
Conothele isan, Decae & Schwendinger & Hongpadharakiree, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.4984.1.22 |
|
publication LSID |
lsid:zoobank.org:pub:26314FBC-18A7-4EBA-B0CF-5ADD5B03F5FD |
|
DOI |
https://doi.org/10.5281/zenodo.5205983 |
|
persistent identifier |
https://treatment.plazi.org/id/FB3B8783-2529-FFB0-FF7E-F94CFC81FF16 |
|
treatment provided by |
Plazi |
|
scientific name |
Conothele isan |
| status |
sp. nov. |
Conothele isan spec. nov.
Figs 6 View FIGURES 6 A−L, 7A−H
Type material. THAILAND: Buri Ram Province: Holotype ♂ (PS-005, sample TH-04/19, MHNG), Buri Ram District, Khao Kradong Forest Park, 14.9378°N, 103.0937°E, 180–300 m elevation, P.J. Schwendinger leg. as juvenile 16.+ 18.12.2004, matured in captivity 11.2.2005. GoogleMaps Paratypes: 2 ♀ paratypes ( 1 ♀: PS-006, sample TH-05/18, 15.12.2005, THNHM; 1 ♀: PS-007, sample TH-04/19, 16.+ 18.12.2004, MHNG), with same data as for holotype GoogleMaps .
Additional material examined. THAILAND: Roi Et Province: 1 ♀ (sample THKH-12/02, MHNG), Nong Phok District, Tham Pha Nam Thip Non-Hunting Area , 16.3644°N, 104.3258°E, 420 m elevation, P.J. Schwendinger leg. 12.+ 15.6.2013. GoogleMaps
Etymology. The species epithet, a name in apposition, refers to northeastern Thailand, locally called the Isan (= Isarn or Esarn), the geographic region where this species was discovered.
Diagnosis. Different in both sexes from C. martensi spec. nov. by the profile of the carapace with a relatively less strongly elevated cephalic part sloping gradually down past the fovea and continuing in a straight line over the thoracic part to the posterior margin of the carapace ( Figs 6B View FIGURES 6 , 7B View FIGURES 7 ), and by the spermathecae that are hooked (rather than twisted) in the median portion and have the globular heads directed inwards rather than forwards ( Figs 7 View FIGURES 7 F−H cf. Figs 5 View FIGURES 5 F−K). Further differences are found in the ventro-lateral spine patterns on the anterior metatarsi and tibiae of males (more spines present; retroventral spine row on tibia II present vs. absent in C. martensi spec. nov.; Figs 6 View FIGURES 6 D−E cf. Figs 4 View FIGURES 4 D−E), in the morphology of the embolus (without abrupt narrowing at midpoint; Figs 6I, K View FIGURES 6 cf. Figs 4I, K View FIGURES 4 ) and in the PLE being relatively large in comparison to the ALE (PLE/ALE = 0.6 in C. isan spec. nov. vs 0.7 in C. martensi spec. nov.).
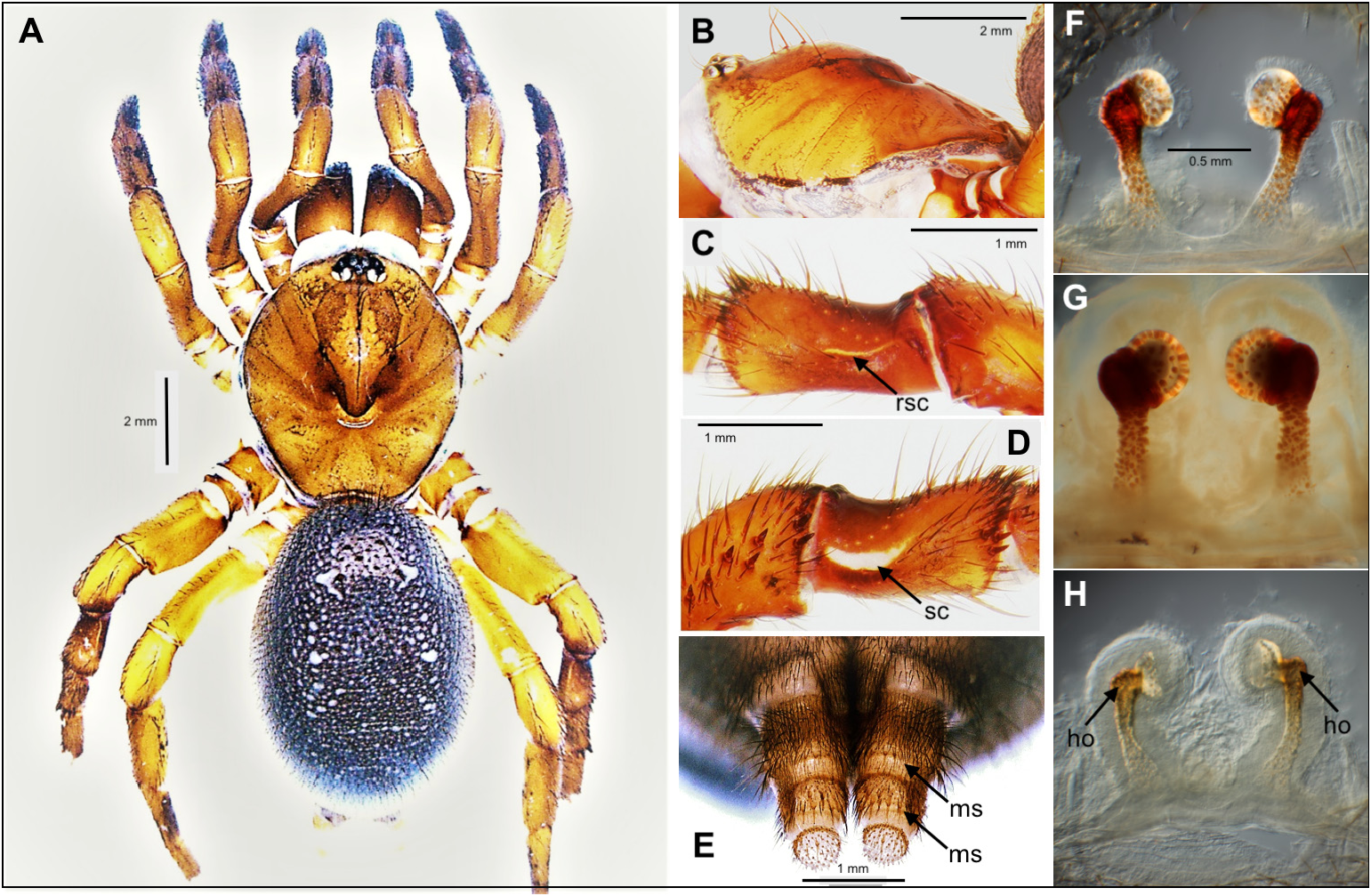
