
Hapalotremus, Simon, 1903
|
publication ID |
https://doi.org/ 10.1080/00222933.2018.1506521 |
|
publication LSID |
lsid:zoobank.org:pub:1C028676-CCD9-4654-A755-6ACA3BCE3E98 |
|
persistent identifier |
https://treatment.plazi.org/id/FD618785-AA41-CA30-1BB2-C836FD4A6699 |
|
treatment provided by |
Carolina |
|
scientific name |
Hapalotremus |
| status |
|
Genus HAPALOTREMUS Simon, 1903 View in CoL
( Figures 1–37 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 View Figure 16 View Figure 17 View Figure 18 View Figure 19 View Figure 20 View Figure 21 View Figure 22 View Figure 23 View Figure 24 View Figure 25 View Figure 26 View Figure 27 View Figure 28 View Figure 29 View Figure 30 View Figure 31 View Figure 32 View Figure 33 View Figure 34 View Figure 35 View Figure 36 View Figure 37 )
Dolichothele Mello-Leitão, 1923: 120 ; Raven 1985: 152.
Goniodontium Mello-Leitão, 1923: 127 . Raven 1985: 153.
Cyclothoracoides Strand 1929: 17 ; Raven 1985: 151.
Type species
Hapalotremus albipes Simon, 1903 View in CoL by monotypy.
Species included
