
Anguillosyllis palpata ( Hartman, 1967 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4793.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:550F8461-03F6-4301-8791-605775D77467 |
|
persistent identifier |
https://treatment.plazi.org/id/FF3487CD-B756-BF34-91AB-FBFD6B13FDC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Anguillosyllis palpata ( Hartman, 1967 ) |
| status |
|
Anguillosyllis palpata ( Hartman, 1967) View in CoL
Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
Braniella palpata Hartman, 1967: 55 View in CoL , pl.14; Blake et al. 2009: 1797.
Braniella View in CoL nr. palpata: Blake & Grassle 1994: 850 View in CoL , 857; Hilbig 1994: 944; Hilbig & Blake 2006: 269.
Anguillosyllis palpata View in CoL — Aguado & San Martín 2008: 38; Barroso et al. 2017: 422–424 View Cited Treatment , Fig. 12 View FIGURE 12 .
Material examined. Antarctica. Drake Passage. Type material: 18 Sep 1963, USNS Eltanin, Station 740, 56°06– 56°07’S to 66°19’– 66°30’W, Blake trawl, 384–494 m, coll. H.L. Sanders, det. O. Hartman, holotype, USNM 55505; off GoogleMaps Cape Horn. 4 Nov 1962, USNS Eltanin, Station 315, 57°11’– 57°15ʹS to 66°19’– 66°30ʹW, Menzies trawl, 3770–3806 m, coll. H.L. Sanders, det. O. Hartman, paratype USNM 55506 GoogleMaps .
Additional material: Antarctica. (11 specimens from 11 stations) ANDEEP I, coll. J.A. Blake. Drake Passage, Sta. PS 61/042-6 , 28 Jan 2002, 59°40.42ʹS, 7°35.75ʹW, 3692 m, large box corer, 1 specimen ( SMF 24864); Sta. PS61/046-5 GoogleMaps , 30 Jan 2002, 60°38.13ʹS, 53°57.68ʹW, 2894 m, large box corer, 1 specimen ( SMF 24865); — Scotia Sea, Sta. PS61/105-2 GoogleMaps , 14 Feb 2002, 61°24.14ʹS, 58°51,15ʹW, 2290 m, multicorer, 1 specimen ( SMF 24866); Sta. PS61/114-9 GoogleMaps , 19 Feb 2002 61°43.58ʹS, 60°43.22ʹW, 2876 m, multicorer, 1 specimen ( SMF 24867). ANDEEP II, Weddell Sea, Sta. PS61/131-8 GoogleMaps , 6 Mar 2002, 65°18.65ʹS, 51° 30.91ʹW, 3068 m, large box corer, 1 specimen ( SMF 24868); Sta. PS61/132-4 GoogleMaps , 6 Mar 2002, 65°17.75ʹS, 53° 22.92ʹW, 2085 m, large box corer, 1 specimen ( SMF 24869); Sta. PS61/132-5 GoogleMaps , 7 Mar 2002, 65°17.68ʹS, 53° 23.00ʹW, 2084 m, multicorer, 1 specimen ( SMF 24870); Sta. PS61/139-7 GoogleMaps , 20 Mar 2002, 58°14.10ʹS, 24° 20.73ʹW, 3935 m, large box corer, 1 specimen ( SMF 24871). ANDEEP III, coll. S. Doner. Weddell Sea, Sta. PS 67/059-4 GoogleMaps , 14 Feb 2005, 67°30,99ʹS, 0°0,16ʹE, 4652 m, multicorer, 1 speci- men ( SMF 24872).— Weddell Sea, off Cape Norvegia, Sta. PS67/074-5 GoogleMaps , 20 Feb 2005, 71°18.11ʹS, 13°56.33ʹW, 1035 m, multicorer, 1 specimen ( SMF 24873).— Southern Ocean, Powell Basin, Sta. PS67/121-10 GoogleMaps , 14 Mar 2005, 63°41.74ʹS, 50°42.99ʹW, 2621 m, large box corer, 1 specimen ( SMF 24874).— Gulf of Mexico , off Louisiana. (207 specimens from 20 stations) GoogleMaps . Matterhorn Lateral Pipeline Survey , coll. J.A. Blake, Chief Scientist, R / V Brooks Mc- Call , 10 July 2008, Sta. 5, 28°50.2021ʹN, 89°02.1818ʹW, 241 m, 99.5% silt-clay sediment, boxcorer, 15 specimens ( MCZ 150563 View Materials ). Matterhorn Platform Survey , coll. J.A. Blake, Chief Scientist GoogleMaps , 10 Nov 2008, boxcorer: Sta. 1N , 28°44.7163′N, 88°49.5337ʹW, 850m, 97.5%silt-clay sediment, 17 specimens ( MCZ150564 View Materials ); Sta. 2N GoogleMaps , 28°44.7798′N, 88°49.5334′W, 825 m, 98.5% silt-clay, 15 specimens ( MCZ 150565 View Materials ); Sta. 3N GoogleMaps , 28°44.8362ʹN, 88°49.5336′W, 840 m, 97.4% silt-clay, 15 specimens ( MCZ 150566 View Materials ); Sta. 4N GoogleMaps , 28°45.1034′N, 88°49.5331′W, 821 m, 99% silt-clay, 20 specimens ( MCZ 150567 View Materials ). Sta. 1S GoogleMaps , 28°44.3406′N, 088°49.5378′W, 875 m, 97.8% silt-clay, 11 specimens ( MCZ 150568 View Materials ); Sta. 2S GoogleMaps , 28°44.2796′N, 88°49.5347′W, 878 m, 98.3% silt-clay, 5 specimens ( MCZ 150569 View Materials ); Sta. 3S GoogleMaps , 28°44.2246′N, 88°49.5391′W, 884 m, 98.7% silt-clay, 19 specimens ( MCZ 150570 View Materials ); Sta. 4S GoogleMaps , 28°43.9584′N, 88°49.5465′W, 908 m, 98.7% silt-clay, 1 specimen ( MCZ 150571 View Materials ); Sta. 5S GoogleMaps , 28°43.4126′N, 88°49.5373′W, 955 m, 98.1% silt-clay, 1 specimen ( MCZ 150572 View Materials ). Sta. 1E GoogleMaps , 28°44.5352′N, 88°49.3039′W, 867 m, 98.6% silt-clay, 6 spec- imens ( MCZ 150573 View Materials ); Sta. 3E GoogleMaps , 28°44.5244′N, 88°49.2116′W, 868 m, 98.8% silt-clay, 5 specimens ( MCZ 150574 View Materials ); Sta. 4E GoogleMaps , 28°44.5232′N, 88°48.8942′W, 891 m, 98.6% silt-clay, 8 specimens ( MCZ 150575 View Materials ). Sta. 1W GoogleMaps , 28° 44.52630′N, 88°49.7627′W, 851 m, 97.4% silt-clay, 8 specimens ( MCZ 150576 View Materials ); Sta. 2W GoogleMaps , 28°44.5327′N, 88°49.8318′W, 853 m, 97.8% silt-clay, 10 specimens ( MCZ 150577 View Materials ); Sta. 3W GoogleMaps , 28°44.5279′N, 88°49.8851′W, 850 m, 98.5% silt-clay, 21 specimens ( MCZ 150578 View Materials ); Sta. 4W GoogleMaps , 28°44.5305′N, 88°50.2023′W, 839 m, 99.2% silt-clay, 8 specimens ( MCZ 150579 View Materials ); Sta. 5W GoogleMaps , 28°44.5332′N, 88° 50.8139′W, 818 m, 98.8% silt-clay, 15 specimens ( MCZ 150580 View Materials ). Matterhorn Lateral Gas Export Pipeline Survey, coll. S. Aubrey, Chief Scientist, R /V Ocean Intervention GoogleMaps , 4 June 2009, Sta. 8C , 28°51.797′N, 088°59.055′W, 241 m, 99.5% silt-clay, 6 specimens ( MCZ 150587 View Materials ); Sta. 9C GoogleMaps , 28°53.821′N, 088°56.766′W, 229 m, 99.5% silt-clay, 1 specimen ( MCZ 150588 View Materials ).— Western North Atlantic continental slope and rise. (196 specimens in 27 samples from 7 stations). off Delaware GoogleMaps , USA. US Mid Atlantic ACSAR Program , coll. R. Petrecca ( WHOI), Chief Scientist, Sta. Mid 4, Cruise M4, R/V Oceanus , 16 May 1985, Rep. 1, 38°44.44ʹN, 72°41.24ʹW, 2100 m, 1 specimen ( USNM 1480243 View Materials ).— North Carolina GoogleMaps , USA, off Cape Lookout, U.S. South AC- SAR Program , coll. J.A. Blake (Battelle), Chief Scientist. Sta. South 1, Cruise SA 2, R/ V Cape Hatteras , 27 Mar 1984, Rep. 2 , 34°15.81ʹN, 75°46.01ʹW, 583 m, 1 specimen ( USNM 1480217 View Materials ); Rep. 3 GoogleMaps , 34°15.81ʹN, 75°45.78ʹW, 593 m, 2 specimens ( USNM 1480218 View Materials ); Cruise SA3, R/V Gyre GoogleMaps , 15 July 1984, Rep. 1, 34°16.4ʹN, 75°45.8ʹW, 584 m, 2 specimens ( USNM 1480219 View Materials ). Sta. South 2, Cruise SA 1, R/ V Columbus Iselin GoogleMaps , 12–13 Nov 1983, Rep. 1 , 34°14.87ʹN, 75°43.79ʹW, 1013 m, 22 specimens ( USNM 1480220 View Materials ); Rep. 2 GoogleMaps , 34°14.23ʹN, 75°43.89ʹW, 1000 m, 4 specimens ( USNM 1480221 View Materials ); Rep. 3 GoogleMaps , 34°14.07ʹN, 75°44.06ʹW, 995 m, 7 specimens ( USNM 1480222 View Materials ). Cruise SA2, R/V Cape Hatteras GoogleMaps , 27 March 1984, Rep. 1 , 34°15.04ʹN, 75°43.66ʹW, 1019 m, 25 specimens ( USNM 1480223 View Materials ); Rep. 2 GoogleMaps , 34°14.46ʹN, 75°43.85ʹW, 1003 m, 2 specimens ( USNM 1480224 View Materials ); Rep. 3 GoogleMaps , 34°14.56ʹN, 75°43.35ʹW, 1000 m, 20 specimens ( USNM 1480225 View Materials ). Cruise SA3, R/V Gyre GoogleMaps , 15 July 1984, Rep. 1 , 34°14.5ʹN, 75°43.9ʹW, 984 m, 26 specimens ( USNM 1480226 View Materials ); Rep. 2 GoogleMaps , 34°15.0ʹN, 75°43.7ʹW, 1002 m, 25 specimens ( USNM 1480227 View Materials ); Rep. 3 GoogleMaps , 34°15.2ʹN, 75°43.6ʹW, 1007 m, 8 specimens ( USNM 1480228 View Materials ).