hooijer’s (1965: pl. I) specimen no. 2, is narrowly grooved in one, and more deeply grooved in another.
The posterior lamina is a chevron-shaped structure that closely resembles the serial homolog on m1. The hypoconid and entoconid are more intimately united and share a common dentin pool after minimal wear. The posterior cingulum is erect and rounded on all specimens. In AMF 68789 ( fig. 5B View Fig ) it has a more elevated connection to the hypoconid than to the entoconid. The anterior and posterior laminae are unconnected above the level of the crown base.
m3: The m3 is represented by one new specimen (part of AMF 68831, fig. 5A View Fig ) and is also present in Hooijer’s (1965: pl. I) specimen no. 2, herein referred to C. buehleri . The m3 is subequal to the m 2 in length and width as measured across the anterior lamina and is only slightly smaller than the m 2 in occlusal area. The occlusal surface is weakly angled on the anterolophid but transversely flat on the posterior lamina.
The anterior lamina is a broadly open chevron, with a slightly concave anterior face and a posterior angle of approximately 95 °. The protoconid is slightly larger than the metaconid in occlusal area. There is no anterolabial cuspid on AMF 68831, but the anterior margin of the protoconid is angular, suggestive of an amalgamated cuspid; this part of the tooth is more smoothly rounded in Hooijer’s specimen no. 2.
The posterior lamina is considerably narrower than the anterior lamina on the m3 of AMF 68831, compared with slightly narrower on the holotype, as described and illustrated by Schaub (1937: fig. 2 View Fig ). It consists of a larger, obliquely oriented hypoconid and a smaller, adpressed conical entoconid that is defined by clear anterior and posterolingual grooves. On Hooijer’s specimen no. 2 the two laminae are almost equal in width. A posterolingual groove alone marks the division between the hypoconid and entoconid on this more heavily worn tooth.
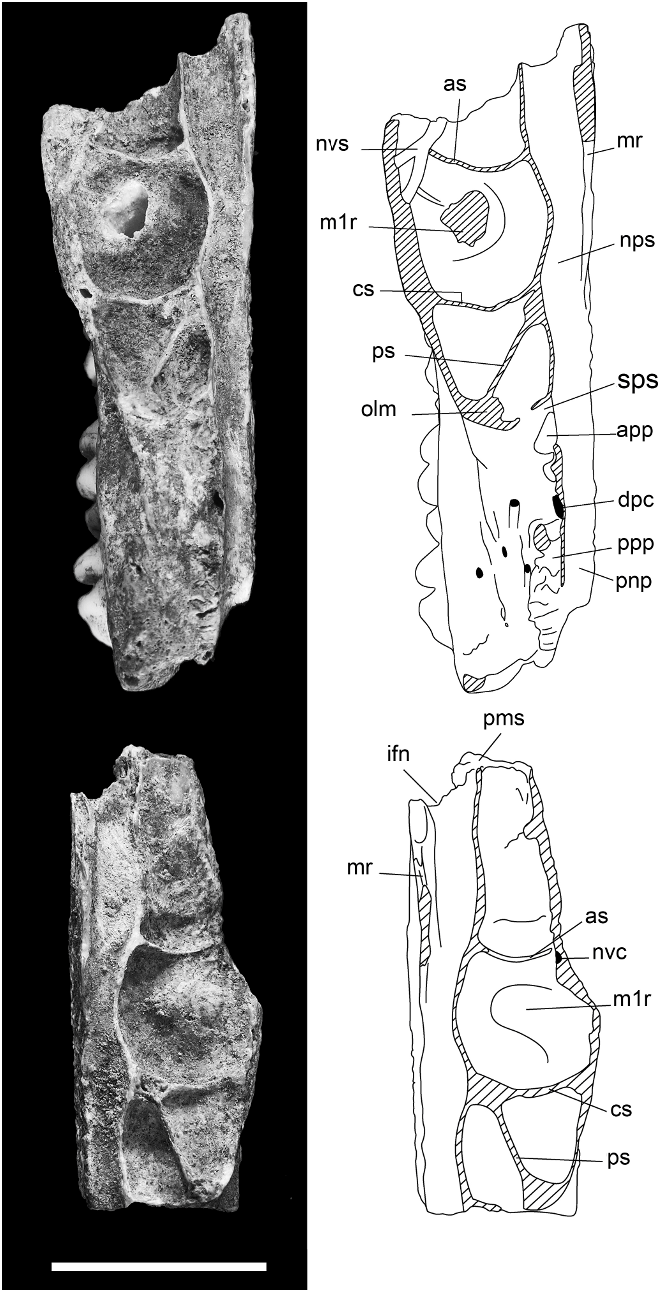
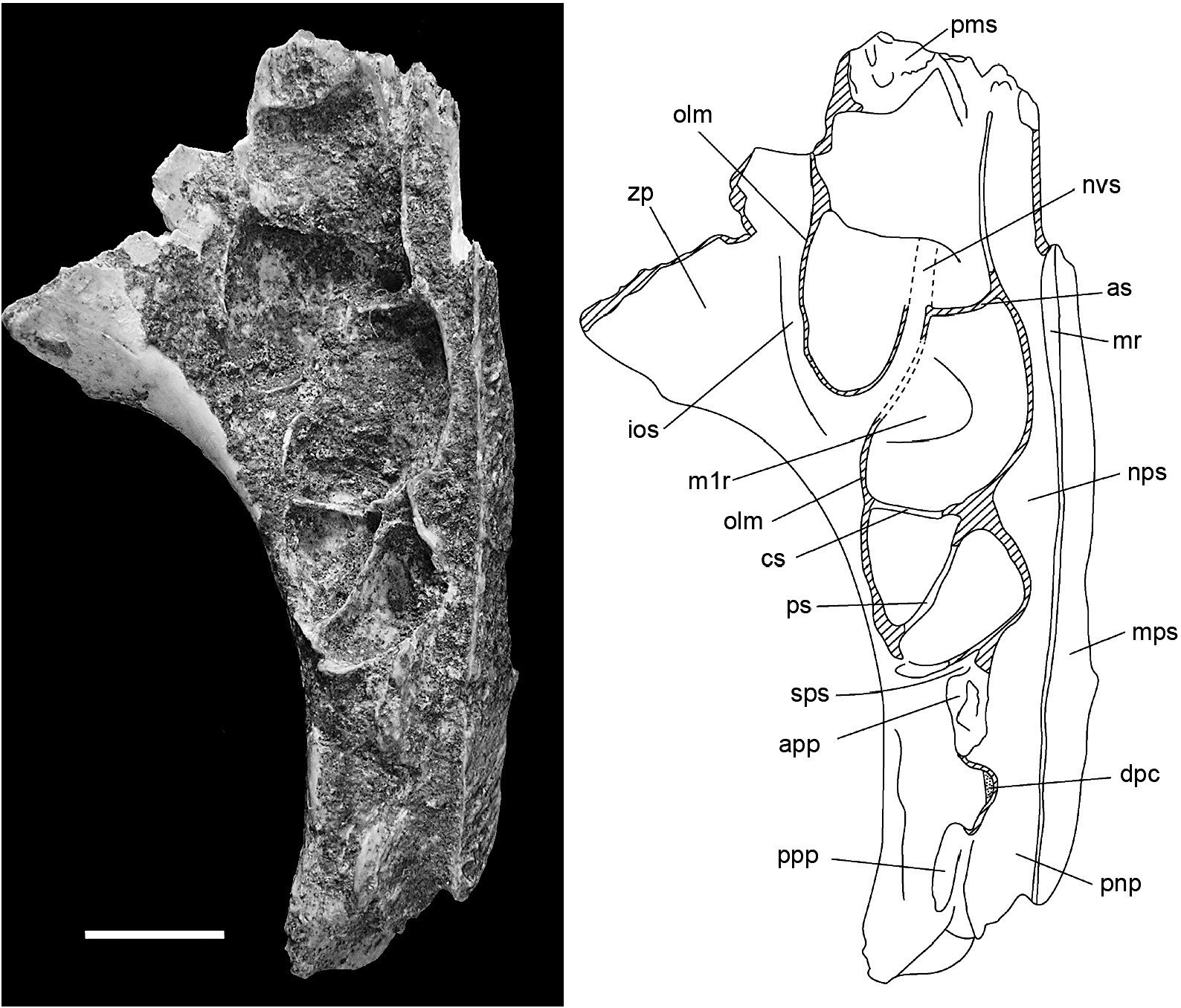
