
Podonomus mina, Shimabukuro, Erika Mayumi, Pepinelli, Mateus & Trivinho-Strixino, Susana, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4290.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:7E616304-B616-405D-98A4-5B08E76A300B |
|
DOI |
https://doi.org/10.5281/zenodo.6020162 |
|
persistent identifier |
https://treatment.plazi.org/id/71A6C38D-8C2E-4C6C-8AFC-F3F00DAB31D6 |
|
taxon LSID |
lsid:zoobank.org:act:71A6C38D-8C2E-4C6C-8AFC-F3F00DAB31D6 |
|
treatment provided by |
Plazi |
|
scientific name |
Podonomus mina |
| status |
sp. nov. |
Podonomus mina sp. nov.
Type material. Holotype: male, Brazil, Minas Gerais State, Passa-Quatro, Serra da Mantiqueira, Pedra da Mina Mountain , 22˚25'51''S/44˚50'590''W, 2.700m a.s.l., 14.x.2014, madicolous habitat, leg. E.M. Shimabukuro, G. Perbiche-Neves & V.S. Saito (N1-29, LEIA-UFSCar) . Paratypes: 3 males (N 1-30-32, LEIA-UFSCar), 2 females (N1-33-34, LEIA-UFSCar), 3 pupal exuviae (N1-35-37, LEIA-UFSCar), and 1 pharate pupa with female (N1-38, LEIA-UFSCar), same data as holotype. Two males from Espirito Santo State, Serra do Caparaó, Pico da Bandeira , Vale Verde , 41°50′45.6″S / 20° 25′ 12″W, 1276 m a.s.l. 18.ix.2012, leg. M. Pepinelli (N1-39-40, LEIA-UFSCar). GoogleMaps
Etymology. Refers to the name of the type-locality: Pedra da Mina Mountain. The specific epithet is treated as a noun in apposition.
Diagnostic characters. The new species can be separated from other Podonomus species by combination of the following characters. Male: presence of a large and rounded swelling entirely covered by microtrichia in the subapical region of the gonostylus; dorsomedian surface of gonocoxite with a group of stout setae. Female: posterior margin of cerci obliquely cut, four setae present at the distal margin of cercus: two large setae in the outer lobe and the inner lobe with upper larger setae and lower smaller setae. Pupa: small, less than 2 mm; lateral processes of segments III and IV curved; large ventral lamella on segment II; five sinuous setae on each side of anal lobe; large anal spur on anal lobe; long plastron plate large and round apically, with proximal margin beginning in the middle of respiratory atrium; respiratory atrium with a lateral margin strongly curved distally.
Descriptions. Male (n=6). Total length 1.18–1.38 mm. Wing length 1.63–1.70 mm. Total length/wing length 0.7–0.86. Wing length/length of profemur 2.2–3.0.
Colouration: Black; wings light brown; legs uniformly dark brown
Head. Antenna with 14 flagellomeres. AR 0.43. Presence of scapal setae 62–66 µm (3) on pedicel. Last flagellomere (1) 61 µm; XIII flagellomere (1) 169 µm. Temporal setae 1 2–21. Clypeus with 2–7 setae. Strong seta on tentorium, 62–77 µm. Palp segment lengths (II-V) 28–62, 94–125, 72–81, 66–94. Third palpomere with 3–4 sensilla clavata (3) 26–38 µm long. Eyes bare.
Thorax. 16–25 acrostichals in mid scutum, 11–24 dorsocentrals, 7–17 antepronotals, 3–6 prealars, 1–3 supraalars, scutellum with 3–6 setae, 2–4 prescutellars.