— North Carolina GoogleMaps , USA, Off Cape Fear. Sta. South 11, Cruise SA 5, R/ V Gyre , 23 Sep 1985, Rep. 1 , 33°04.83ʹN, 76°25.19ʹW, 796 m, 2 specimens ( USNM 1480229 View Materials ); Rep. 2 GoogleMaps , 33°04.83ʹN, 76°25.17ʹW, 896 m, 2 specimens ( USNM 1480230 View Materials ); Rep. 3 GoogleMaps , 33°04.86ʹN, 76°25.12ʹW, 797 m, 1 specimen ( USNM 1480231 View Materials ); Cruise SA6, R/V Cape Hatteras GoogleMaps , 22 Nov 1985, Rep. 3, 33°04.84ʹN, 76°25.06ʹW, 807 m, 1 specimen ( USNM 1480232 View Materials ). Sta. South 12, Cruise SA 6, R/ V Cape Hatteras GoogleMaps , 21 Nov 1985, Rep. 1, 33°00.55ʹN, 76°07.45ʹW, 1992 m, very soft mud, 1 specimen ( USNM 1480233 View Materials ).— South Carolina GoogleMaps , USA, northern Blake Plateau , off Charleston, SC. Sta. South 14, Cruise SA 4, R/V Cape Hatteras , 20 May 1985, Rep. 1 , 32°23.64ʹN, 77°01.13ʹW, 805 m, 6 specimens ( USNM 1480234 View Materials ); Rep. 2 GoogleMaps , 32°23.64’N, 77°01.19’W, 802 m, 8 specimens ( USNM 1480235 View Materials ); Rep. 3 GoogleMaps , 32°23.67ʹN, 77°01.12ʹW, 803 m, 2 specimens ( USNM 1480236 View Materials ). Cruise SA5, R/V Gyre GoogleMaps , 19 Sep 1985, Rep. 1, 32°23.67ʹN, 77°01.18ʹW, 796 m, pteropod ooze/green mud, 2 specimens ( USNM 1480237 View Materials ) GoogleMaps ; 20 Sep 1985, Rep. 3, 32°23.63ʹN, 77°01.11ʹW, 799 m, 10 specimens ( USNM 1480238 View Materials ). Cruise SA 6, R/ V Cape Hatteras GoogleMaps , 18 Nov 1985, Rep. 1 , 32°23.73ʹN, 77°01.10ʹW, 799 m, 7 specimens ( USNM 1480239 View Materials ); Rep. 2 GoogleMaps , 32°23.67ʹN, 77°01.09ʹW, 799 m, 3 specimens ( USNM 1480240 View Materials ); Rep. 3 GoogleMaps , 32°23.70ʹN, 77°01.06ʹW, 799 m, 5 speci- mens ( USNM 1480241 View Materials ). Sta. South 16, Cruise SA6, R/V Cape Hatteras GoogleMaps , 20 Nov 1985, Rep. 3, 31°35.16ʹN, 75°10.22ʹW, 3012 m, 1 specimen ( USNM 1480242 View Materials ).— Gulf of the Farallones , California GoogleMaps continental slope. (137 specimens in 65 samples). US Navy, 103 Site Selection Survey, west of the Farallon Islands , coll. J.A. Blake. Sta. B2 , 19 Jul 1991, 37°40.36ʹN, 123°27.96′W, 2701 m, 1 specimen ( CASIZ 230506 ); Sta. B3 GoogleMaps , 20 Jul 1991, 37°40.06′N, 123°23.69′W, 2385 m, 2 specimens ( CASIZ 230505 ); Sta. B8 GoogleMaps , 24 Jul 1991, 37°38.58′N, 123°29.86′W, 2985 m, 2 specimens ( CASIZ 230504 ).— EPA 102 Site Survey, off San Francisco, north of Pioneer Canyon , R/V Point Sur , coll. J.A. Blake. Sta. 3-8 GoogleMaps , 13 Sep 1991, 37°27.13′N, 123°23.02′W, 2005 m, 1 specimen (NJM); Sta. 3-13, 15 Sep 1991, 37°24.30′N, 123°20.02′W, 1780 m, 2 specimens (LACM-Poly 10477); Sta. 3-18, 15 Sep 1991, 37°22.31′N, 123°19.24′W, 1990 m, 3 specimens ( CASIZ 230518 ); Sta. 3-19 GoogleMaps , 16 Sep 1991, 37°26.05′N, 123°19.99′W, 1760 m, 1 specimen ( CASIZ 230519 ); south of Pioneer Canyon Sta. 4-15 GoogleMaps , 19 Sep 1991, 37°12.42′N, 123°15.81′W, 1730 m, 1 specimen (LACM-Poly 10478).— San Francisco Deep Ocean Disposal Site (SF-DODS), west of the Farallon Islands , coll. JA Blake. Sta. 6 GoogleMaps , 16 Oct 2000, 37°39.97′N, 123°26.95′W, 2730 m, 1 specimen ( CASIZ 230502 ) GoogleMaps ; 22 Sep 2002, 37°40.02′N, 123°27.00′W, 2697, 1 specimen ( CASIZ 230503 ); Sta. 10 GoogleMaps , 8 Oct 1999, 37°39.96′N, 123°31.06′W, 3015 m, 3 specimens (LACM-AHF Poly 10479); 23 Oct 2001, 37°40.02′N, 123°31.00′W, 2985 m, 1 specimen (LACM-AHF Poly 10480); 22 Sep 2002, 37°40.00′N, 123°31.00ʹW, 2739 m, 4 specimens ( CASIZ 230490 ) GoogleMaps ; 14 Sep 2005, 37°39.970′N, 123°31.997′W, 2750 m, 1 specimen ( CASIZ 230491 ) GoogleMaps ; 26 Sep 2006, 37°40.197′N, 123°30.834′W, 2760 m, 1 specimen ( CASIZ 230489 ). Sta. 11 GoogleMaps , 11 Dec 1996, 37°39.09′N, 123° 30.99′W, 2990, 1 specimen (NJM); 3 Nov 1997, 37°39.10′ N, 123° 31.03′ W, 2995, 2 specimens ( CASIZ 230511 ). Sta. 12 GoogleMaps , 5 Dec 1996, 37°38.92′ N, 123° 29.85′ N, 3097 m, 3 specimens ( CASIZ 230510 ). Sta. 16 , 11 Jul 2007, 37°37.981′ N, 123°26.957′ W, 2699 m, 2 