Hapalotremus albipes Simon, 1903 View in CoL , Hapalotremus carabaya View in CoL sp. nov., Hapalotremus chasqui View in CoL sp. nov., Hapalotremus chespiritoi View in CoL sp. nov., Hapalotremus kuka View in CoL sp. nov., Hapalotremus major ( Chamberlin, 1916) View in CoL , Hapalotremus marcapata View in CoL sp. nov., Hapalotremus martinorum Cavallo and Ferretti, 2015 View in CoL , Hapalotremus perezmilesi View in CoL sp. nov., Hapalotremus vilcanota View in CoL sp. nov.
Etymology
The Greek adjective Hapal, followed by the Greek connective vowel o, followed by the Greek noun trêma, followed by the Latin suffix us meaning of or pertaining to. Hence, the derivative compound name Hapalotremus ‘of the soft hole’, possibly refers to the way it lives in a soft hole in the ground. Gender would be masculine.
Generic diagnosis
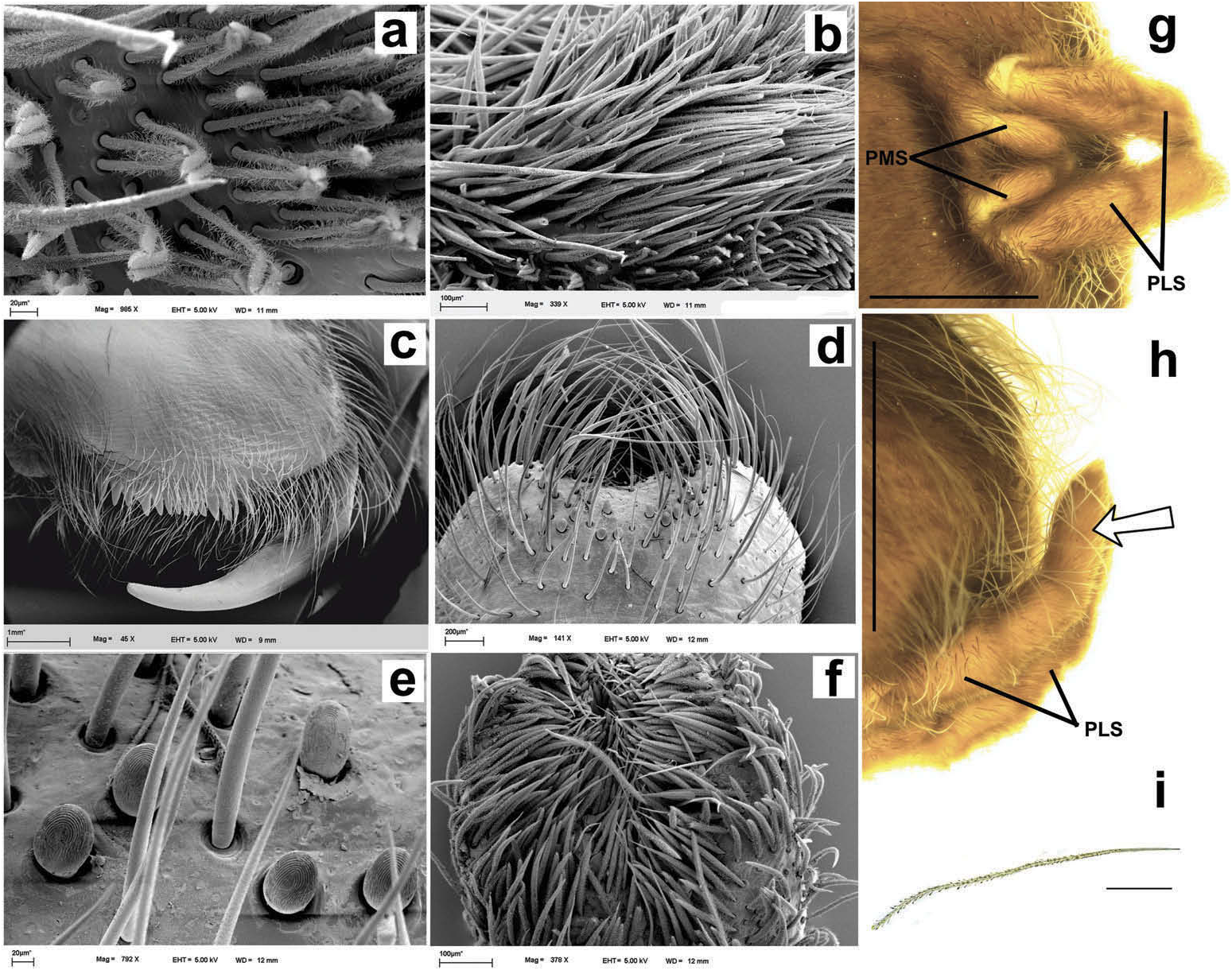
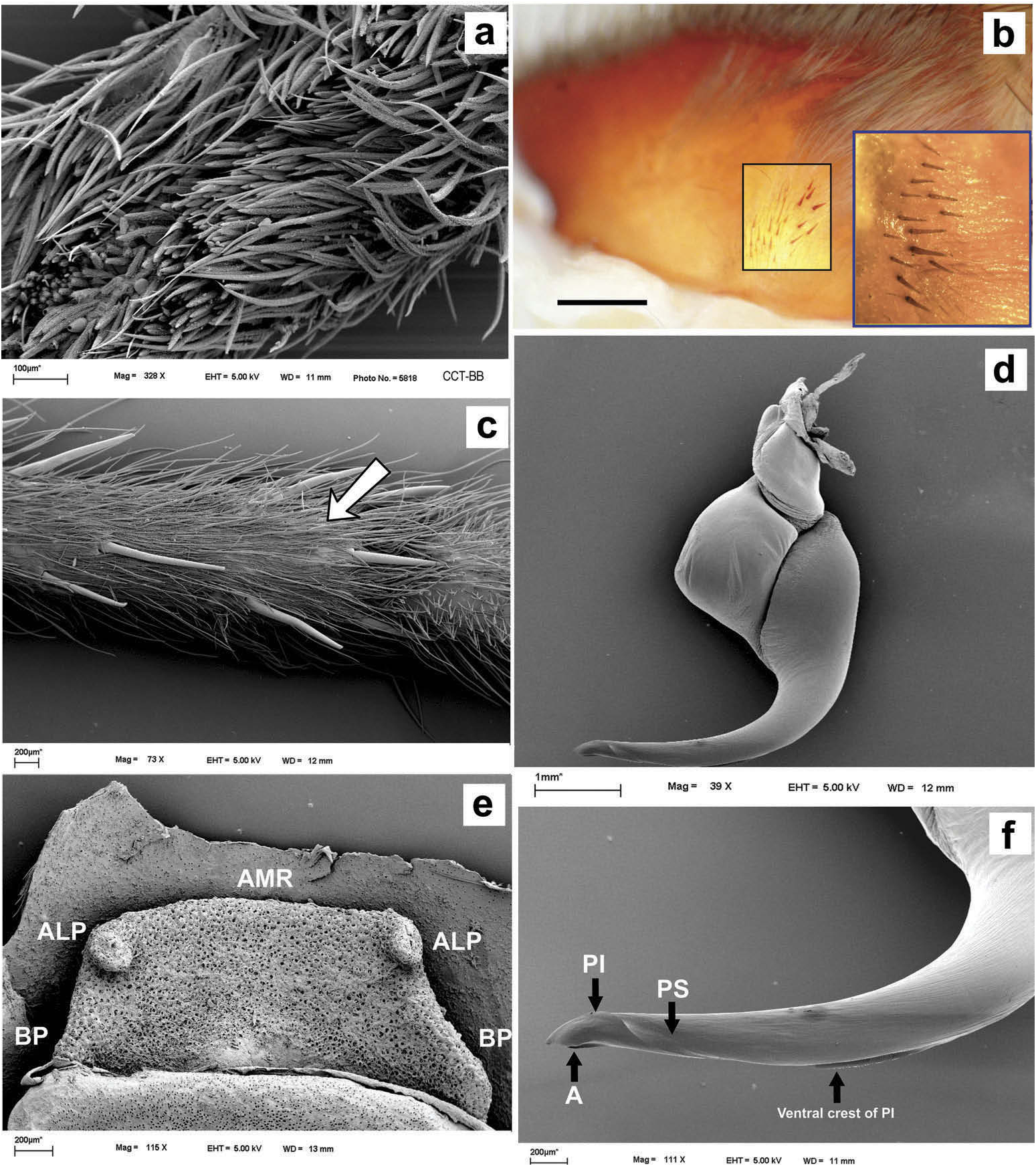
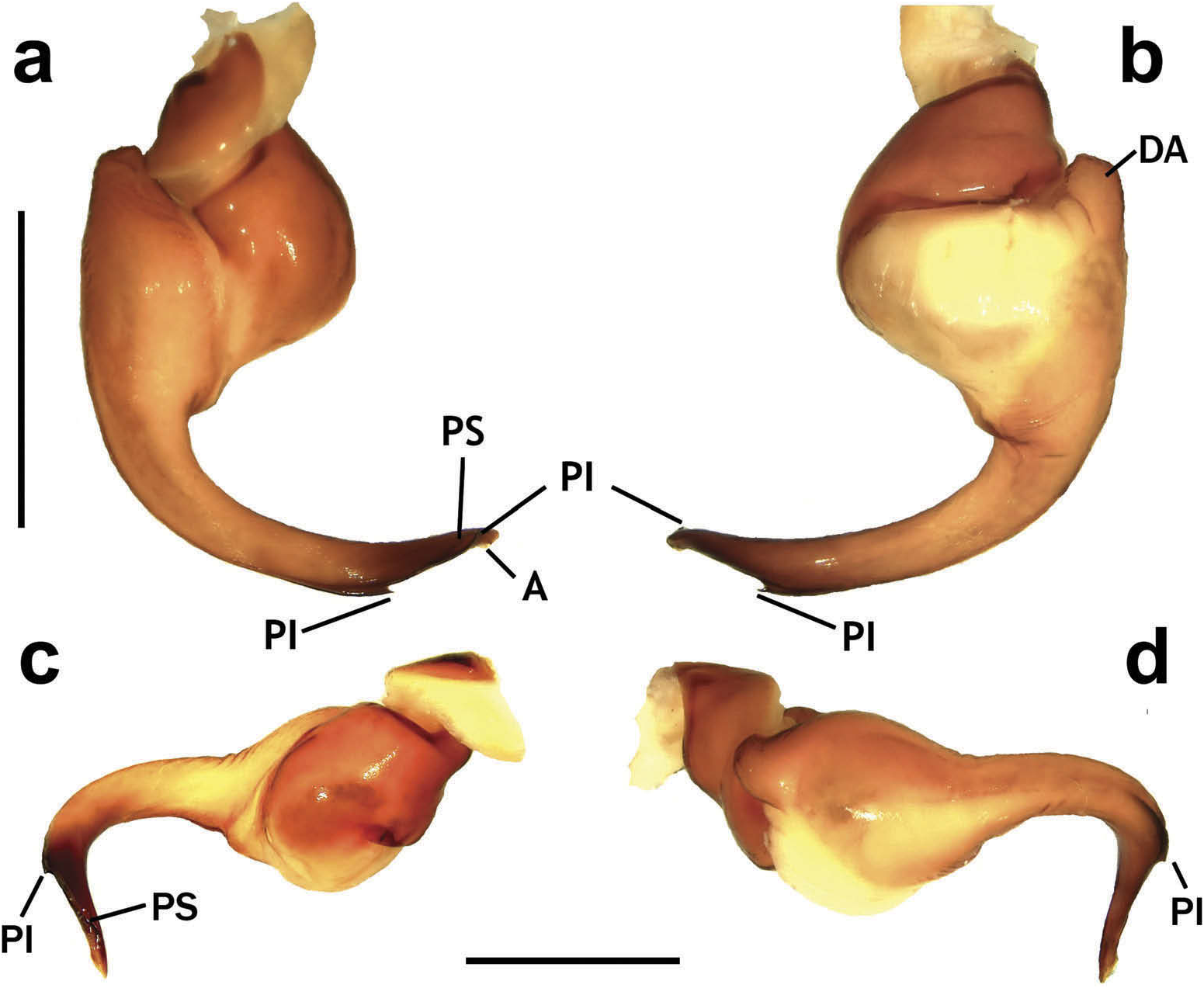
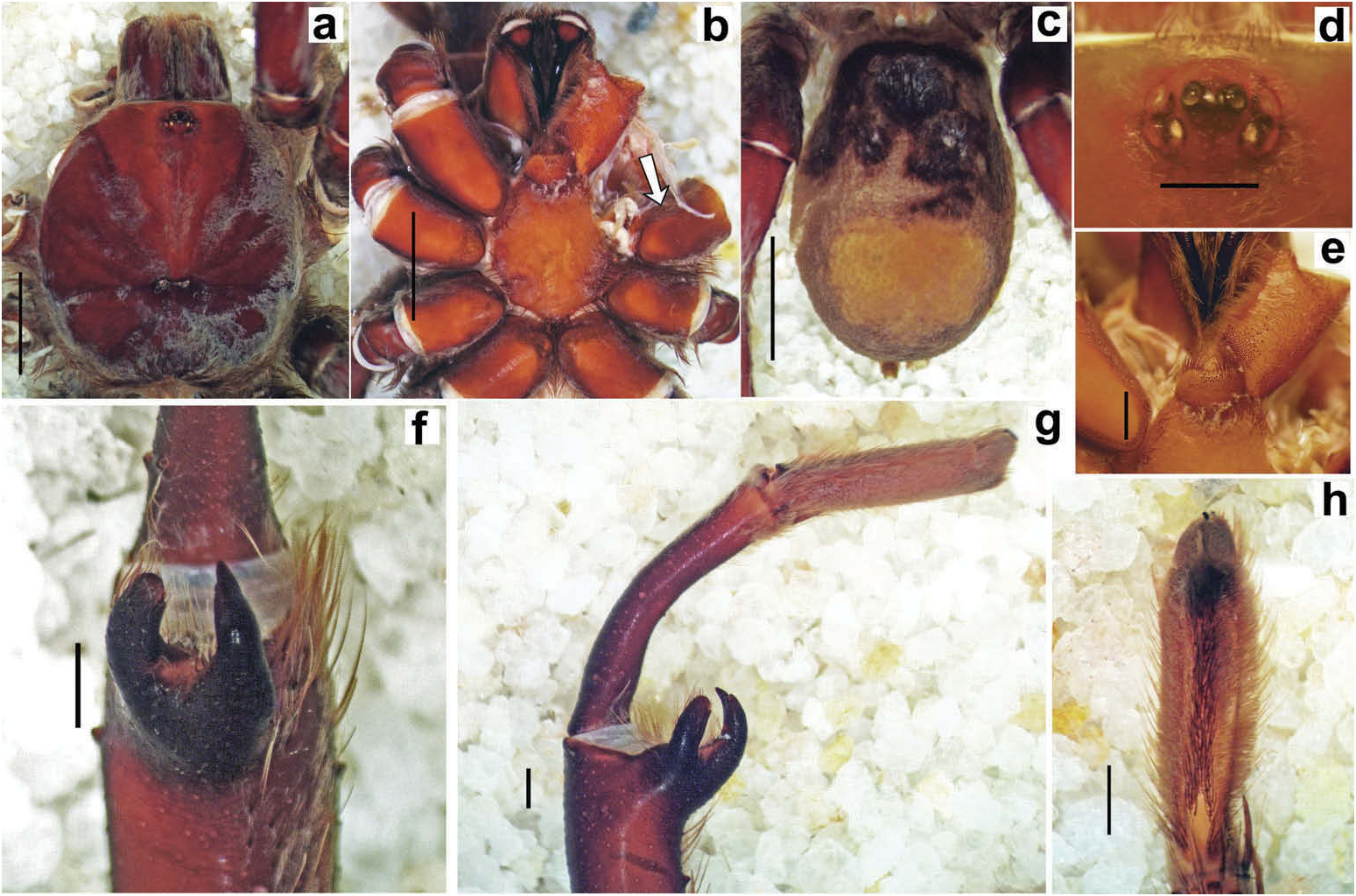
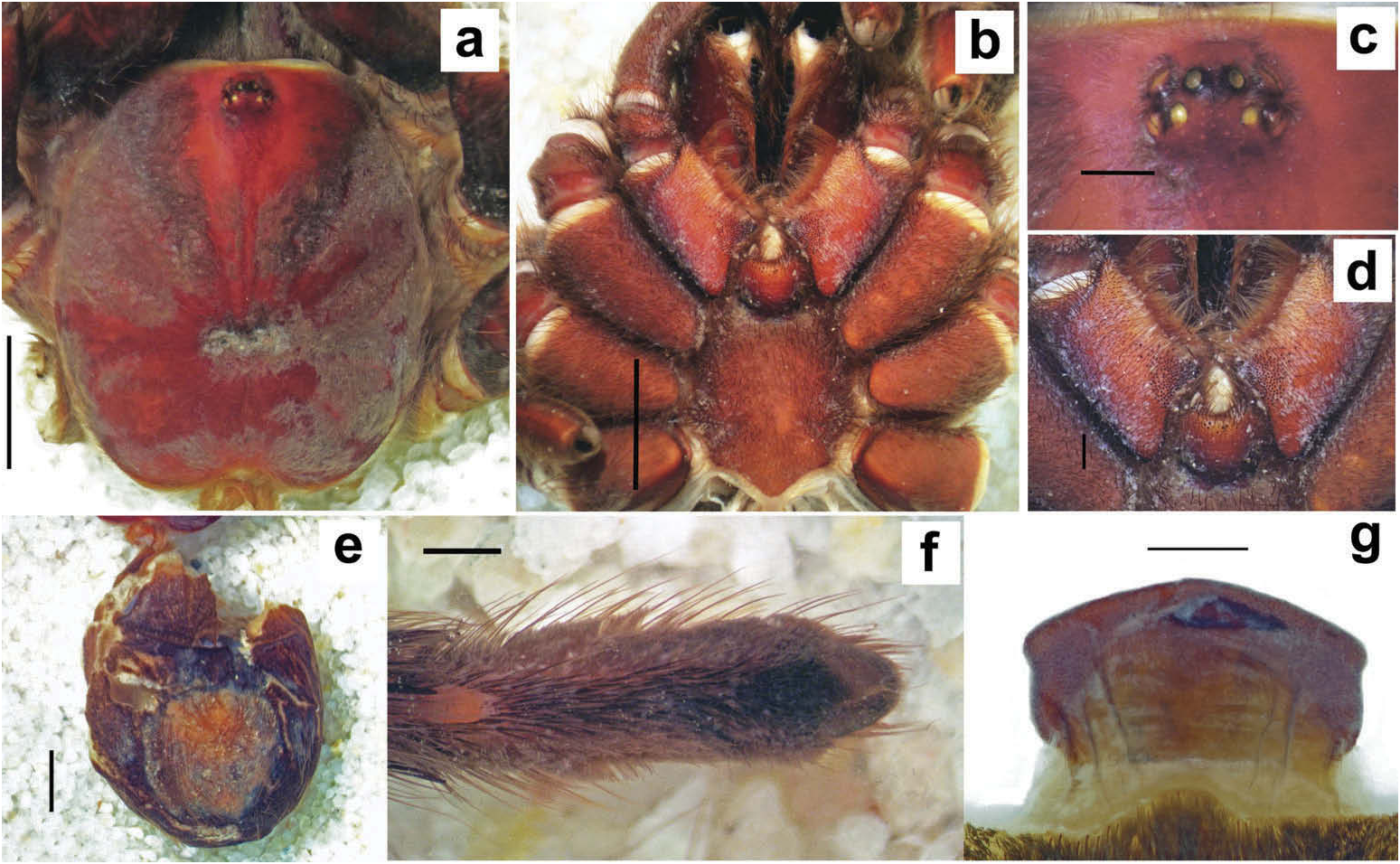
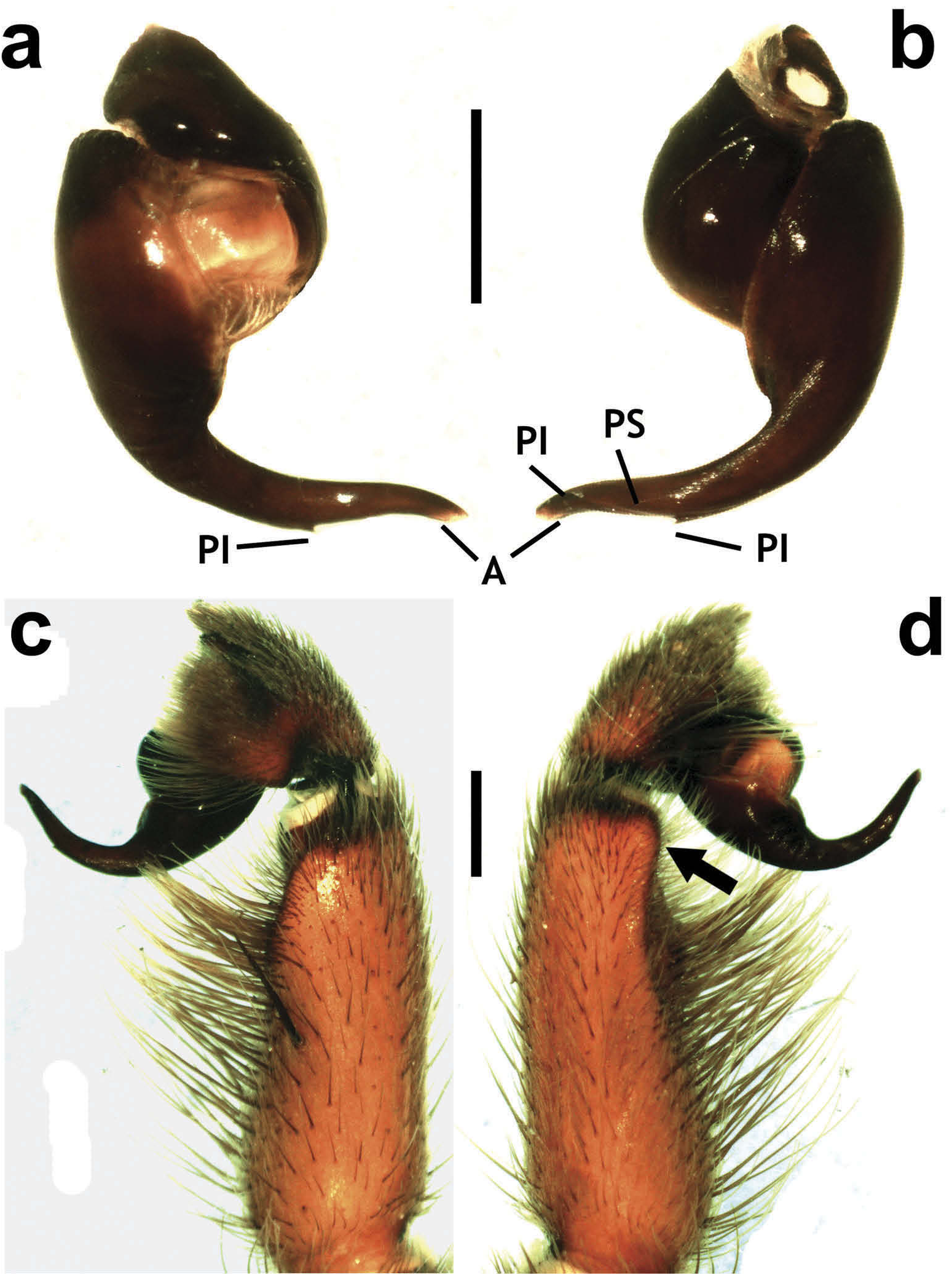