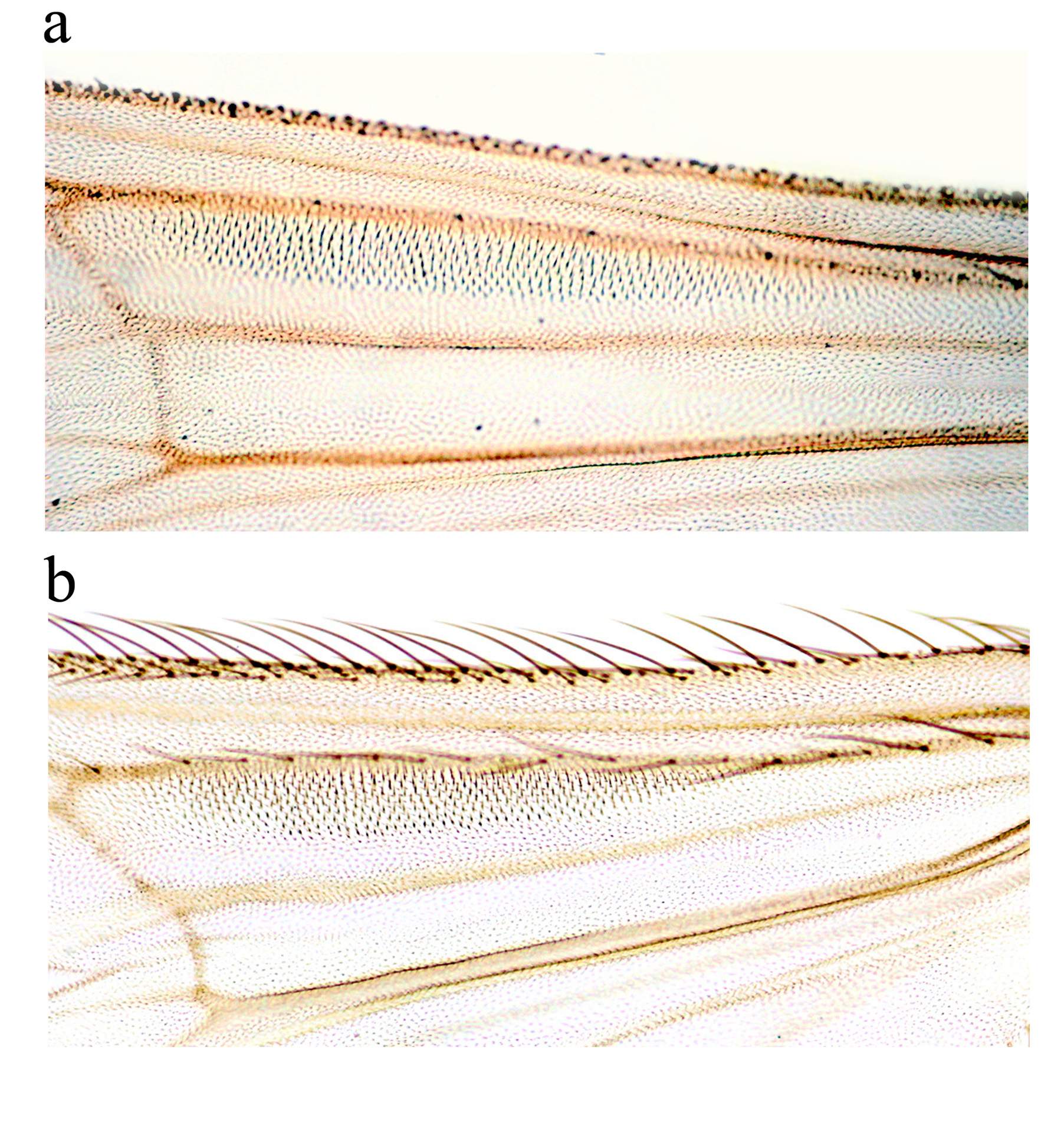
Wing. Membrane with a patch of perpendicular setae on r cell ( Figure 1 View FIGURE 1 a). VR 0.89–0.92 (2). Costal extension (4) 91–126 µm long. r1 cell (4) 77–86 µm wide, r4+5 cell (4) 112–132 µm wide. R with 9–14 setae, R1 with 1–6, R4+5 with 0–3 setae. Brachiolum and squama with 3–5 setae each.
Legs. Spur of fore tibia 46–49 µm long, spurs of mid tibia 26–31 µm and 31–46 µm long, spurs of hind tibia 31–41 µm and 77–89 µm long. Tibial comb (4) with 10–13 bristles. Lengths and proportions of legs as in Table 2.