specimens (LACM-AHF Poly 10481). Sta. 17 GoogleMaps , 7 Oct 1998, 37°38.00′N, 123°27.99′W, 2830 m, 1 specimen (CA- SIZ 230500); 13 Oct 1999, 37°38.02′N, 123°20.00′W, 2770 m, 1 specimen ( CASIZ 230501 ) GoogleMaps ; 21 Oct 2001, 37°38.00′N, 123°28.03′W, 2862 m, 2 specimens ( CASIZ 230499 ) GoogleMaps ; 21 Sep 2002, 37°38.01′N, 123° 27.94′W, 2775 m, 6 specimens (LACM-AHF Poly 10482); 15 Sep 2005, 37°37.998′N, 123° 27.995′W, 2750 m, 1 specimen ( LACM- AHF Poly 10483). Sta. 18 GoogleMaps , 30 Oct 1997, 37°37.95′N, 123°28.89′W, 2905 m, 1 specimen ( CASIZ 230520 ) GoogleMaps ; 8 Oct 1999, 37°37.86′N, 123°28.86′W, 2900m, 1specimen ( CASIZ 230521 ). Sta. 19 GoogleMaps , 7Oct 1998, 37°38.04′N, 123°30.02′W, 3030 m, 4 specimens (LACM-AHF Poly 10484); 15 Oct 2000, 37°37.95′N, 123°29.95′W, 2940 m, 2 specimens ( CASIZ 230483 ) GoogleMaps ; 22 Oct 2001, 37°37.98′N, 123°30.03′W, 3123 m, 2 specimens ( CASIZ 230484 ) GoogleMaps ; 22 Sep 2002, 37°38.01′N, 123°30.00′W, 3000 m, 3 specimens ( CASIZ 230482 ) GoogleMaps ; 25 Sep 2006, 37°37.975′N, 123°30.013′W, 3083 m, 2 specimens ( CASIZ 230481 ) GoogleMaps ; 11 Jul 2007, 37°38.166′N, 123°30.213′W, 3100 m, 2 specimens (LACM-AHF Poly 10485). Sta. 20 GoogleMaps , 15 Oct 2000, 37°37.96′N, 123°31.06′W, 3060 m, 2 specimens (LACM-AHF Poly 10486); 23 Oct 2001, 37°38.04′N, 123°31.01′W, 3142 m, 1 specimen (LACM-AHF Poly 10487); 22 Sep 2002, 37°38.06′N, 123°31.01′W, 3050 m, 4 specimens ( CASIZ 230496 ) GoogleMaps ; 15 Sep 2005, 37°37.987′N, 123°30.989′W, 3037 m, 4 speci- mens ( CASIZ 230497 ) GoogleMaps ; 25 Sep 2006, 37°37.960′N, 123°30.996′W, 3052 m, 1 specimen ( CASIZ 230495 ) GoogleMaps ; 11 Jul 2007, 37°38.163′N, 123°31.265′W, 2980 m, 1 specimen ( CASIZ 230498 ). Sta. 23 GoogleMaps , 10 Dec 1996, 37°37.13′N, 123°29.08′W, ~ 2950 m, 2 specimens ( CASIZ 230514 ) GoogleMaps ; 3 Nov 1997, 37°37.06′N, 123°29.15′W, ~ 2950 m, 3 speci- mens ( CASIZ 230513 ) GoogleMaps ; 6 Oct 1998, 37°37.23′N, 123°29.18′W, 2970 m, 1 specimen ( CASIZ 230517 ) GoogleMaps ; 9 Oct 1999, 37°36.96′N, 123°28.93′W, 2970 m, 6 specimens ( CASIZ 230516 ) GoogleMaps ; 26 Sep 2002, 37°36.95′N, 123°29.02′W, 2954 m, 1 specimen with possible dorsal egg mass ( CASIZ 230515 ) GoogleMaps ; 24 Sep 2003, 37°36.86′N, 123°28.85′W, 2821 m, 5 specimens (LACM-AHF Poly 10488); 25 Sep 2006, 37°36.984′N, 123°28.928′W, 2929 m, 1 specimen ( LACM- AHF Poly 10489) GoogleMaps ; 13 Jul 2007, 37°37.170′N, 123°29.222′W, 2995 m, 1 specimen (LACM-AHF Poly 10490). Sta. 24 GoogleMaps , 19 Oct 2000, 37°36.97′N, 123°27.92′W, 2650 m, 1 specimen ( CASIZ 230479 ) GoogleMaps ; 22 Oct 2001, 37°36.94′N, 123°27.93′W, 2680 m, 4 specimens ( CASIZ 230480 ). Sta. 27 GoogleMaps , 16 Oct 2000, 37°40.97′N, 123°31.98′W, 2740 m, 3 specimens (LACM-AHF Poly 10491); 22 Oct 2001, 37°40.98′N, 123°31.95′W, 2770 m, 1 specimen ( CASIZ 230508 ) GoogleMaps ; 23 Sep 2002, 37°40.96′N, 123°31.94′W, 2825 m, 4 specimens ( CASIZ 230507 ) GoogleMaps ; 11 Jul 2007, 37°41.141′N, 123°32.078′W, 2832 m, 1 specimen ( CASIZ 230509 ). Sta. 57 GoogleMaps , 22 Oct 2001, 37°42.96′N, 123°32.97′W, 2750 m, 1 specimen ( CASIZ 230485 ). Sta. 64 GoogleMaps , 25 Sep 2002, 37°36.01′N, 123°33.01′W, 3136 m, 7 specimens ( CASIZ 230494 ) GoogleMaps ; 14 Sep 2005, 37°36.079′N, 123°32.954′W, 3120 m, 1 specimen ( CASIZ 230492 ) GoogleMaps ; 27 Sep 2006, 37°35.968′N, 123°32.989′W, 3145 m, 1 specimen ( CASIZ 230493 ) GoogleMaps ; 13 Jul 2007, 37°36.028′N, 123°33.078′W, 3115 m, 1 speci- men (LACM-AHF Poly 10492). Sta. 92 GoogleMaps , 24 Sep 2006, 37°45.080′N, 123°35.134′W, 2815 m, 1 specimen ( LACM- AHF Poly 10493). Sta. 116 GoogleMaps , 25 Sep 2002, 37°34.99′N, 123°28.98′W, 2975 m, 4 specimens ( CASIZ 230488 ; 14 Sep 2005, 37°34.927′N, 123°28.935′W, 2733 m, 2 specimens ( GoogleMaps CASIZ 230486 ; 27 Sep 2006, 