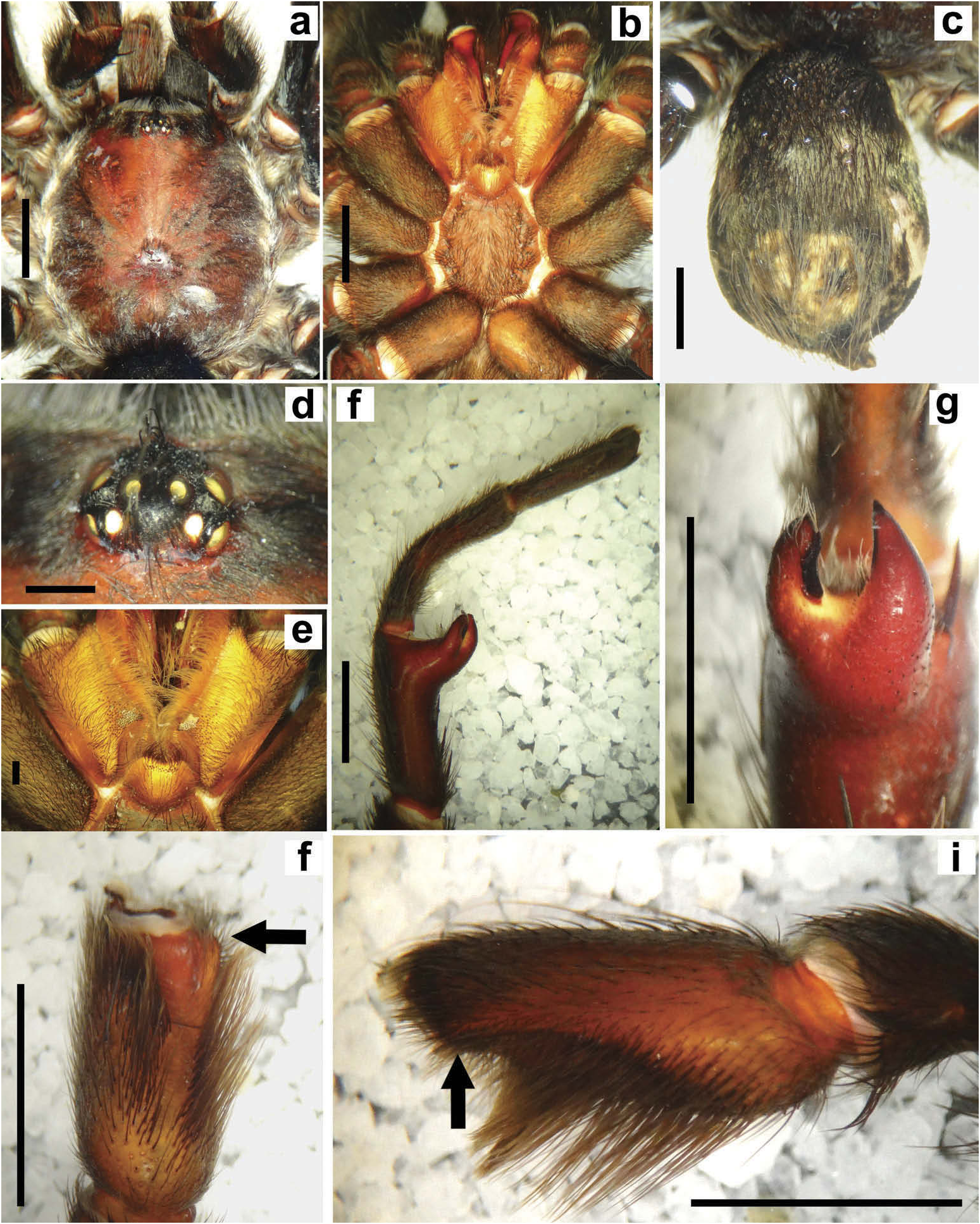
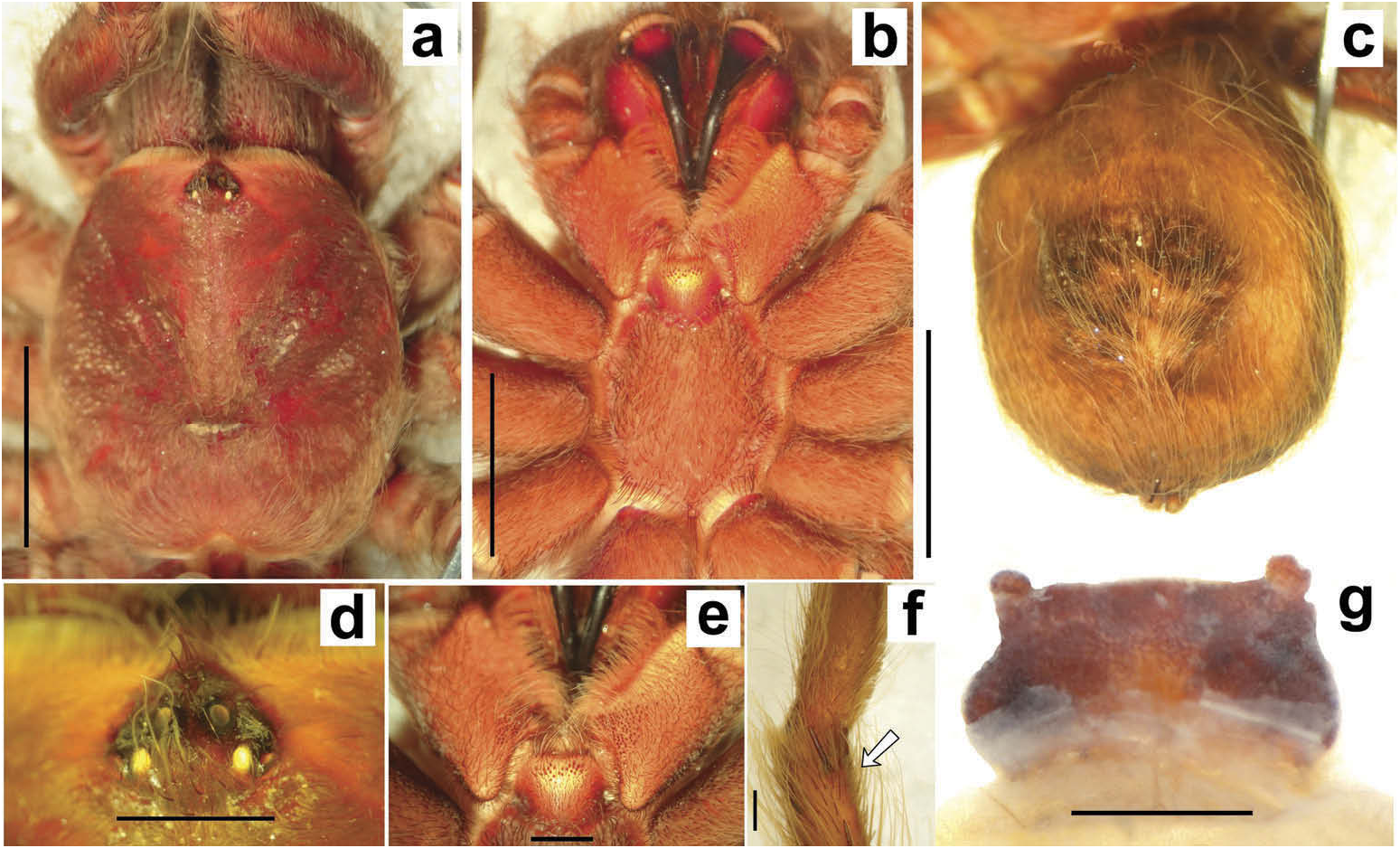
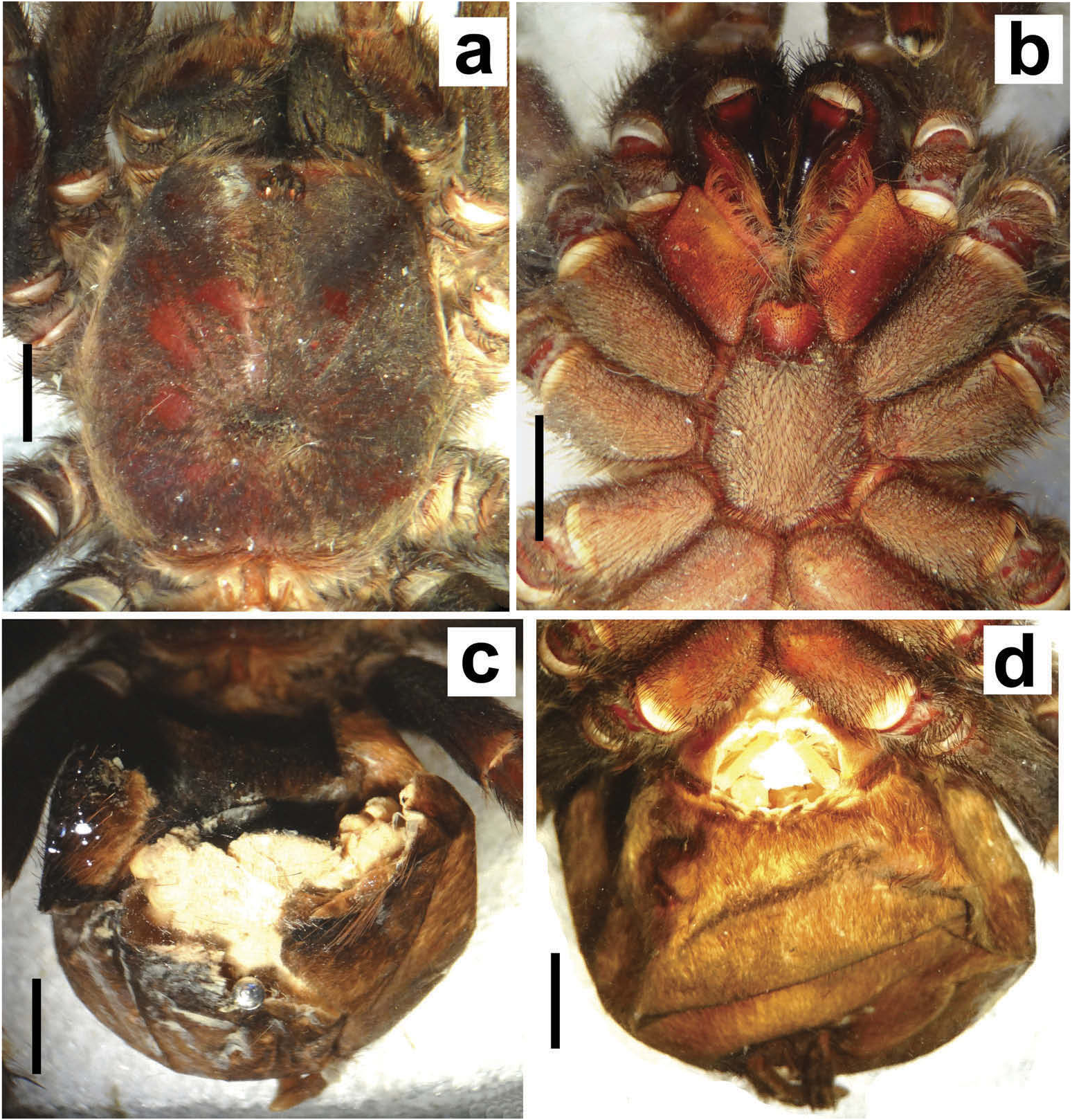
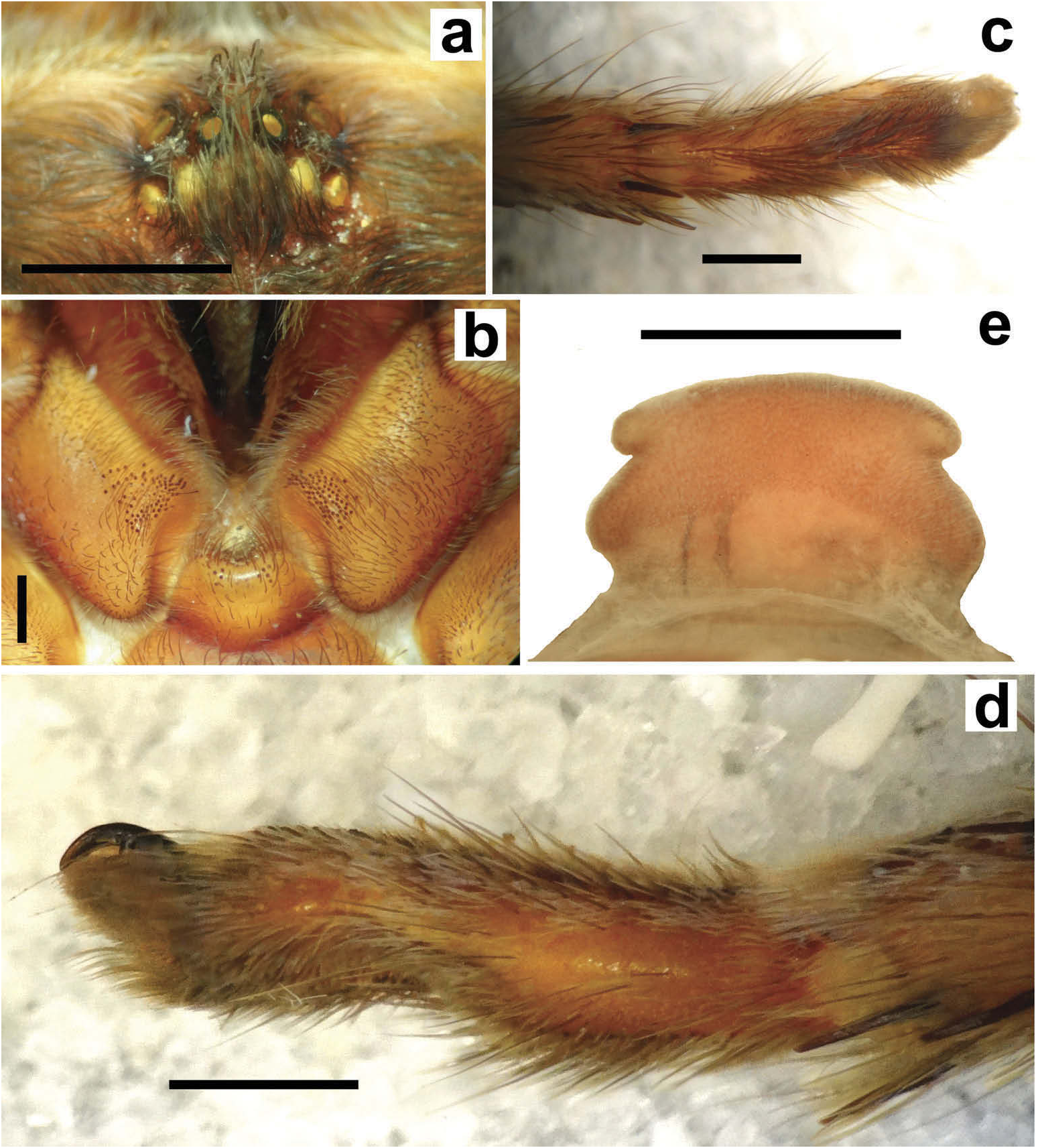
Hapalotremus resembles Hapalopus Ausserer, 1875 and Davus Pickard-Cambridge, 1892 by having the metatarsus I strongly curved ( Pérez-Miles et al. 1996; Bertani 2000; Gabriel 2016). However, males of Hapalotremus differ from those of Davus in the shape of male palpal bulb without a twisted embolus and from those of Hapalopus by having a longer and curved embolus with a continuous PI (split in two in Hapalopus ), also by a shorter tibia I and by the shape of the spermathecae on females without having a membranous base. Hapalotremus also resembles Schizopelma Pickard –Cambridge, 1897; since females have one single spermathecal receptacle ( Pérez-Miles et al. 1996; Gabriel 2016), but differ in male characteristics: lacking a bifid embolus on palpal bulb and tibial apophysis with two branches. Additionally, representatives of Hapalotremus can be characterized by the