Description. Male ( holotype). Specimen preserved for 16 years in 70% ethanol in good condition ( Fig. 6A View FIGURES 6 ); right palpal organ removed for study and kept in microvial. General colouration: carapace and chelicerae blackish brown; dorsal side of opisthosoma dark grey, with fine light grey speckles, ventral side anterior of epigastric furrow, book-lung covers and spinnerets cream-coloured; legs yellowish brown, anterior legs slightly darker than posteriors; palps dark brown; sternum yellow, with sharply outlined brown margin.
Morphology: Carapace ( Figs 6 View FIGURES 6 A−B), except for lower profile of cephalic part (see diagnosis), as described for C. martensi spec. nov. Ocular tubercle steep; clypeus short; eye group rectangular, twice as wide as long (EL/PR = 0.50); AME less than their diameter apart from each other (disAME/diaAME = 0.72); PR straight. Chelicera with small apical rastellum; fangs smooth (possibly a diagnostic difference from C. martensi spec. nov., its male holotype with serrated fangs; see above). Palpal coxae with 16 cuspules spread over proximal half of article; prolateraldistal lobe short and rounded. Labium with two distal cuspules; labio-sternal suture deep and procurved. Sternum and most parts of palps (except for palpal organ) as in male of C. martensi spec. nov. Palpal organ ( Figs 6 View FIGURES 6 H−L) with bulbous proximal part almost as wide as long (BuW/GL = 0.96); embolus thin and evenly tapering, with a harpoon-shaped tip ( Fig. 6L View FIGURES 6 ), mostly as in C. martensi spec. nov. but without uneven narrowing halfway. Legs I−II: scopulate tarsi proximally inflated ( Fig. 6C View FIGURES 6 ); short lateral spines present on tarsi, metatarsi and tibiae; few clavate trichobothria dorsally in proximal half of tarsi, more ventro-lateral spines on metatarsi and tibiae of anterior legs than in C. martensi spec. nov. ( Figs 6 View FIGURES 6 D−E cf. Figs 4 View FIGURES 4 D−E). Leg III with shallow, glabrous saddle-shaped dorsal depression in tibia, saddle crescents indistinct/absent ( Figs 6F–G View FIGURES 6 ); spine groups dorso-distally on metatarsus and tibia; patella with strong, short spines prodorsally. Leg IV slender, with weak spines on metatarsus; metatarsus longer than tibia (MetIV/TibIV = 1.2). PTC of anterior legs with a single proximal tooth, of posterior legs with one large and one small tooth. Leg formula 4123. Opisthosoma ovoid, anteriorly narrowing, carrying evenly spread, short stiff bristles; wart-like sockets not evident (possibly a diagnostic difference from C. martensi spec. nov., its male types possessing wart-like sockets; see above). Spinnerets as in C. martensi spec. nov.