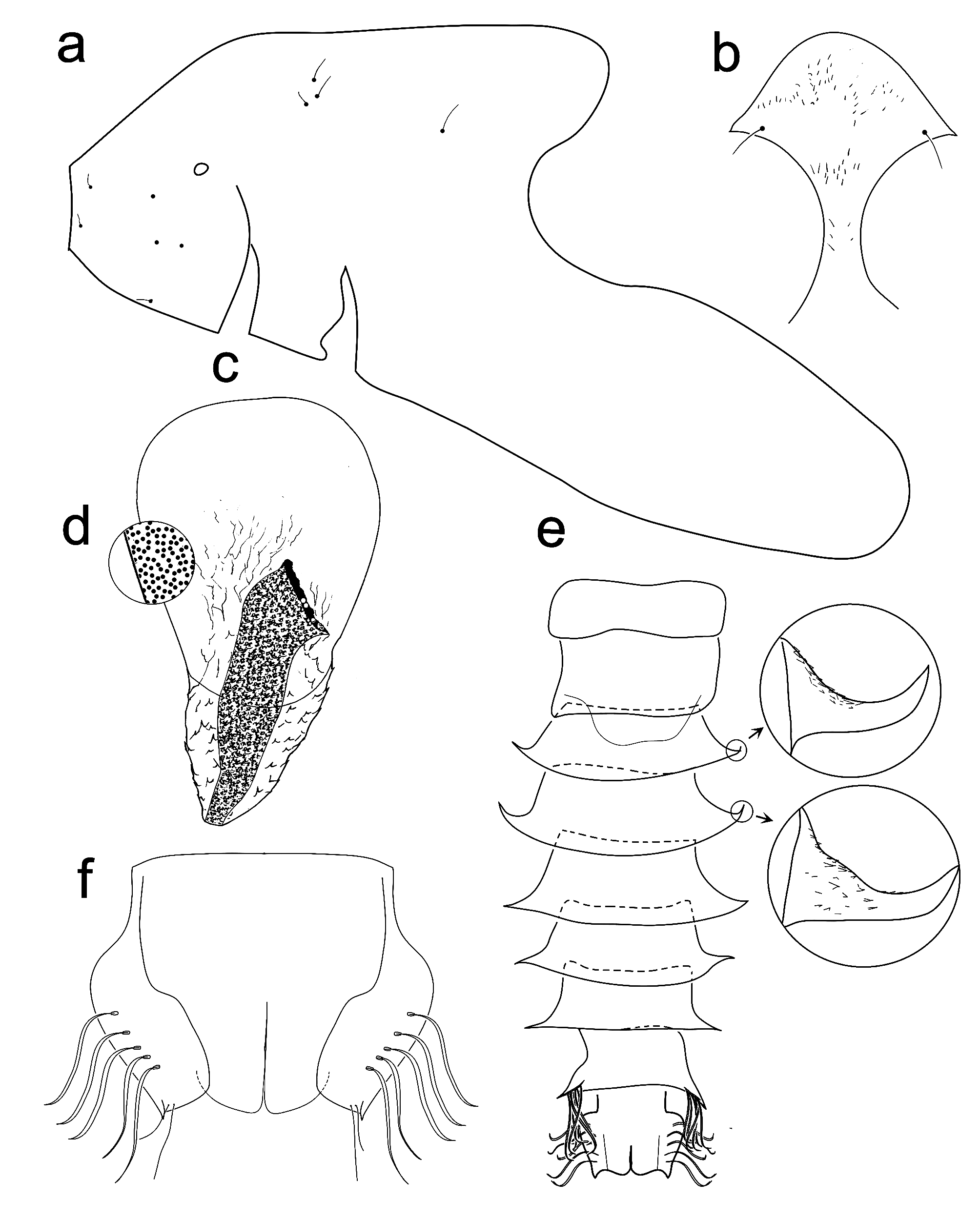
Hypopygium ( Figure 2a View FIGURE 2 ). Gonocoxite 127–161 µm long with a group of stout setae in dorsomedian extension, and strong setae externally. Phallapodeme (4) 52–89 µm long; transverse sternapodeme (3) 50–59 µm long.
Gonostylus 46–58 µm long ( Figure 2b View FIGURE 2 ). True subapical lobe absent, only a large swell covered by microtrichia present; P, x and y setae not conspicuous; long apical lobe 12–15 µm with a t setae of normal type 13–25 µm.
Female (n = 2). Total length 1.2–1.4 mm. Wing length 1.6–1.7 mm. Total length/wing length 0.71–0.85. Wing length/length of profemur 2.9–3.5.
Colouration: Black; wings light brown; legs uniformly dark brown.