combination of the following characters: presence of red or white setae on the dorsum of the abdomen ( Figures 1 View Figure 1 , 2 View Figure 2 ) and a labium bearing a reduced number of cuspules (about 4 to 12) ( Figure 5d, e View Figure 5 ), excepting on H. martinorum , H. chasqui sp. nov. and H. kuka sp. nov. (more than 20 cuspules). Males and females are also characterized by presenting a slightly developed scopula or no scopulae on apical metatarsus IV ( Figure 6c View Figure 6 ) and by having a small field of strong short spiniform setae on basal retrolateral coxal face of palp and legs I–IV ( Figure 6b View Figure 6 ). Additionally, males are further distinguished by showing the following combination of characters: short tibia I, more than 10% shorter than metatarsus I ( Figure 7g View Figure 7 ) with the exception of H. martinorum ( Figure 13g View Figure 13 ) and H. major ( Figure 10g View Figure 10 ) (tibia I roughly the same length as metatarsus I or longer), tibia I with two apophyses which originate from a common base ( Figures 7f View Figure 7 , 10f View Figure 10 ), curved metatarsus I ( Figures 7g View Figure 7 , 10g View Figure 10 ), presence of a rounded retrolateral process on male palpal tibia ( Figures 7h,i View Figure 7 , 11c,d View Figure 11 , 13h,i View Figure 13 ), palpal bulb with digitiform apophysis ( Figure 8b View Figure 8 ) and prolateral inferior keel presenting a ventral crest (blunt or pointed) ( Figure 6d, f View Figure 6 ). Females are distinguished by having a single oblong spermathecal receptacle with a basal portion usually wider than apically and the presence of basal and apical lateral projections ( Figure 6e View Figure 6 ).
Distribution and habitat
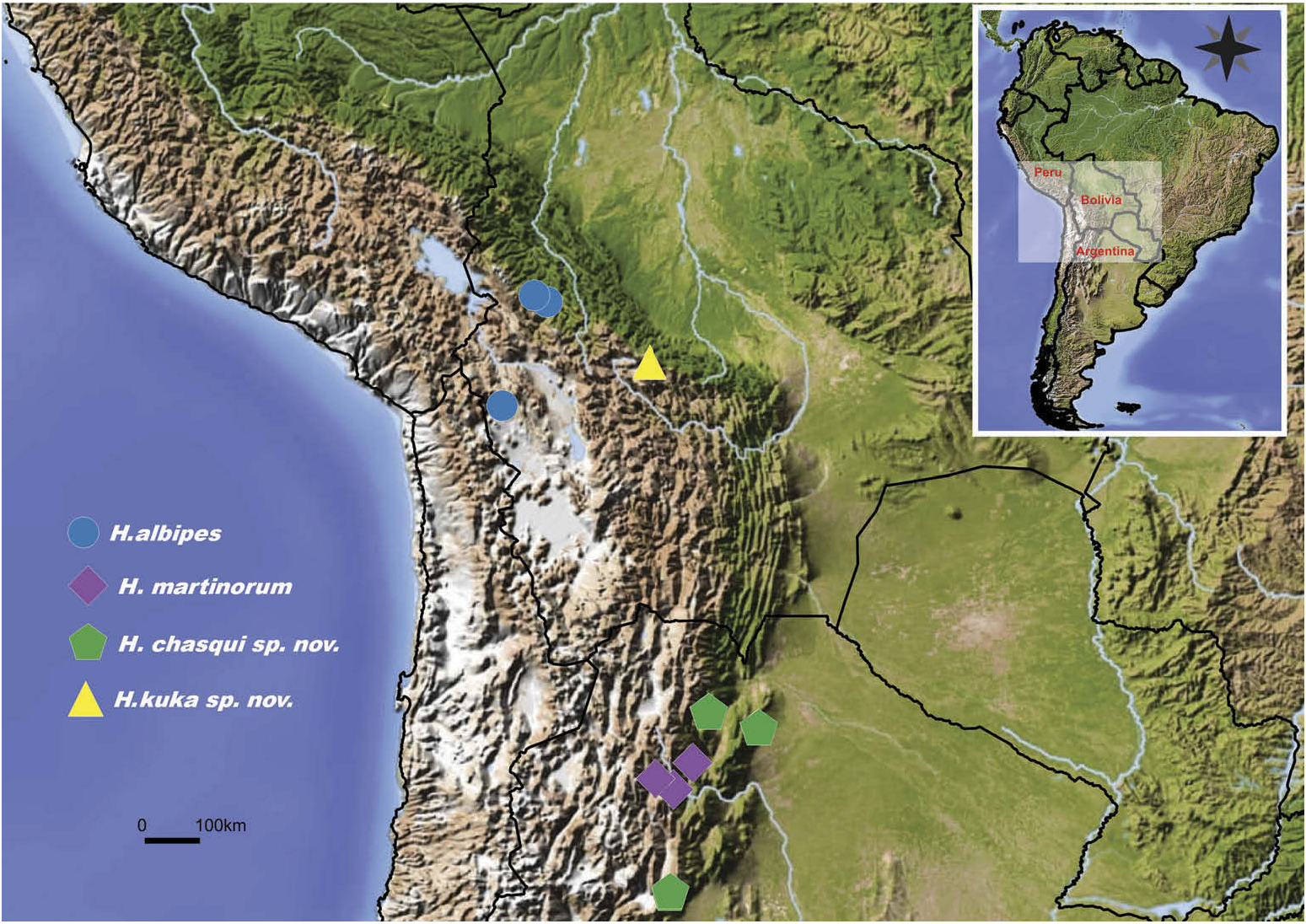
The genus is endemic to South America and is mostly distributed in the western Andean region from central Peru in the north to northwestern Argentina in the south ( Figures 36 View Figure 36 , 37 View Figure 37 ). Specimens of Hapalotremus clearly show habitat preferences involving high-altitude cloud forests ( Figure 3a–c View Figure 3 ) and grassy rocky mountains ( Figure 3d–j View Figure 3 ). Specimens can be found among fallen logs or under rocks; they make short burrows like other theraphosid spiders ( Figure 2e, f View Figure 2 ), with one or more co-existing. Some species can be found at very high altitudes in the Andean region, with records of more than 4500 m above sea level (a. s.l.) ( Figure 3d–j View Figure 3 ). As far as we know, Hapalotremus are the only known theraphosid spiders in the world found living at such high elevation (up to 4524 m a.s.l.). Some of the highest elevations recorded for Theraphosidae are those of Haplocosmia himalayana ( Pocock, 1899) (about 2100 m a.s.l.) ( Pocock 1899), Aguapanela arvi Perafán, Cifuentes and Estrada –Gómez, 2015 (up to 2400 m a.s.l.) ( Perafán et al. 2015), Cyriocosmus spp. (up to 3000 m a.s.l.) ( Pérez-Miles and Weinmann 2009), Euathlus spp. (up to 3200 m a.s.l.) ( Perafán and Pérez-Miles 2014; Ferretti 2015) and recently described species of Bistriopelma Kaderka, 2015 with elevation records up to 4398 m a.s.l. ( Kaderka 2015).