Measurements. TBL 12.4; CL 5.4; CW 5.1; CP 3.7; AR 1.17; PR 1.17; EL 0.59; diaALE 0.35; diaPLE 0.21; diaAME 0.18; diaPME 0.22; disALE 0.64; disPLE 0.87; disAME 0.13; disPME 0.43; SL 3.1; SW 2.7; LL 0.5; LW 1.0; palp 8.6 (1.1 + 2.5 + 1.8 + 3.2); leg I 12.5 (1.1 + 1.8 + 2.7 + 2.3 + 4.6); leg II 11.3 (1.3 + 1.7 + 2.3 + 2.1 + 3.9); leg III 10.1 (1.5 + 1.6 + 2.0 + 1.8 + 3.2); leg IV 13.4 (1.6 + 2.9 + 2.5 + 2.1 + 4.3); BuL 1.92; BuW 0.75; EmL 1.14.
Female ( paratype; PS-007): Specimen preserved for 16 years in 70% ethanol in good condition ( Fig. 7A View FIGURES 7 ); spermathecae dissected and stored in microvial.
General colouration: carapace, chelicerae, legs and palps brown; dorsal side of opisthosoma dark grey, with fine light grey speckles, ventral side, book-lung covers and spinnerets yellowish white; sternum yellow, with sharply outlined brown margin; carapace and sternum with distinct grey shading.
Morphology: Carapace ( Figs 7A–B View FIGURES 7 ) smooth, longer than wide (CW/CL = 0.9), otherwise as described for male. Eyes as in male. Chelicerae carrying an apical rastellum composed of a curved row of 9–10 strong teeth on a low process; fangs ventrally with fine serration. Palpal coxae with 15–17 cuspules spread over proximal half of article; prolateral-distal lobe rudimentary. Labium, labio-sternal suture and sternum as in male. Labium with two distal cuspules. Palps as described above for Ummidiinae ; claw with strong and bifid proximal tooth. Legs I−II as described above for Ummidiinae . Leg III with tarsus carrying a dense disto-ventral group of spines; metatarsus with dorsodistal group of very strong short spines; tibia with short rows of spines along dorso-distal margin, saddle-shaped depression prolaterally bordered by relatively wide saddle crescent ( Fig. 7D View FIGURES 7 ), retrolaterally by a narrower one ( Fig. 7C View FIGURES 7 ); patella with short strong spines along anterior margin and on prolateral side; femur ventro-proximally inflated; distinct opposing protuberances on dorsal and prolateral sides of trochanter and coxa. Leg IV with patella carrying a prodorsal-proximal group of short stiff bristles. PTC as described above for Ummidiinae . Leg formula 4123. Dorsal surface of opisthosoma with wart-like bristle sockets. Spinnerets ( Fig. 7E View FIGURES 7 ) as in C. martensi spec. nov. Spermathecae tripartite; proximal part narrow and lightly pigmented; median part strongly pigmented, bent inwards, distally widening; distal part globular and inwards-directed ( Fig. 7F View FIGURES 7 ). Similar to spermathecae of Conothele baisha and C. baoting from Hainan Island, China ( Liu et al. 2019: figs 5G, 6A–F, 7G), of C. daxinensis from Guanxi, China ( Xu et al. 2017: fig. 2E) and of C. vali from India ( Siliwal et al. 2009: fig. 27).