Head. Antenna with 9 flagellomeres ( Figure 2c View FIGURE 2 ), last flagellomere 92 µm; I to VIII flagellomeres 261–277 µm. AR 0.33–0.35. Temporal setae 7–8. Clypeus with 3 setae. Palp segment lengths (II–V): 26–31, 43–94, 35–57, 37– 46 µm. Third palpomere with 2–3 sensilla clavata in apical third; 26–28 µm long. Eyes bare.
Thorax. 20–26 acrostichals, 3 antepronotals, 24–27 dorsocentrals, 21–24 prealars, 1 supraalar; scutellum with 7–8 setae, prescutellars 4–5 setae.
Wing ( Figure 2d View FIGURE 2 ). Costal extension 66–77 µm long. r1 cell 118–125 µm wide. r4+5 cell 142–155 µm wide. R with 15–17 setae. R1 distally swollen with approximately 30 setae. R4+5 with 23–32 setae. Brachiolum with 2 setae and squama with 3–4 setae.
Legs. Spur of fore tibia 43 µm long, spurs of mid tibia 26–31 and 35–38 µm long, spurs of hind tibia 37–38 µm and 75–80 µm long. Long setae on outer margin of hind tibia 108–129 µm. Tibial comb with 8–9 bristles. Lengths and proportions of legs as in Table 3.
Genitalia ( Figure 2e View FIGURE 2 ). Cercus 108–117 µm long, with 4 setae in the apex. Postgenital plate 86–108 µm long. Gonocoxapodeme 105 (1) µm long. Coxostenapodeme 34 (1) µm long. Seminal capsules 31–32 µm long. Notum 98–111 µm long.
Pupa (n = 3). Total length 1.7 (1) mm. Exuviae brown.
Cephalothorax ( Figure 3a View FIGURE 3 ). Frontal apotome as in Figure 3b View FIGURE 3 . Thoracic horn 343–364 µm long and 184–195 µm maximum width ( Figure 3c–d View FIGURE 3 ). Plastron plate large and rounded apically, 231–249 µm long; proximal margin of plastron plate starting in the middle of respiratory atrium; respiratory atrium 197–231 µm long. Respiratory atrium with distal portion strongly curved. Dc 1 23–49 µm long, Dc2 (2) 15–33 µm long, Dc3 (1) 35 µm long. Sa 7–24 µm long. Longest median antepronotal 31–52 µm long. Lateral antepronotal 29–63 µm long. Precorneals 5 µm long. Distance between Dc1 and Dc 2 18–30 µm, between Dc2 and Dc 3 17–29 µm, between Dc3 and Sa 183–207 µm ( Figure 3a View FIGURE 3 ).
Abdomen ( Figure 3e View FIGURE 3 ). Ventral lamella present. Segment II to VIII with lateral processes. Segments III and IV with lateral processes slightly curved. TI–TII bare, TIII–TVIII with fine and sparse spinules, without coarse shagreen. TIX with relative coarse anterior spinules. SI–SII bare; SIII–SVIII with fine, sparse spinules; SIX with anterior spinules. Segment VIII with 6–9 L setae, lengths (in µm) of L1 to L7 respectively: 338–400, 338–346, 331–338, 338–374, 300–338, 292 (1), 331 (1). Anal lobe ( Figure 3f View FIGURE 3 ) 215–226 µm long with 5 sinuous setae; distal macrosetae on anal lobe with distinct sizes, smaller with 42 µm long, larger with 87 µm long. Genital sac of male pupae (1) 238 µm long reaching apex of anal lobe; presence of a large anal spur 9 µm long.
Notes on distribution and biology of the species. P. mina sp. nov. was found in madicolous habitats of high mountains in southeastern Brazil. The species was recorded in temporary films of water that percolates exposed rocks above 2700 m a.s.l. in Pedra da Mina Mountain. At the same mountain, P. mina sp. nov. was found at the margin of a streambed, situated at 2500 m a.s.l., constantly wet by the water splashing from the rapids. In Pico da Bandeira (Serra do Caparaó), P. mina sp. nov. was found in the marginal bedrock on the side of the main water channel of Rio Caparaó, where a few specimens were collected at lower altitude of 1270 m a.s.l.. P. mina sp. nov. was also recorded in Pedra do Sino Mountain, in Rio de Janeiro State, at 1580 m a.s.l.