37°35.051′N, 123°29.034′W, 3052 m, 3 specimens ( GoogleMaps CASIZ 230487 ); 13 Jul 2007, 37°35.080′N, 123°29.039′W, 2908 m, 3 specimens ( GoogleMaps LACM- AHF Poly 10494). Sta. DR 2A, 29 Sep 2006, 37°22.924′N, 124°01.001′W, 3775 m, 3 specimens (LACM-AHF Poly 10495) GoogleMaps . Sta. DR 2B, 29 Sep 2006, 37°22.924′N, 124°01.001ʹW, 3775 m, 2 specimens ( GoogleMaps CASIZ 230512 ). Sta. DR 3B, 13 Jul 2007, 37°17.537′N, 124°09.192′W, 3864 m, 1 specimen (LACM-AHF Poly 10496). GoogleMaps — Abyssal Pacific, Clarion-Clipperton Fracture Zone. (2 specimens from 2 samples). NOAA DOMES Site C , coll. R.R. Hessler, ECHO I, sample H352, Sandia box corer, 14 June 1983, 14 o 38.21'N, 125 o 26.32'W, 4502 m, 1 specimen ( LACM-AHF Poly 10172 ) GoogleMaps ; sample H353, Sandia box corer, 18 June 1983, 14 o 42.09'N, 125 o 24.27'W, 4516 m, 1 juvenile ( LACM-AHF Poly 10173 ). GoogleMaps — South China Sea, off Brunei. (38 specimens from 20 stations). coll. J.A. Blake, Chief Scientist. Sta . 6, 7Jun2011, 5°51′40.17938″N, 114°10′54.32730″E, 1353 m, 1juvenile ( NJM) GoogleMaps ; Sta.9 , 6June 2011, 5°52′38.23770″N, 114°13′42.28254″E, 1459m, 3specimens ( MCZ148547 View Materials ) GoogleMaps ; Sta.11 , 6June2011, 5°50′27.84205″N, 114°10′56.95330″E, 1405 m, 5 specimens ( MCZ 148548 View Materials ) GoogleMaps ; Sta. 23 , 5 June 2011, 5°50′24.47069″N, 114°14′04.40344″E, 1411 m, 1 specimen ( MCZ 148549 View Materials ) GoogleMaps ; Sta. 27 , 5 June 2011, 5°46′11.81146″N, 114°09′16.72096″E, 1510 m, 3 specimens ( MCZ 148550 View Materials ) GoogleMaps ; Sta. 28 , 4 June 2011, 5°44′29.74715″N, 114°07′42.47198″E, 1557 m, 1 specimen ( MCZ 148551 View Materials ) GoogleMaps ; Sta. 29 , 4 Jun 2011, 5°43′01.15997″N, 114°06′07.18190″E, 1537 m, 1 specimen ( MCZ 148552 View Materials ) GoogleMaps ; Sta. 34 , 4 June 2011, 5°50′32.29819″N, 114°15′47.44656″E, 1369 m, 3 specimens, poor condition ( NJM) GoogleMaps ; Sta. 38 , 3 June 2011, 5°42′59.40134″N, 114°09′16.14114″E, 1353 m, 1 specimen ( MCZ 148553 View Materials ) GoogleMaps ; Sta. WH-Jokit-NE , 6 Jun 2011, 5°53′28.38401″N, 114°11′57.24436″E, 1400 m, 4 specimens ( MCZ 148554 View Materials ) GoogleMaps ; Sta. WH-Jokit-SW , 5 Jun 2011, 5°46′27.76503″N, 114°07′33.98027″E, 1487 m, 2 specimens ( MCZ 148555 View Materials ) GoogleMaps . Sta. WH-Jaring-SWB , 4 June 2011, 5°43′13.87265″N, 114°08′13.15762″E, 1463 m, 3 specimens ( MCZ 148556 View Materials ) GoogleMaps .— coll. P. Neubert, Chief Scientist. Sta. AN5 , 27 Jun 2011, 5°26′51.39448″N, 113°46′41.57612″E, 1632 m, 2 specimens ( MCZ 148557 View Materials ) GoogleMaps ; Sta. ME7 , 4 July 2011, 5°22′08.21298″N, 113°37′25.97978″E, 1725 m, 1 specimen ( MCZ 148558 View Materials ) GoogleMaps ; Sta. SA6 , 29 June 2011, 5°36′19.08413″N, 113°37′45.57822″E, 2178m, 1specimen ( MCZ148559 View Materials ) GoogleMaps ; Sta.TU2 , 24June2011, 5°18′25.22812″N, 113°48′13.59077″E, 1224 m, 1 specimen ( MCZ 148560 View Materials ) GoogleMaps ; Sta. TU4 , 25 Jun 2011, 5°12′34.01837″N, 113°42′17.60077″E, 1439m, 1specimen ( MCZ148561 View Materials ) GoogleMaps ; Sta.TU7 , 4Jul2011, 5°19′24.21956″N, 113°47′15.61845″E, 1327 m, 2 specimens ( MCZ 150560 View Materials ) GoogleMaps ; Sta. TU8 , 24 June 2011, 5°21′35.85828″N, 113°49′14.10977″E, 1325 m, 1 specimen ( MCZ 150561 View Materials ) GoogleMaps ; Sta. TU14 , 23 June 2011, 5°22′23.91273″N, 113°48′25.59116″E, 1444 m, 1 specimen ( MCZ 150562 View Materials ). GoogleMaps
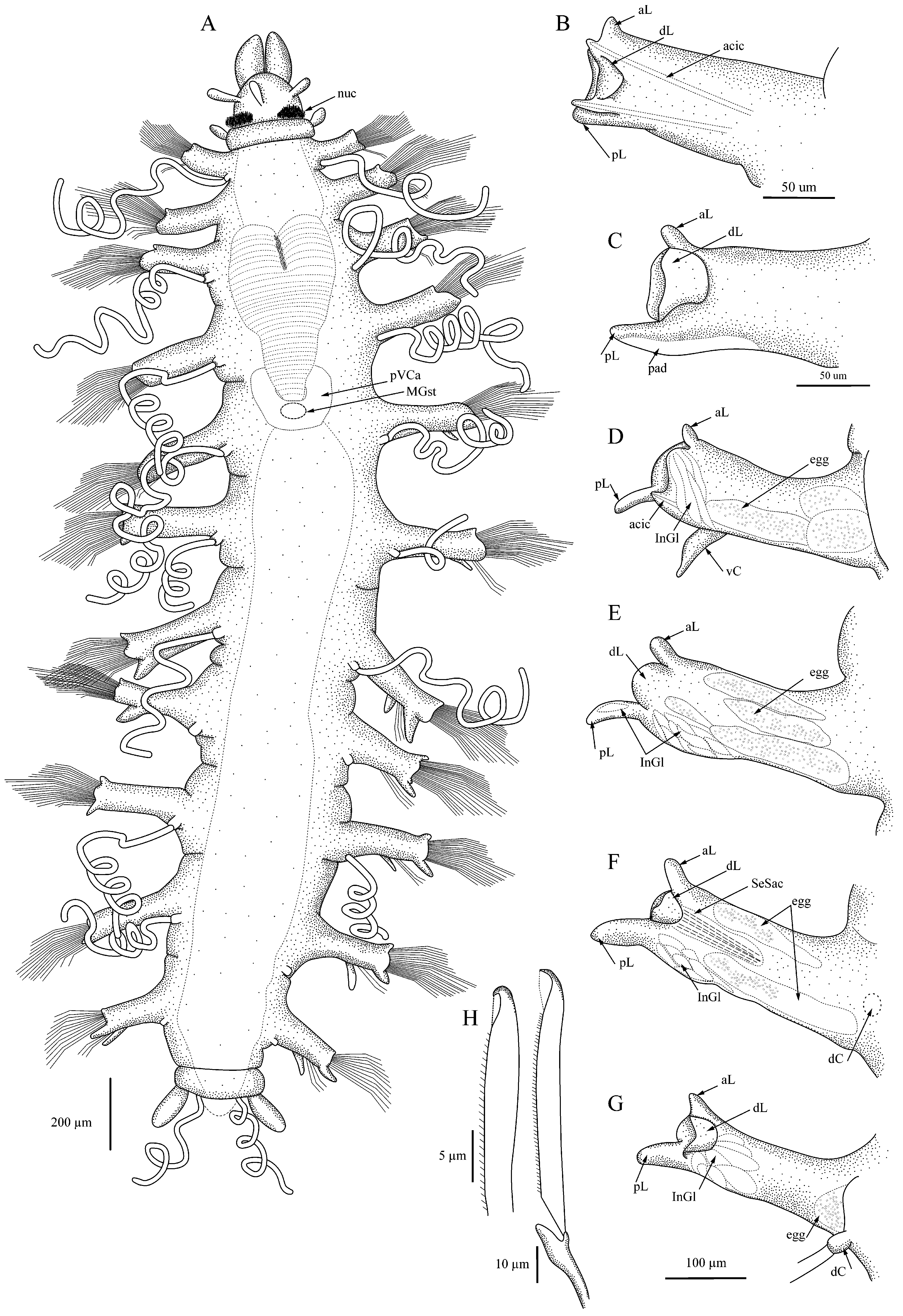
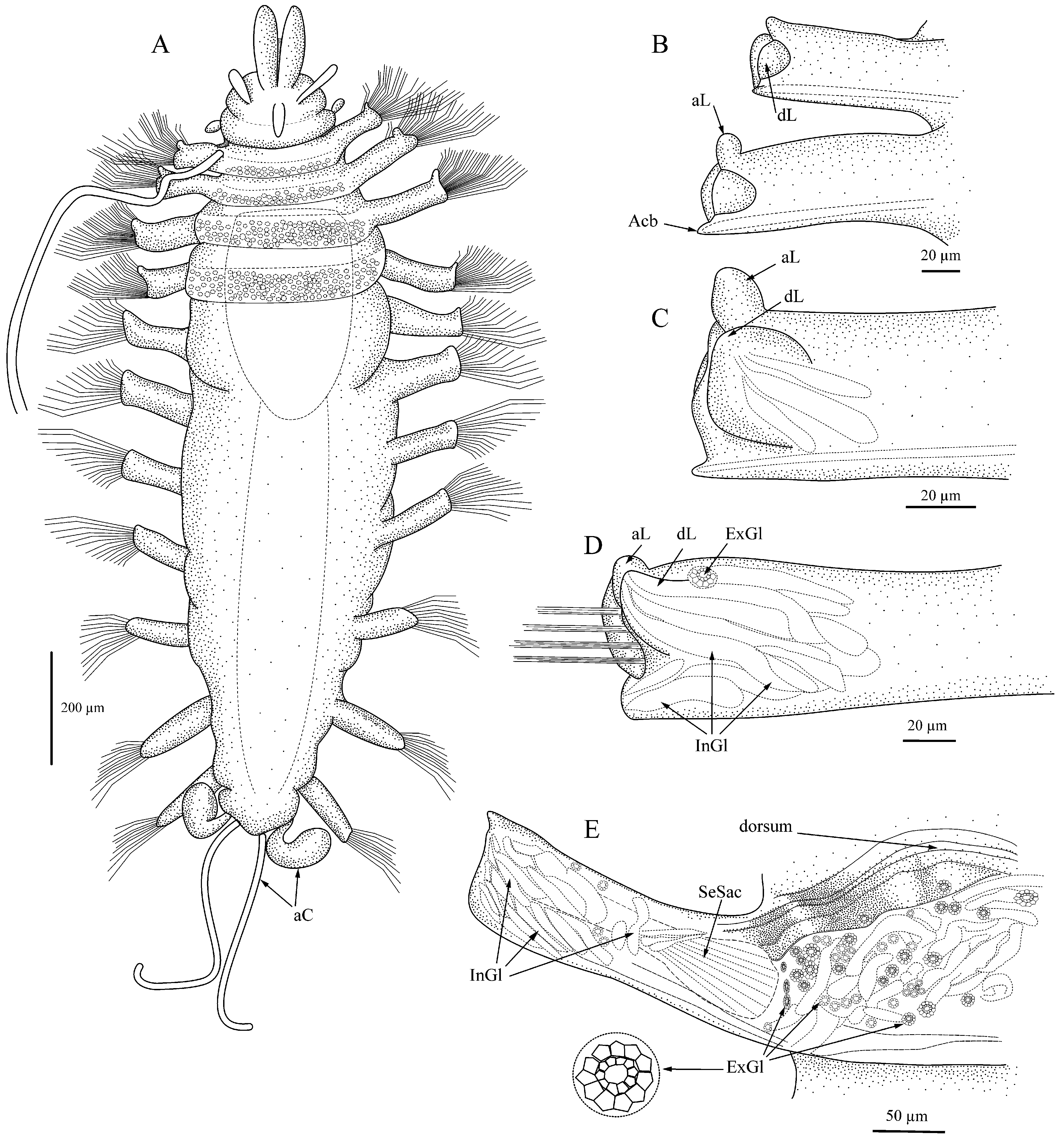
Description. Body with 11 setigers, linear in overall shape ( Figs. 1A View FIGURE 1 , 3A View FIGURE 3 ), maximal length 2.8 mm without anal cirri, most specimens 1.0 mm long; maximal width 1.0 mm without setae; juveniles with 9–10 setigers 0.5 mm long or less.