MAXILLA ( figs. 6–8 View Fig View Fig View Fig , 15 View Fig ): Information on the structure of the maxilla and palatine bones comes from associated left and right palatal fragments ( AMF 68751, fig. 6 View Fig ). The left palatal fragment ( AMF 68751a, fig. 6D View Fig ) retains the full length of the maxilla, from the medial suture with the premaxilla back to the posterior end of the alveolar portion, and preserves a complete midline palatal suture that includes a fragment of the palatine. Only the roots of the zygomatic and orbital processes are preserved. The right palatal fragment ( AMF 68751b, fig. 6C View Fig ) is less complete posteriorly but retains a larger portion of the antemolar palatal lamina. The specimen is partially mirrored in figure 7 View Fig to produce a composite that illustrates the major features of the palate. A left zygomatic plate from the same excavation unit probably derives from the same individual ( AMF 68751c, fig. 6A–B View Fig ). Though no direct contact exists between this specimen and the left maxilla, the two specimens are complementary and show an identical state of preservation and discoloration. Two other reasons for accepting this association are: (1) the basal portion of the plate is consistent with this region in the second species of Coryphomys , to be described below ; and (2) the morphology of the zygomatic plate differs from that of confidently associated specimens of each of the three other genera.
The most prominent features of the palate are the exceptionally large size of the molar rows relative to the bony support structures, the narrowness of the palatal bridge that separates them, the midpalatal antemolar depression, the short posterior palatal bridge, and the short and extremely narrow incisive foramina.
The palatal lamina of the maxilla is extremely thick, measuring 3.8–4.0 mm in depth at the medial suture and only thinning rapidly toward the anterior suture with the premaxilla ( fig. 6E View Fig ). The sutural contact between the palatine and maxilla, visible on the medial palatal suture, is situated level with the center of M2. The rear of the bony palate is level with cusp t4 of M3. The anterior sutural contact with the premaxilla is located 11.1 mm forward of the anterior face of M1. The total length of the maxilla is reconstructed as 33.3 mm, compared with the M1–3 alveolar length of 19.8 mm.
