
Pseudostenhelia bathyalis, Gómez, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.5051.1.11 |
|
publication LSID |
lsid:zoobank.org:pub:82B4BCC0-BE67-4BC2-BE3E-68D39F72CEEE |
|
DOI |
https://doi.org/10.5281/zenodo.5715369 |
|
persistent identifier |
https://treatment.plazi.org/id/D86B0811-F015-43F6-BF3A-9397B4280D20 |
|
taxon LSID |
lsid:zoobank.org:act:D86B0811-F015-43F6-BF3A-9397B4280D20 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudostenhelia bathyalis |
| status |
sp. nov. |
Pseudostenhelia bathyalis sp. nov.
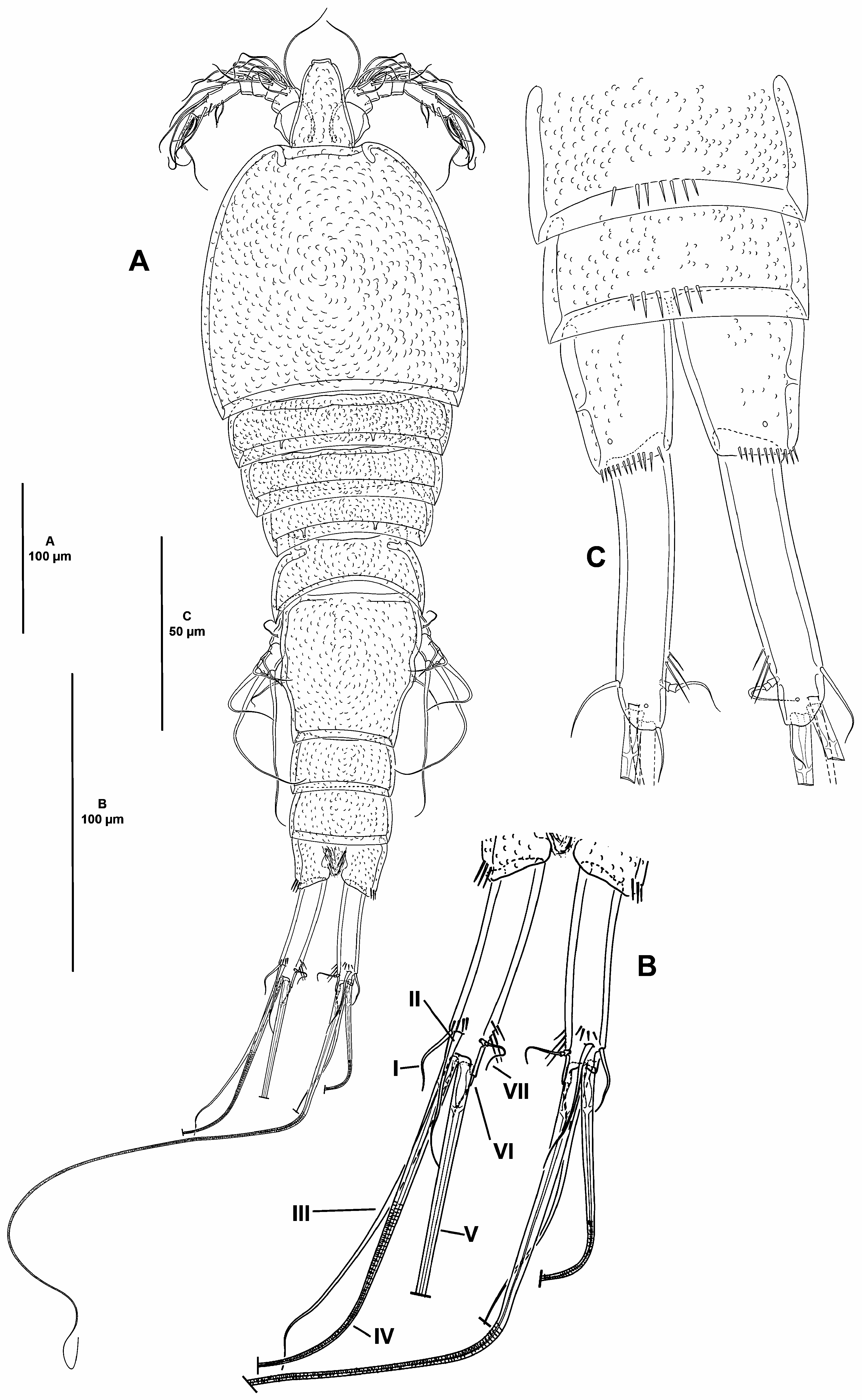
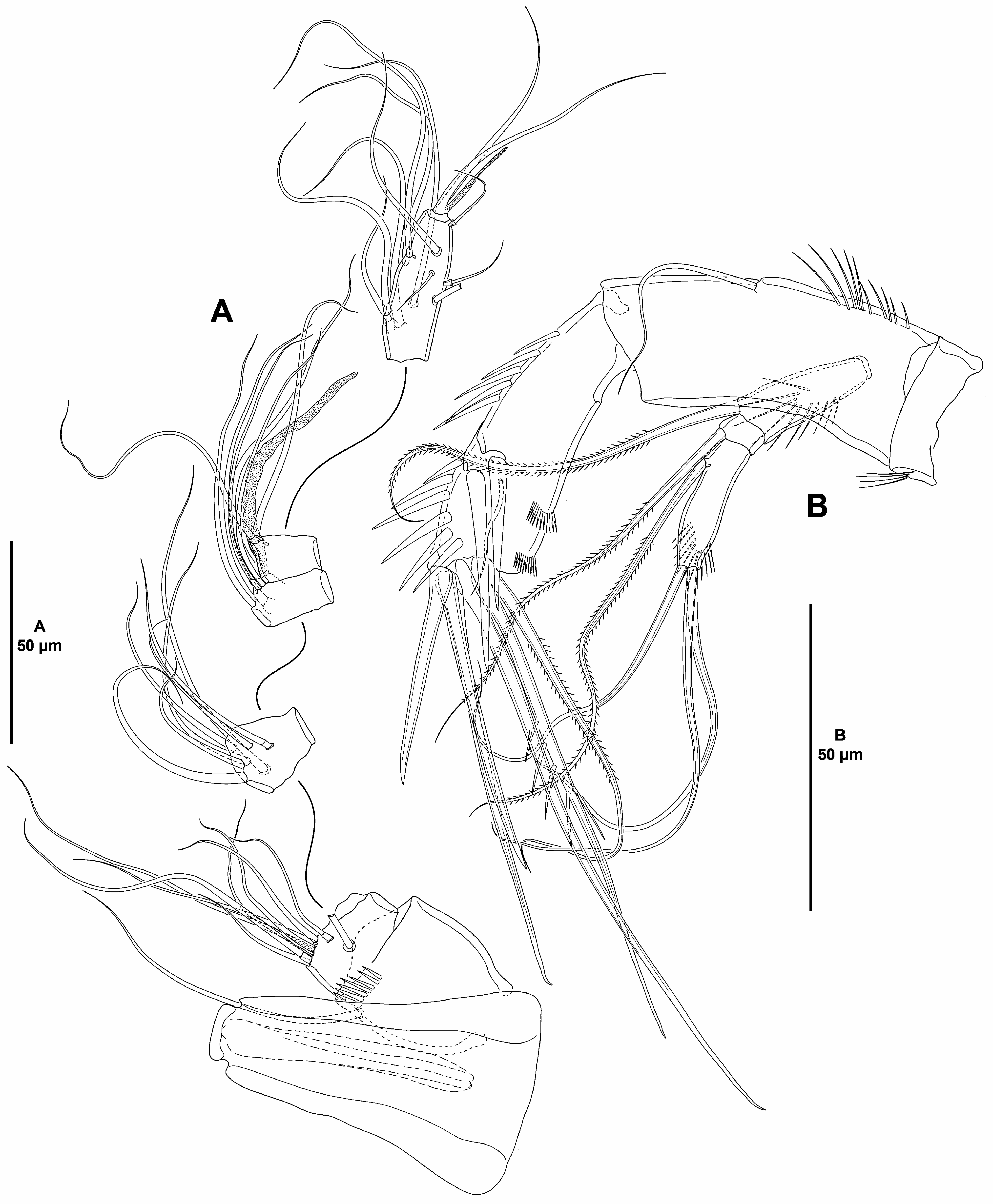
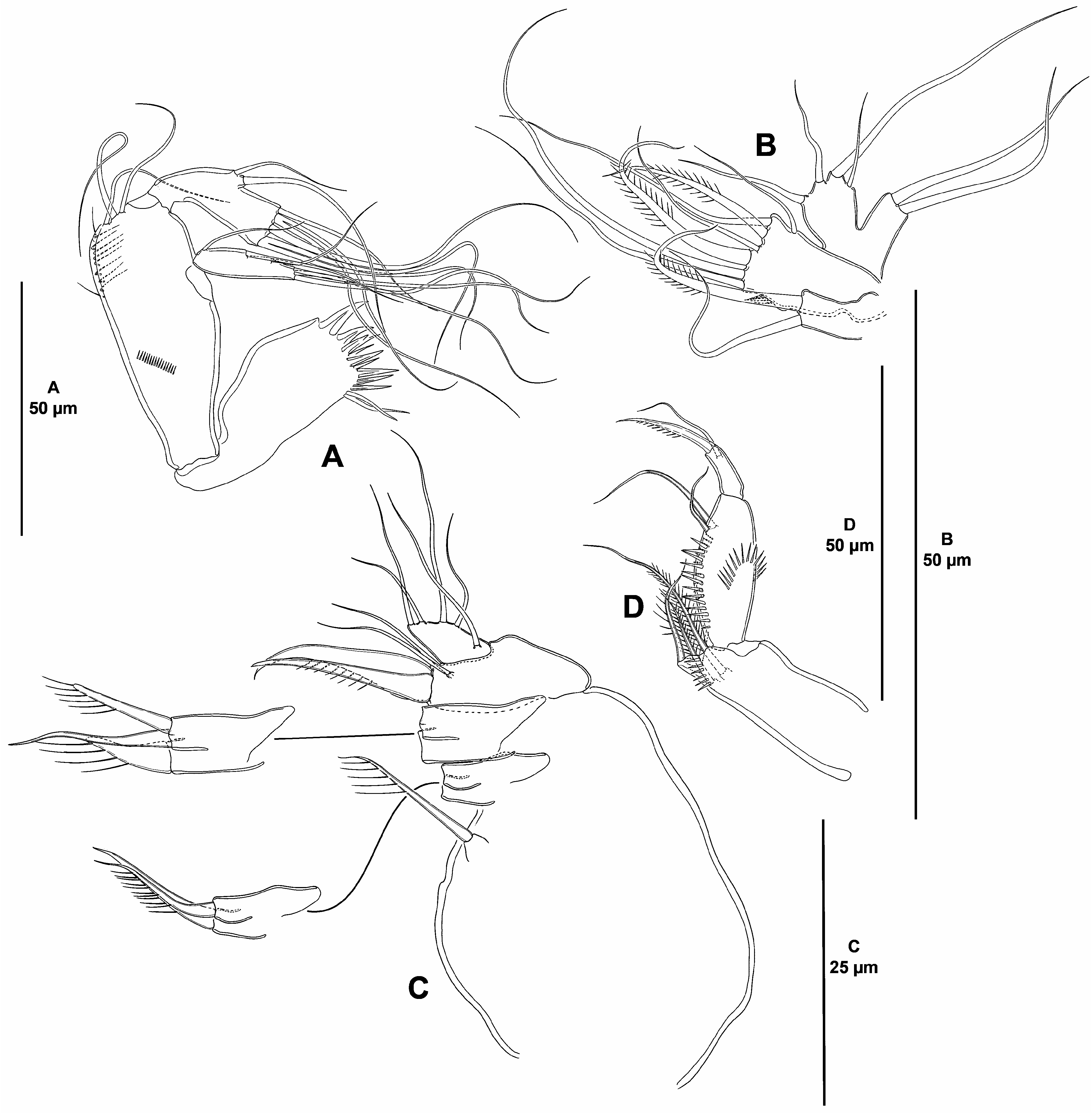
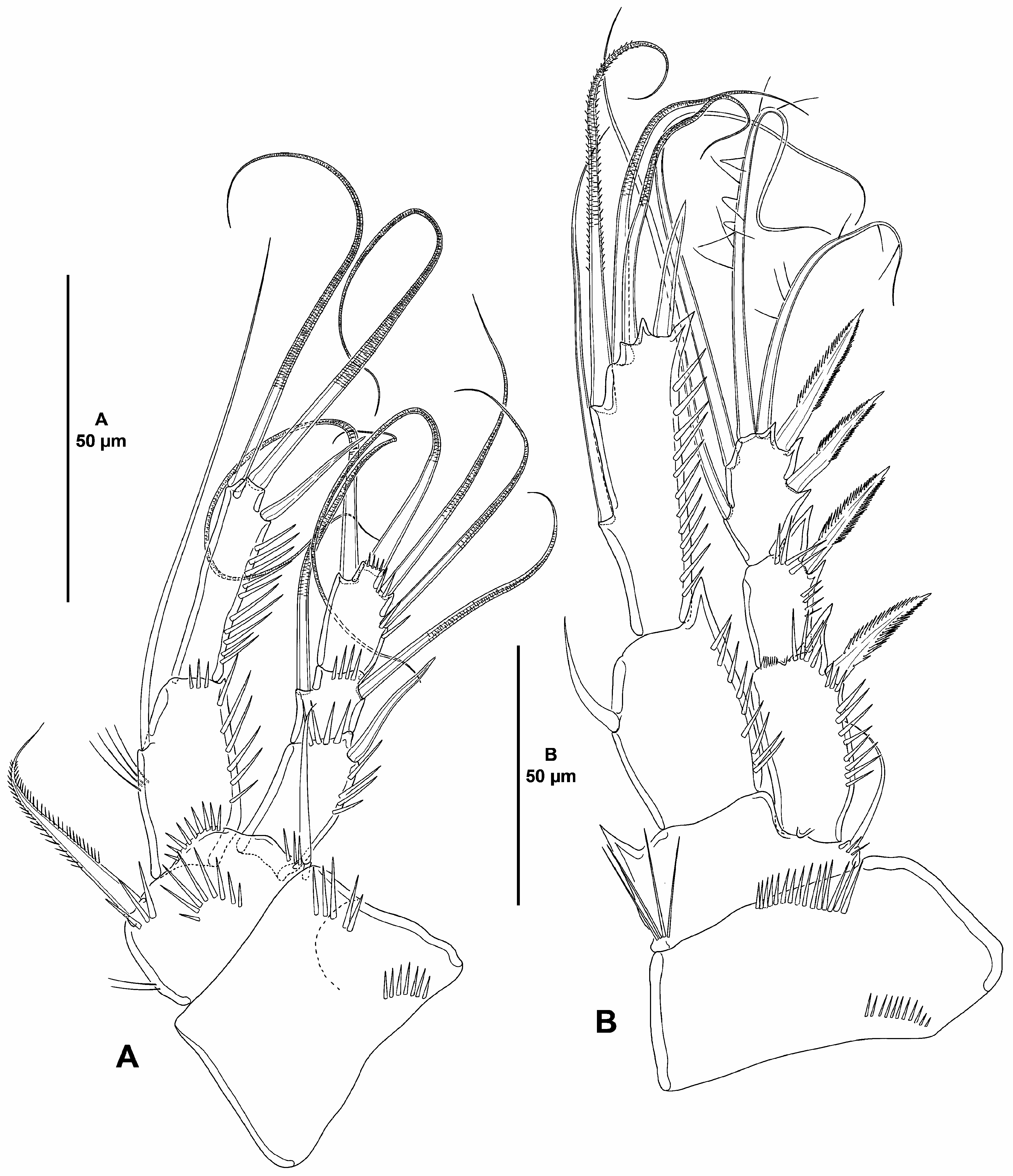
( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