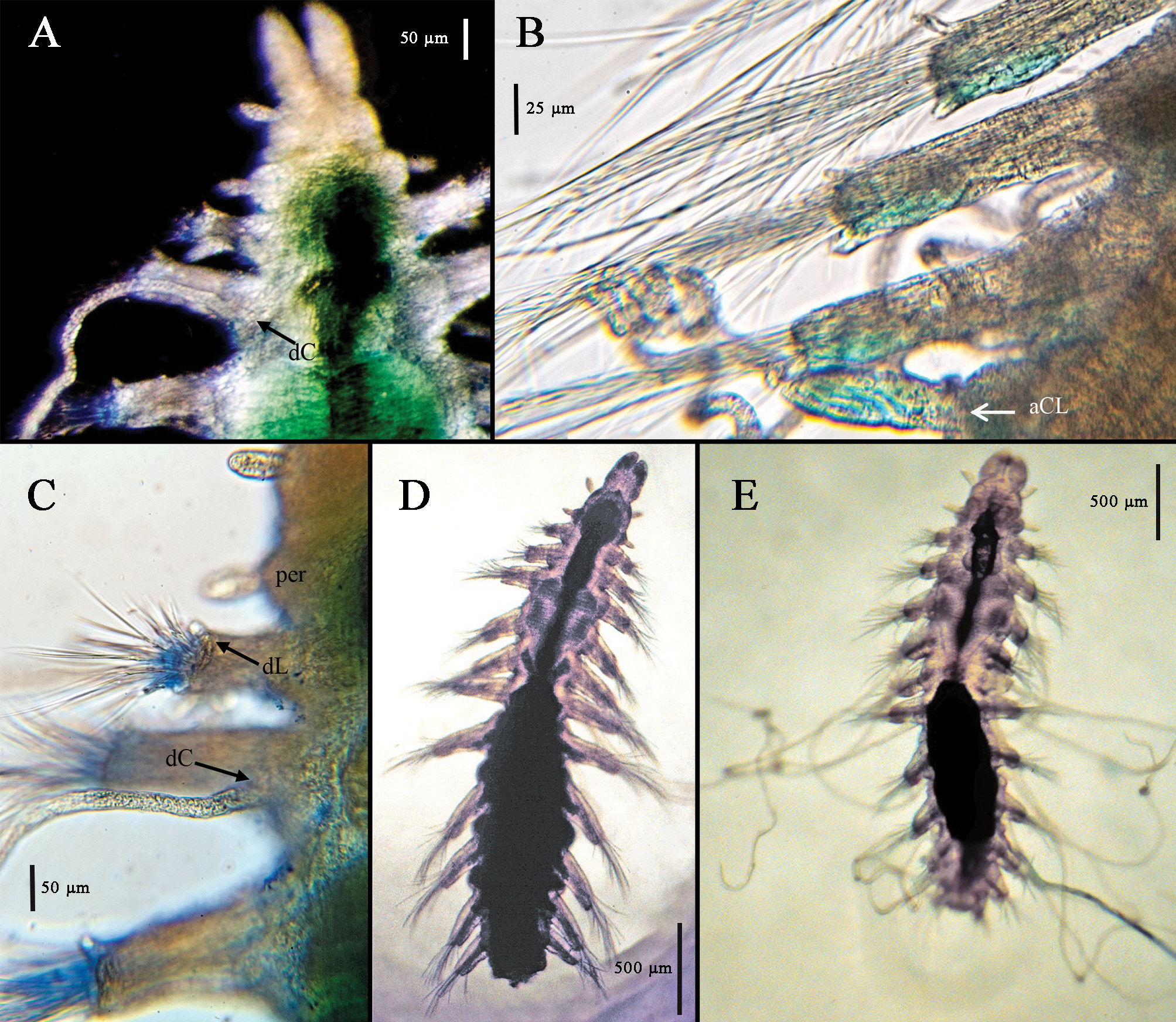
Palps elongated bean shape ( Figs. 1A View FIGURE 1 , 3A View FIGURE 3 , 4A View FIGURE 4 ), free to base but often appearing closely applied on proximal half, dorsally with distinct furrow and separation, ventrally joined giving fused appearance at low magnifications; prostomium longer than wide, convex medially resulting in dome-shaped appearance, with three club-shaped antennae in nearly transverse row, two lateral antennae slightly anterior to medial one; eyes lacking; large oval nuchal pits between prostomium and peristomium filled with long cilia ( Figs. 1A View FIGURE 1 , 3B View FIGURE 3 ). Peristomium shorter than prostomium, with two tentacular cirri similar in shape to prostomial antennae, shorter than prostomial antennae but sometimes appearing inflated. Eversible pharynx with distal ring of 10 soft papillae; tooth absent. Proventricle in three setigers, distinctly heart-shaped with abruptly tapered posterior end ( Figs. 1A View FIGURE 1 , 3 View FIGURE 3 A–E); rows of muscle cells somewhat indistinct, perhaps 30–35; post-ventricle caeca wrapped around ventricle ( Figs.1A View FIGURE 1 , 3E View FIGURE 3 ), with dorsal circlet of cells retaining deep MB stain ( Fig. 3A, I View FIGURE 3 ).
Dorsum with first 2–3 setigers sometimes appearing slightly biannulate. Parapodia uniramous, elongated, rectangular, shortest on first and last setiger; with anterior and posterior lobes small on anterior setigers, becoming larger through posterior setigers ( Figs. 1 View FIGURE 1 B–G, 2A–I); posterior lobe larger than anterior lobe in middle and posterior setigers; large dorsal lobe or hood present on all setigers, rounded, largest on setigers 5–6, may be extended over setae or curled or folded back toward midline. Posterior margin of parapodial lobe with pad of cells that retain MB stain ( Figs. 1C View FIGURE 1 , 4B View FIGURE 4 ); elongate internal parapodial glands extending up into dorsal lobe ( Fig. 1 View FIGURE 1 E–G). Dorsal cirri with basal cirrophores, cirri extremely long, thin, measuring up to 2.5 times body length when gently stretched out or in life ( Fig. 4E View FIGURE 4 ), may be coiled into corkscrew or relaxed; cirri present on setiger 1, usually lacking on setiger 2 but sometimes present ( Fig. 4A, C View FIGURE 4 ), present on setigers 3–11. Ventral cirri short, widest at base, tapering toward distal end, inserted midway between body and distal end of parapodia on all setigers.