A deep palatal groove extends from the rear of the bony palate through to just forward of M1. Situated within this groove is the posterior palatal foramen, located approximately level with the front of M2. A sulcus located behind M3 marks the position of a posterior palatal pit and associated foramen.
The antemolar palatal lamina of each maxilla is arched dorsally, creating a broad midpalatal antemolar fossa. This surface lacks vascular grooving. The anterior margin of the palatal lamina bears a 2.7 mm deep notch for the posterior border of the incisive foramen. These paired structures were evidently very short, terminating some 9.8 mm forward of the molar rows, and narrow, with a combined width of only 3 mm. The lateral margin of the antemolar fossa is marked by a short ( 3 mm) but sharply defined masseteric ridge, marking the ventral limit of insertion of the anterior portion of the deep part of the masseter. This ridge originates at the centre of the M1 alveolus and runs forward to the medial side of a rugose fossa situated 2.5 mm forward of the alveolus and aligned with the labial row of cusps of M1. This is identified as the superficial masseteric fossa (for attachment of the inferior zygomatic plate aponeurosis); it is positioned remarkably close to the M1, but we note a similar condition in Mallomys . No nutrient foramen is visible anterior to M1.
The alveolar body of the maxilla is unusually shallow, especially given the great size of the molars. The lateral surface is near vertical above M1, but it twists to face dorsolaterally above M3. This rotation parallels the helical twist of the occlusal plane of the molar row (see below).
The dorsal surface of the maxilla ( figs. 7– 8 View Fig View Fig ) is divided into two major regions: (1) an endocranial region, representing the ventrolateral portion of the posterior rostral cavity; and (2) an orbitotemporal region, represent- ing the floor of the orbitotemporal fossa. These regions are separated by several maxillary contributions to the medial wall of the orbitotemporal fossa, namely its orbital lamina and palatine processes.