P. mina sp. nov. has been found in the two highest mountains of the southeast Brazil: Pico da Bandeira ( 2892 m a.s.l.) and Pedra da Mina ( 2798 m a.s.l.). However, it was recorded at 1270 m a.s.l. and 1580 m a.s.l., in Pico da Bandeira and Pedra do Sino, respectively, indicating that the distribution of this species occurs in a wide range of altitude (1270–2700 to our knowledge).
Taxonomic discussion. The male hypopygium of Podonomus mina sp. nov. is similar to that of Podonomus albinervis Edwards 1931 , both presenting a pronounced subapical swelling covered with microtrichia, and gonocoxite with an excavated area dorsomedially. However, they differ by the presence of a group of strong setae in the dorsomedial excavated region in P. mina sp. nov. gonocoxite. In addition, P. albinervis possesses a much more robust megasetae on gonostylus and a whitish area that crosses the middle of the wing. The new species also shares many similarities with Podonomus setosus Brundin 1966 , and Podonomus caranqui Roback 1970 , especially by the presence of the conspicuous group of stout setae in the gonocoxite, but differ from the former by the presence of a notable rounded swelling in the gonostylus, and from P. caranqui by the presence of microtrichia covering the swelling.
Although most of Podonomus spp females are superficially described, the female of P. mina sp. nov. is similar to P. pepinellii and Podonomus besti Brundin 1966 , differing from the former by the number of setae in R4+5 and by the shape of distal part of the cerci, which is obliquely cut in the P. mina sp. nov., and from the later by the antennal number of segments. Additionally, while P. besti presents 11 flagellomeres ( Brundin 1966) P. mina sp. nov. has only 9.
The pupa of P. mina sp. nov. resembles P. pepinellii as both have a large and rounded plastron plate with small pores closely arranged. P. pepinellii also presents a lamella on SII and 5 sinuous setae at the anal lobe. Nonetheless P. mina sp. nov. can be separated from P pepinellii by the following characteristics: presence of slightly curved lateral processes on III and IV segments; proximal margin of plastron plate starting in the middle of respiratory atrium; shorter border of respiratory atrium with a distal portion strongly curved.
P. mina sp. nov. does not fit perfectly in any of the five groups proposed by Brundin (1966). Although the gonostylus of the male is similar to those found in the albinervis group, without a true subapical lobe, and presenting only a swelling covered by microtrichia, the size of the swelling seems to be more prominent than other species of the group, except P. caranqui . This feature would transfer the species to the decarthrus group, although species in this group do not have the swelling covered with microtrichia. On the other hand, the female features seem to fit well in the decarthrus group, once the posterior margin of the cerci is obliquely cut and deeply sinuous, also presenting 4 setae apically. Finally, the hypothesis that P. mina sp. nov. belongs to the decarthrus group was also not supported by pupal characters, especially diverging in the number of sinuous setae on anal lobe (only five in P. mina sp. nov.) and the presence of a large lamella in the segment II.
DNA Barcoding. The six new sequences combined with a published dataset on Podonomus specimens (Trivinho-Strixino et al. 2012) confirmed that P. mina sp. nov. is one of the molecularly distinct groups previously revealed by Trivinho-Strixino et al. (2012). However, DNA barcoding was previously performed mostly from larval tissues, and further comparative larval morphological analyses did not support the evidence of several species. With more adults available for analyses, DNA Barcoding was crucial to link temporal and geographical diversity of Podonomus . Adults collected in Pedra da Mina mountain in 2014, were clustered together with larvae collected in Caparaó mountain in 2012, corroborating that one of the molecular distinct groups revealed by Trivinho-Strixino et al. (2012) is in fact a valid species, molecularly and morphologically distinct from P. pepinellii . The other Podonomus clade from Caraça mountain still remains known only from its larval stage ( Figure 4). Maximum intraspecific genetic divergence was 1.13% within P. pepinellii (n=27), 2.6% within P. mina sp. nov. (n=9) and 0.3% within Podonomus sp. (Caraça, n=5). Among the three distinct groups formed, the mean distance between P. mina sp. nov. and P. pepinellii was 8.0%, between P. mina sp. nov. and Podonomus sp. was 10.26% and between P. pepinellii and Podonomus sp. was 10.25%.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Podonominae |
|
Genus |