All setae compound, with heterogomph shafts; up to 20 long-bladed setae emerging from distal end of parapodium, additional group of up to 10–12 short-bladed falcigerous setae emerging in two rows from ventral face of parapodium between insertion of ventral cirrus and distal end ( Fig. 3H View FIGURE 3 ); fewer setae in posterior setigers. Setal shafts measuring 70–130 µm long, with shorter ones ventral, shafts often equal to blade length, never exceeding 3x length of blades. All setae serrated; falcigers 20–50 µm long, with small shoulder before hooked tip, tip dark, with small clear hood between tip and shoulder of blade ( Fig. 1H View FIGURE 1 , 3 View FIGURE 3 F–G). Longer, spiniger-like blades up to 200 µm long. Each parapodium with acicula in both anterior and posterior edges ( Fig. 1B View FIGURE 1 ); tip of posterior acicula sometimes appearing bent at nearly right angle in posterior setigers.
Pygidium with four anal cirri: two lateral, oval and two ventromedial, thin, filiform, often coiled ( Fig. 1A View FIGURE 1 ); cirri easily lost.
Reproductive characteristics. Several specimens from each location were found to have eggs in the coelom starting after the end of the proventricle in setiger 6 or 7 and continuing to the end of the body. The eggs measured up to 75 µm in diameter and were usually on either side of the midline, pushed up against the parapodia; in the material from California and the western North Atlantic, eggs were seen squeezing into the parapodia and in at least one specimen the eggs appeared to have dark inclusions ( Fig. 1 View FIGURE 1 D–G). One specimen (Sta. 23, 2002, CASIZ 230515) had an attached mass ( Fig. 3D View FIGURE 3 ) that appeared to contain at least one round object; whether this might be an external brood pouch is uncertain. Dorsal brooding was recently documented in A. lanai by Fukuda & Barroso (2019).
Methyl Green staining pattern. All specimens retained light MG stain on the pharyngeal papillae, the cells along the posteroventral edge of the parapodia, and the broad, oval anal cirri. Deep stain was retained by the area immediately anterior to the proventricle and a dorsal circlet of cells on the post-ventricle ( Fig. 3A View FIGURE 3 ).
Ecological Notes. Anguillosyllis palpata often co-occurred with other species of Anguillosyllis , usually a fusedpalp species. For example, in the collections from California, A. palpata was found with A. inornata n. sp. In some areas, notably ASCAR South Sta. 2, A. palpata occurred in numbers up to 20–26 in each 0.1-m 2 box-core sample, resulting in this species being among the dominant 20 species in the benthic community ( Blake & Grassle 1994, as Braniella nr. palpata ).
Remarks. Anguillosyllis palpata is one of the larger species of the genus, reaching 2.8 mm in length in the Antarctic material ( Hartman 1967, this study) and 4.3 mm in length for specimens from Brazil ( Barroso et al. 2017). The California, North Atlantic slope, and Gulf of Mexico material also included some exceedingly small specimens that measured no more than 0.3–0.5 mm in length; these usually had fewer than 11 setigers (i.e., 8 or 9 setigers) and are considered to be juveniles.
The present description is based primarily on 10 specimens from Antarctica, including the type specimens and eight additional specimens collected by Dr. J.A. Blake during ANDEEP surveys in 2002–2005. Extensive collections (593 specimens in all) from the South China Sea, the abyssal Pacific, the North American Pacific and Atlantic coasts, and the Gulf of Mexico were carefully compared to the Antarctic material. All material assigned to A. palpata in this study had 11 setigers (with the exception of the few juveniles mentioned above), parapodia with anterior and posterior lobes as described for the Antarctic material, and the distinctive hooked and hooded tip on falcigers along the length of the body. Differences in morphology among populations in different locations (e.g., size of the nuchal area, which in some material appeared more as a band than a large oval patch; the degree of biannulation, which was not observed in some material; the number of setae in anterior parapodia, which varied slightly among locations; and the number and ease of observation of internal parapodial glands) were not considered indicative of representing a different species. Specimens referred to A. palpata by Barroso et al. (2017) differ from the Antarctic material examined here in being of a larger size and having more setae in the first setigers, many fewer muscle rows in the proventricle, and longer spiniger-like blades. It is not clear from their description if the Brazilian specimens have the hooked and hooded tips on the falcigers, while Fig. 12G View FIGURE 12 in Barroso et al. (2017) suggests that a small hood might be present, this character should be verified. It is possible that molecular studies would find differences between the various populations, but that technique was not available for this study. Eight additional species similar to A. palpata were separated out after careful morphological examination; those specimens have distinctive characters that distinguish them from A. palpata , including number of setigers, shape of the falcigers, and the development or lack of parapodial lobes.
A further discussion of the variability in several characters of A. palpata is presented below and comparison with other species that have free or partially free palps is presented in Table 1 View TABLE 1 .
Degree of fusion of the palps: The palps were described by Hartman (1967) as “separated to the base” but other authors have referred to the palps as fused halfway ( Aguado & San Martín 2008) or sometimes more than halfway ( Barroso et al. 2017). While many specimens in the present collections had palps clearly separate to the base, some adults had palps that appeared to be joined for half their length. The confusion may arise because when the dorsal surface is in focus, the palps appear separate to the prostomium, but when the focus is adjusted ventrally, the palps appear joined along the lower half. Photographs of living specimens taken by J.A. Blake from Antarctic samples show palps that are clearly separate for the distal half, perhaps with a membrane joining the palps basally; this condition appears similar to that pictured in Barroso et al. (2017, Fig. 12 View FIGURE 12 ).
Nuchal patches: The nuchal patches were described by Hartman as “transverse ridges” but are strikingly large oval patches in the Antarctic material; they are visible without staining, but their size and density are especially obvious when stained with Shirlastain A. They are similarly large in some of the material from California, the Gulf of Mexico, and the South China Sea; other specimens from those collections appear to have smaller, less obvious areas of nuchal cilia and some, from other geographic areas, have nuchal areas that are not obvious, even with stain. Specimens from the US Atlantic continental slope off North Carolina had narrow ridges of cilia rather than large oval patches. It is possible that these areas can contract to various degrees, allowing some of the cilia to be retracted and hidden. Conversely, the size of the patch might be indicative of cryptic species.
Peristomial and tentacular cirri: The tentacular cirri were described by Hartman (1967) as “short, cirriform tentacles which resemble the prostomial antennae.” Both Aguado & San Martín (2008) and Barroso et al. (2017) referred to them as “papilliform,” a term that suggests a much smaller structure than is actually present. They are typically only a bit smaller than the prostomial antennae but, in some specimens, including living specimens from the Antarctic, the peristomium appears swollen and the tentacular cirri are noticeably large.