Measurements. TBL 15.7; CL 6.0; CW 5.3; CP 4.1; AR 1.25; PR 1.31; EL 0.66; diaALE 0.42; diaPLE 0.27; diaAME 0.17; diaPME 0.30; disALE 0.57; disPLE 0.92; disAME 0.12; disPME 0.45; SL 3.3; SW 3.1; LL 0.8; LW 2.0; palp 9.0 (1.9 + 1.9 + 1.9 + 3.3); leg I 10.4 (1.0 + 1.4 + 2.1 + 2.4 + 3.5); leg II 9.4 (1.1 + 1.2 + 1.7 + 2.3 + 3.1); leg III 9.1 (1.4 + 1.3 + 1.5 + 1.9 + 3.0); leg IV 11.4 (1.3 + 2.2 + 2.1 + 2.2 + 3.6).
Variation. Females (n = 2): TBL 14.5−15.7; CL 5.3−6.0; CW 4.8−5.3; CP 3.4−4.1;AR 1.10−1.25; PR 1.13−1.31; EL 0.59–0.66; diaALE 0.35−0.42; diaPLE 0.24−0.27; diaAME 0.17−0.19; diaPME 0.22−0.30; disALE 0.51−0.57; disPLE 0.79−0.92; disAME 0.08−0.12; disPME 0.31−0.45; SL 3.0−3.3; SW 2.7−3.1; LL 0.6−0.8; LW 1.0−2.0; palp 7.9−9.0; leg I 8.7−10.4; leg II 8.0−9.4; leg III 7.5−9.1; leg IV 8.8−11.4. Variation in the shape of the spermathecae of three females is shown in Figs 7 View FIGURES 7 F−H.
Habitat and biology. The type specimens were collected from roadsides in a seasonally dry deciduous forest between 180 and 300 m elevation, on an inactive and strongly eroded volcano. The non-type female is from a roadside in a lush mixed evergreen-deciduous forest at 420 m elevation. The spiders lived in short burrows (maximally 3.5 cm long) lined with very dense silk and closed by a thin (wafer-type) trapdoor with a maximal length of 1.1 cm and a maximal width of 1.6 cm. This burrow structure is different from that of the co-occurring Latouchia incerta spec. nov. The male holotype matured in captivity (in Geneva) in early February, less than two months after being captured. The mating season thus appears to be at the end of the cool part and into the hot part of the dry season. Spiderlings presumably hatch and emerge from maternal burrows at the beginning of the rainy season when prey becomes more abundant again.
The types were collected together with a single female of a second Conothele species (with strongly coiled spermathecae) which lived in a short burrow with a second thin trapdoor opening into a blind chamber behind the bottom. Two Conothele species also co-occur at the type locality of C. martensi spec. nov., and at the type locality of C. isan spec. nov. even a third ummidiine species can be found, L. incerta spec. nov. This is quite remarkable.
Notes. The difficulty in distinguishing some Ummidiinae taxa at the genus level (as discussed above) is also encountered at the species level; C. martensi spec. nov. and C. isan spec. nov. are difficult to distinguish on morphological characters alone. Particularly the male palpal organ, often the morphologically most characteristic structure used to distinguish and taxonomically classify mygalomorph spider species, is very similar in these two species ( Figs 6 View FIGURES 6 H−L cf. Figs 4 View FIGURES 4 H−L). In both species the globular proximal part and the slender, harpoon-tipped embolus are virtually identical in shape. A similar palpal organ was described by Yang & Xu (2018) for C. dequin from China ( Yunnan Province). Genetics-based research is required to establish the phylogenetic relationships between the Thai species and C. dequin , and to show if their morphological similarities are not due to convergent evolution.
Female genitalia of Conothele spp. show an equally low diversity in shapes and forms. The two different forms present in C. martensi spec. nov. and C. isan spec. nov. are also found in several other, geographically distant, named species ( Siliwal et al. 2009; Xu et al. 2017; Liu et al. 2019), as well as in Conothele females from numerous localities in SE-Asia where the corresponding males are distinct (personal observations). Especially the hooked type of spermathecae of C. isan spec. nov. is very common. Therefore the description of new Conothele species on the basis of only one sex (particularly the female) is not advisable.



| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ummidiinae |
|
Genus |