Description
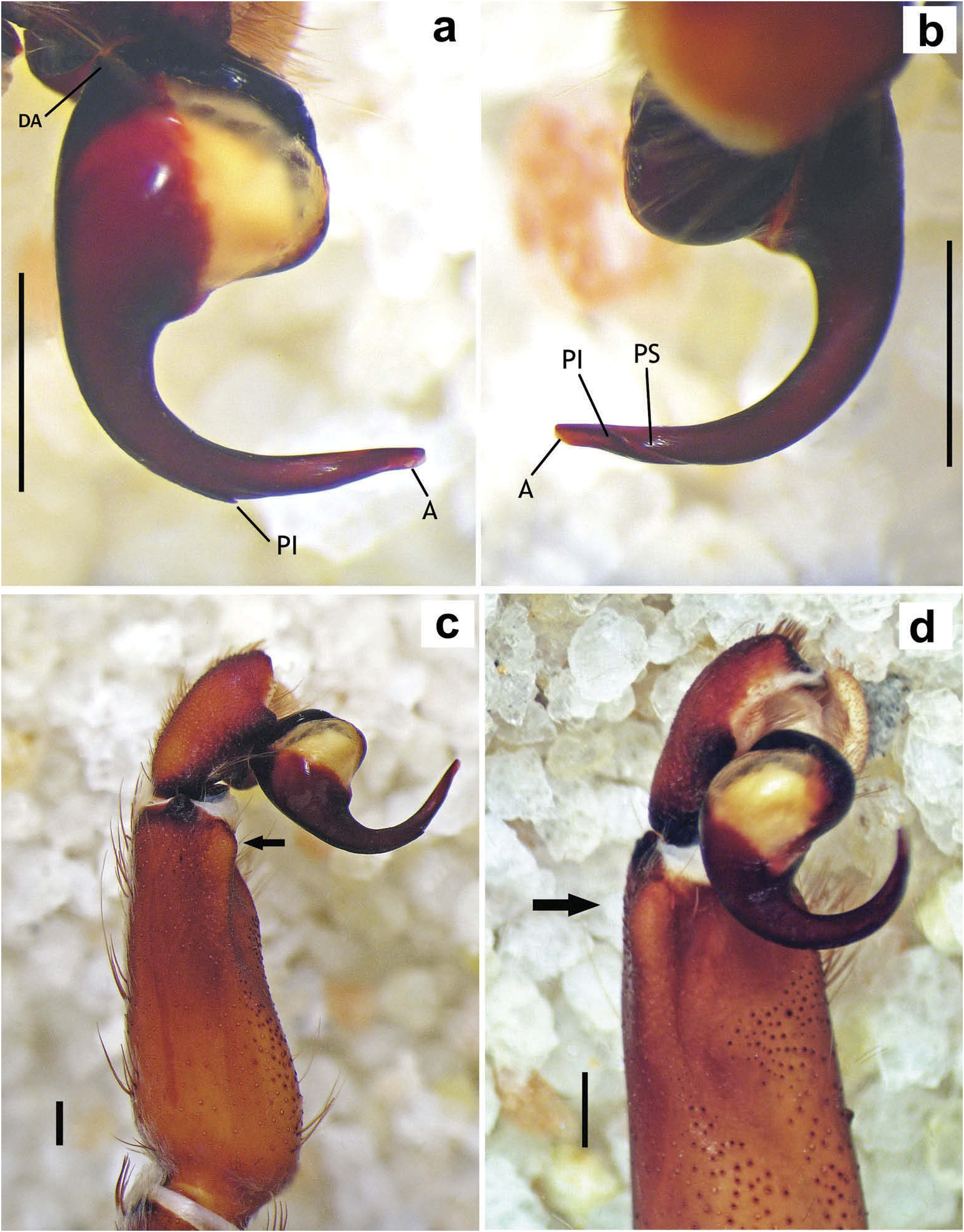
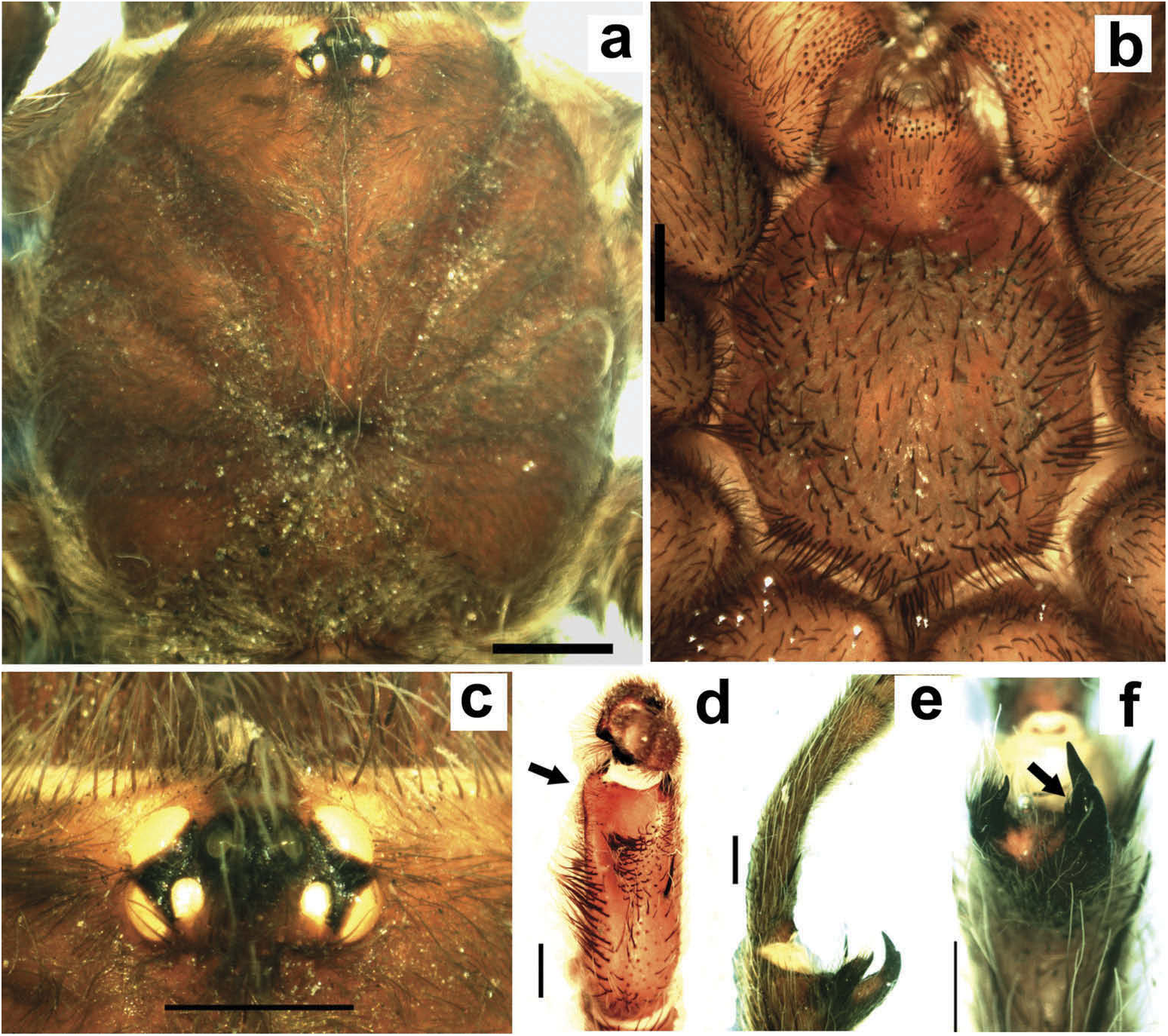
Chelicerae without rastellum ( Figure 5c View Figure 5 ). Clypeus narrow. Overall shape of eye group rectangular ( Figures 9g View Figure 9 , 10d View Figure 10 , 12c View Figure 12 , 13d View Figure 13 ). Eye tubercle elevated, wider than long. Anterior eye row procurved, posterior recurved. Males and females with oval carapace, both with fovea straight or slightly procurved ( Figures 7a View Figure 7 , 9a View Figure 9 , 12a View Figure 12 ). Intercheliceral tumescence absent. One row of big teeth on cheliceral promargin furrow and a group of small teeth on retromargin ( Figure 5c View Figure 5 ). Labium slightly wider than long or subequal, with about 4–27 conspicuous blunt cuspules on the subapical margin ( Figure 5d, e View Figure 5 ). Endites with distinct anterior lobe, with 53–126 blunt cuspules on inner basoventral surface. Sternum oval, six sternal sigillae, median and posterior away from margin by about one length or next to margin, oval-elongated or oval-rounded in H. carabaya sp. nov. Urticating hairs: typical type III ( Figure 5i View Figure 5 ) occurring in a patch on central dorsum of the abdomen ( Figures 1 View Figure 1 , 2a–d View Figure 2 ). Stridulatory apparatus absent. Trichobothria: tarsi I–IV (males and females): dorsally, two parallel rows of non-plumose clavate trichobothria ( Figure 4c,d View Figure 4 ) interspersed with filiform ( Figure 4e, f View Figure 4 ), separated by a row of long, thin setae. On metatarsi and tibia only filiform trichobothria. All tarsi without pseudosegmentation ( Figure 4a, b View Figure 4 ). Males and females, scopulae: ventrally, all tarsi densely scopulate and divided by conical long setae increasing in number from tarsus I to IV ( Figure 5a, b View Figure 5 ). Metatarsi I–III scopulate, entire or divided by conical long setae; metatarsus IV with no scopulae, almost inconspicuous ( Figure 6c View Figure 6 ) or welldeveloped apical scopulae (only in H. kuka sp. nov.) ( Figure 27f View Figure 27 ). Field of spiniform setae on basal retrolateral coxal face of palp and legs I–IV ( Figure 6b View Figure 6 ). Female palpal claw without teeth. Superior tarsal claw of all tarsi with one row with few small teeth. Third claw absent on all tarsi. Claw tufts well-developed on all tarsi ( Figure 4b View Figure 4 ). Four spinnerets ( Figure 5g View Figure 5 ): posterior lateral spinnerets three-segmented with digitiform apical segment ( Figure 5g, h View Figure 5 ); posterior median spinnerets entire ( Figure 