Pharynx: The pharynx is straight and lacks a medial tooth ( Hartman 1967); the everted pharynx has two crowns with 10 large papillae associated with the distal crown ( Barroso et al. 2017, this study). In most specimens, the papillae are long and oval and retain MB stain, but in some samples from the Gulf of Mexico, many specimens appeared to have a plain rim on the everted pharynx, while others had small papillae and still others had large papillae. These differences might be attributable to difficulty in observing this character in small specimens, but possibly the degree of development of the papillae is related to growth, as the smallest specimens had the least obvious papillae.
Proventricle: The proventricle in A. palpata is distinctly heart-shaped or cordate, with an indented anterior edge and a tapered posterior end that becomes very narrow ( Barroso et al. 2017, this study). Hartman (1967) described it only as “barred”; individual muscle rows are obscure and hard to count reliably even when the proventricle is free of overlying tissue. In this study, the proventricle of A. palpata was estimated to have 30–35 muscle rows (dark bands or stripes) ( Antarctica), ca. 25 (California), ca. 20 + (Gulf of Mexico), and ca. 20–25 (southeastern US slope). Barroso et al. (2017) reported 12 muscle rows for their specimens from Brazil.
Dorsal cirri: Hartman implied but did not state specifically whether dorsal cirri are present on setiger 2, but her illustration ( Hartman 1967: Plate 14B) shows long filiform cirri on all setigers. Aguado & San Martín (2008) used this character in their key (presumably those cirri were present on the non-type specimen that they examined, or they relied on Hartman’s published illustration, which often included more details than those mentioned in her texts). In the present material, dorsal cirri were often mostly lost, but the basal stubs of missing cirri could usually be seen easily; no such bases were noted on setiger 2 of the large majority of specimens although often those same specimens had cirri present on setigers 1 and 3. However, two specimens from off California did have long filiform dorsal cirri on setiger 2 (CASIZ 230481, Fig. 4A,C View FIGURE 4 and LACM-AHF Poly 10492); these exceptions demonstrate the unreliability of using this character, especially at the generic level. Barroso et al. (2017) amended the generic definition to reflect the lack of dorsal cirri on setiger 2 of all four of the then-known Anguillosyllis species. However, the presence of dorsal cirri on setiger 2 of some Anguillosyllis material, including A. palpata and other species described in this study precludes making this a genus-level character.
Parapodial lobes: The dorsal or superior lobe is obvious on all specimens of A. palpata ; it was often visible as a large rounded flap, sometimes extended as a hood over the setae and sometimes curled back toward the midline. Barroso et al. (2017) described a “semicircular flap” on the dorsal surface of the parapodia. This lobe or flap does not appear retractile as described by Day (1963) for A. capensis . Many internal parapodial glands appear to enter this lobe in several if not all of the Anguillosyllis species.
Setae: Hartman’s (1967) original description mentioned composite setae that included both falcigers and spinigers, but although the narrow hooked or twisted tips and the serration of the falcigers were illustrated, they were not specifically described in the text. The falcigers as illustrated also appear to have a small membrane that is reminiscent of the hoods seen in other polychaetes reaching from the tip to the shaft. The obvious serration of both the falcigers and longer spiniger-like blades was seen on the type specimens and new material from the Antarctic. The serrations on the blades were more difficult to see in some collections than in others, with the material from the Gulf of Mexico having the smallest and hardest-to-see serrations (although visible at 1000x); whereas, the blades on the specimens from the US Mid- and South Atlantic had coarse serrations that were more easily seen. The hooked and hooded tips on the falcigers are considered to be a defining character for this species and were seen on all specimens referred to A. palpata in this study. It is not clear if material from Brazil has similar tips on the falcigers, the figured seta may or may not have a small hood ( Barroso et al. 2017, Fig. 12G View FIGURE 12 ).
Number of setae per parapodium: The specimens from Brazil were reported by Barroso et al. (2017) to have 30+ setae in the first few anterior setigers. Most of the material examined in this study had fewer setae in the anterior setigers, usually 20, in a few cases (GOM) as many as 25, but never more than 30.
Length of setal blades: In the Brazilian specimens ( Barroso et al. 2017), the shortest setal blades were reported as measuring 20–30 µm, similar to the 20–50 µm measured for the Antarctic material; the longest blades measured 320–450 µm, which is twice the length measured here for the Antarctic material (200 µm).
Aciculae: The two aciculae per parapodium were not mentioned by Hartman (1967) but were seen by Barroso et al. (2017) and this study. The specimens from Brazil have aciculae that are subdistally slightly enlarged and distally acute, whereas those from the Antarctic sometimes have bent tips but no noticeable subdistal enlargement. All aciculae are encased in parapodial tissue and are not emergent.
Parapodial glands: Almost all material had elongated tubular glands in the middle to distal sections of the parapodia. The glands are usually oriented diagonally across the parapodium and feed into the dorsal lobe or distal portion of the parapodium.
Pygidial cirri: Hartman’s (1967:53) description limited pygidial cirri to two lateral digitate cirri; however, the full complement actually includes four cirri: two long, thin, coiled ones similar to the dorsal cirri and two wider lateral elliptical ones as illustrated by Hartman (1967).
Discussion. The material examined for the present study is consistent across all geographic locations in major morphological characters, including the number of setigers, the shape and degree of (or lack of) fusion of the palps, the shape and size of the proventricle, the presence of obvious dorsal lobes, as well as anterior and posterior parapodial lobes, and the kind and distribution of acicula and setae. Minor variability appears to occur in the number of muscle rows in the proventricle (a very difficult character to observe), the degree of serration of the setal blades (which could be due to the angle of observation), the number of setae in anterior setigers, and the degree of development of the parapodial lobes (especially in the material from the South China Sea, which often had damaged tissue where the parapodial lobes would be).
At this time, it appears that Anguillosyllis palpata is very widespread, reported from the Antarctic to the South China Sea, and from depths ranging from 210 to 4652 m. Further investigations as to whether any of these populations represent separate species will necessitate the use of additional technologies, including SEM and molecular studies.
Records. Antarctica: Drake Passage , Bransfield Strait, Weddell Sea, 210–4652 m . Brazil: continental slope 393–3028 m . Gulf of Mexico : off Louisiana, 229– 955 m. Western Atlantic : off Delaware, 2100 m ; southeastern U.S. continental slope and rise, off North Carolina and South Carolina, 583–3012 m (most common at 800–1000 m) . Eastern Pacific : off San Francisco, California, 1780–3864 m . South China Sea : off Brunei, 1224–2135 m


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anguillosyllis palpata ( Hartman, 1967 )
| Maciolek, Nancy J. 2020 |
Anguillosyllis palpata
| Barroso, R. & Paiva, P. C. & Nogueira, J. M. M. & Fukuda, M. V. 2017: 422 |
| Aguado, M. T. & San Martin, G. 2008: 38 |
Braniella
| Hilbig, B. & Blake, J. A. 2006: 269 |
| Blake, J. A. & Grassle, J. F. 1994: 850 |
| Hilbig, B. 1994: 944 |
Braniella palpata
| Blake, J. A. & Maciolek, N. J. & Ota, A. Y. & Williams, I. P. 2009: 1797 |
| Hartman, O. 1967: 55 |