The endocranial surface features the nasopharyngeal sulcus, running along the medial side, and an elaborate series of depressions (herein termed the ‘‘maxillary sinus complex’’), situated anterolaterally ( fig. 8 View Fig ). The nasopharyngeal sulcus is relatively narrow throughout its length but narrows further in the zone between the molar rows. Anterior of the molar rows, the sulcus has an elevated medial rim that presumably supported the maxilloturbinate. Behind this point, the sulcus is unenclosed medially, indicating an undivided posterior nasopharynx. The maxillary sinus complex is a large and elaborate depression. It extends from just behind the premaxilla-maxilla sutural zone, back to above the front of M2, and laterally onto the root of the zygomatic plate, and it is partially subdivided by three low septa. The anterior septum is transverse and located well forward of the molar row. The central septum, also transverse, is level with the central loph of M1. The posterior septum is oblique, sharing a common medial origin with the central septum and terminating laterally at the level of the anterior loph of M2. Although the lateral margin of the maxillary sinus complex is mostly lost through damage, it is clear that it was enclosed by an extensive orbital lamina of the maxilla that crossed the root of the maxillary zygomatic plate. The floor of the maxillary sinus is marked by a short neurovascular sulcus that marks the passage of nasal branches of the infraorbital artery and superior alveolar nerve (see Discussion). On the left maxillary fragment, this sulcus passes obliquely across the lateral end of the anterior septum, while on the right fragment it is represented by an intraosseous canal exposed on the fractured edge. In various other murines surveyed by us (e.g., Rattus spp. , Mus musculus , Uromys caudimaculatus , Mammelomys lanosus , Paramelomys platyops , Bandicota indica ), this canal passes forward within the body of the maxilla to exit on the maxillary-premaxillary suture. The sutural surface in the fossil specimen is partially filled with matrix and the exact location of the anterior end of this canal cannot be determined without risk of damage.
The orbitotemporal surface takes the form of a narrow ledge above the posterior molars. This surface is irregular and includes several matrix-filled pits that probably represent foramina for branches of the superior alveolar nerves and vessels that supplied the upper molars. Anteriorly, it is delimited by the curving orbital lamina of the maxilla, and posteromedially, by two short palatine processes, aligned one in front of the other, and separated by a 1.75 mm gap. The orbital lamina and anterior palatine process are separated by a short sulcus that marks the position of the sphenopalatine foramen. The two palatine processes are rugose and represent sutural contacts with the orbital lamina of the palatine. The gap between the two palatine processes represents the dorsal palatine foramen. Medial to and aligned with this gap is a slitlike foramen that represents the orbital end of the intraosseous canal for the descending palatine artery and corresponding nerve.
The associated zygomatic plate ( fig. 6A–B View Fig ) is tall and relatively narrow, with a length of 9.7 mm at the narrowest point and a minimum height of 12.5 mm measured from the anteroventral border to the dorsal rim of the masseteric fossa. The anterior margin of the plate is thin and was either vertical or sloped slightly forward from the ventral root. The posterior margin of the plate is rounded and relatively thin, as appropriate to accommodate the lateral expansion of the maxillary sinus complex, as described above. The dorsal border of the masseteric fossa is more deeply excavated posteriorly than anteriorly. Although the dorsal root of the zygomatic arch is damaged posteriorly, enough survives to show that it was broad, with a shallow zygomatic notch that was probably no more than 1.5 mm in depth. The dorsal surface of the root is smooth and lacks the shallow fossa seen in some murines (e.g., Rattus spp. ) for insertion of the frontalis muscle. The attached, anterior portion of the zygomatic arch is extremely robust, with a depth of 4.5 mm. It is distinctly rugose at the point of union with the plate, presumably advertising the attachment of the medial zygomatic plate aponeurosis. This latter feature is also evident on two other fragmentary zygomatic plates referred at generic level (see below).
DENTARY: Three new dentaries are referred to C. buehleri ( fig. 9 View Fig ). Notable features of this element are the large size of the molars relative to the bony framework, the deep and robustly formed symphyseal process that surrounds a stout lower incisor, and the well-developed retromolar fossa. The more delicate processes of the posterior ramus are absent in each case; the articular condylar is preserved in the holotype ( Schaub, 1937: fig. 1 View Fig ).