5f, g View Figure 5 ). Tibia I of males usually short (shorter than metatarsus I) ( Figure 7g View Figure 7 ) with tibial apical apophysis originated from a common base, protruded and formed by two branches (prolateral and retrolateral) ( Figure 7f View Figure 7 , 10f View Figure 10 , 13g View Figure 13 , 19e View Figure 19 , 23g View Figure 23 , 30f View Figure 30 ), which are subequal or retrolateral slightly larger (except in H. martinorum with a noticeably larger retrolateral branch, Figure 13f, g View Figure 13 ). Prolateral branch with a basal ventral strong spine, and retrolateral with a shorter spine on inner face (except in H. chasqui sp. nov. bearing two spines). Metatarsus I curved and close outside the tibial apophysis branches ( H. albipes , H. chasqui sp. nov. and H. martinorum ) or between the two branches ( H. chespiritoi sp. nov., H. major and H. perezmilesi sp. nov.). Male with palpal tibia with a retrolateral rounded process ( Figure 7h, i View Figure 7 ). Palpal bulb globose at base with a large extended subtegulum. A basal digitiform apophysis well-developed ( H. albipes , H. major , H. martinorum and H. chespiritoi sp. nov.) ( Figure 8b View Figure 8 ) or absent ( H. chasqui sp. nov. and H. perezmilesi sp. nov.) ( Figure 20a View Figure 20 ) on male palpal bulb. Embolus thick, subcylindrical, strongly curved to the retrolateral side, bearing two prolateral keels (superior and inferior) with a slight development ( Figure 31a–d View Figure 31 ) or well-developed and strong ( Figure 8a–d View Figure 8 ). The prolateral inferior keel presents a ventral crest blunt ( Figure 31a, b View Figure 31 ) or pointed ( Figure 24a, b View Figure 24 ). Genitalia of females consists of one oval ( Figure 29e View Figure 29 ) or oblong ( Figure 33e View Figure 33 ) receptacle with an apical median region domed ( Figure 29e View Figure 29 ), straight ( Figure 33e View Figure 33 ) or depressed ( Figure 9j View Figure 9 ) and apical lateral projections varying in a more lateral ( Figure 26e View Figure 26 ) or apical location ( Figure 22e View Figure 22 ) and in development ( Figure 6e View Figure 6 ).
























No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Hapalotremus
| Ferretti, Nelson, Cavallo, Patricio, Chaparro, Juan C., Ríos-Tamayo, Duniesky, Seimon, Tracie A. & West, Rick 2018 |
Cyclothoracoides
| Raven RJ 1985: 151 |
| Strand E 1929: 17 |
Dolichothele Mello-Leitão, 1923: 120
| Raven RJ 1985: 152 |
| Mello-Leitao CF 1923: 120 |
Goniodontium Mello-Leitão, 1923: 127
| Raven RJ 1985: 153 |
| Mello-Leitao CF 1923: 127 |