The body of the ramus is deepest below the first molar, then shallows posteriorly along the molar row before deepening again onto the angular process. This creates a distinctly concave ventral border of the ramal body. The lateral surface bears a prominent and anteriorly expansive masseteric fossa, defined ventrally by a strong inferior masseteric crest and dorsally by a less prominent superior masseteric crest. The anterior union of these crests is located below the anterior root of m1 and is variably V-shaped or U-shaped. The mental foramen is situated at the intersection of the ramal body and the symphyseal process, just forward of the anterior root of m1, well below the diastemal border and well forward of the masseteric fossa.
The molars are large relative to the bony framework of the dentary, and the third molar is only slightly smaller than the second. To support the relatively large posterior molars, a prominent alveolar shelf overhangs the medial surface of the dentary. This shelf is linked to the medial surface of the ascending ramus by a well-developed postalveolar ridge that terminates at the mandibular foramen. The anterior margin of the ascending ramus arises lateral to the anterior root of m2, partially obscuring this tooth in lateral view. The trough between the molar row and the medial surface of the ascending ramus is narrow alongside m2 but opens posteriorly into a broad and well-developed retromolar fossa. This muscular insertion area is enclosed medially by the alveolar rim and posteriorly by the postalveolar ridge and presumably signifies hypertrophy of the posterior portion of the anteromedial portion of the anterior part of the temporal muscle.
Although the ascending ramus is damaged in all specimens, enough remains to demonstrate that the posterior end of the incisor alveolus is contained within the ascending ramus, without formation of a distinct tubercle. Damaged specimens indicate that the alveolus terminates level with the posterior end of the alveolar shelf and forward of a well-developed fossa for the posterior part of the medial layer of the masseter muscle.
The articular condylar, as preserved on the holotype and illustrated by Schaub, is supported by a broad condylar process, weakly incised behind by a broadly concave sigmoid notch. The condylar process on AMF 68831 ( fig. 9A View Fig ) is appropriately deep and stout. A low ridge on the lateral surface of the condylar process represents a continuation of the inferior margin of the muscular fossa. The medial surface of the condylar process is flat and lacks any equivalent buttressing. The angular process is also incomplete on all specimens. Surviving portions indicate that the internal pterygoid fossa was deeply excavated dorsally, below the postalveolar ridge, but smoothly continuous anteriorly with the medial surface of the ramal body. A small inferior mandibular foramen is preserved in two specimens, situated low and well forward in the internal pterygoid fossa.
The symphyseal process of the dentary rises at a steep angle relative to the molar row and ramal body. It is deep and robust, reflecting the stout nature of the lower incisor, and terminates posteroventrally in a prominent, triangular digastric process that has its apex located below the anteroconid of m1. The mandibular symphysis occupies much of the medial surface of the symphyseal process. It is broadest anteriorly and narrows to terminate on the digastric process. The anterior and posteroventral portion of this surface is highly rugose, marking the position of the symphyseal cartilage anteriorly, and of the anterior symphyseal ligament posteriorly. A discrete low ridge situated posterior and dorsal to the digastric process indicates the presence of a separate posterior symphyseal ligament. This ridge and the posterior end of the symphysis are separated by a sulcus that would have carried the submental artery and vein. Judging from the condition in other murines, the posteroventral rim of the digastric process would have provided attachment for two muscle units: anteromedially, for the aponeurosis of the anterior digastric muscle; and posterolaterally, for a straplike superficial portion of the transverse mandibular muscle. A small fossa located on the inner surface of the digastric process, just behind the symphysis, is interpreted as the point of origin of the deep portion of the transverse mandibular muscle. A low but continuous crest running along the dorsolateral margin of the symphyseal process marks the insertion of the pars orbicularis of the buccinator muscle.



| NMB |
Naturhistorishes Museum |
| R |
Departamento de Geologia, Universidad de Chile |
| JM |
Jura Museum, Eichstatt |
| MK |
National Museum of Kenya |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.