urn:lsid:zoobank.org:act:
Type locality. Southeastern San Quintin Basin , off Bahia del Rosario, Baja California, Mexico; Talud XVIB cruise, sampling station 16 ( 29.81333°N, 116.12166°W); depth, 1,752 m; organic carbon content, 4.08%; organic matter content, 7.01%; sand 5.45%; clay, 11.94%; silt, 82.61 GoogleMaps %.
Specimens examined. Female holotype (EMUCOP-290514-01) dissected and mounted onto seven slides; Talud XVIB cruise, sampling station 16 ( 29.81333°N, 116.12166°W); May 29, 2014; coll. S. Gómez.
Etymology. The specific epithet bathyalis from the Greek βαθύς (bathys), deep, refers the environment where the species was found.
Description of female. Total body length measured from tip of rostrum to posterior margin of caudal rami 620 µm. Habitus pyriform, widest at posterior end of cephalothorax, tapering posteriad ( Fig. 2A View FIGURE 2 ); cephalothorax/body length ratio, 0.3.
Prosome consisting of cephalothorax with fused first pedigerous somite, and second to fourth free pedigerous somites. Cephalothorax ( Fig. 2A View FIGURE 2 ) slightly wider than long, no surface sensilla detected, hyaline fringe smooth, surface covered by small depressions. Free pedigerous somites without expansions laterally nor dorsally, without spinular ornamentation, no surface sensilla detected, integument covered by small depressions, weakly sclerotized, hyaline fringe smooth, width of second to fourth pedigerous somites decreasing progressively ( Fig. 2A View FIGURE 2 ).
Urosome consisting of fifth pedigerous somite (first urosomite), genital double-somite (genital—second urosomite—and third urosomites fused), two free urosomites, and anal somite; urosomites without expansions laterally nor dorsally, integument weakly sclerotized, surface covered by small depressions ( Fig. 2A View FIGURE 2 ).
Fifth pedigerous somite without spinular ornamentation, seemingly without sensilla ( Fig. 2A View FIGURE 2 ), hyaline fringe narrow.
Second and third urosomites completely fused forming genital double-somite, without any trace of division, slightly longer than wide, widest part measured in the middle, no surface sensilla nor spinular ornamentation detected, posterior hyaline fringe smooth ( Fig. 2A View FIGURE 2 ).
Fourth and fifth urosomites seemingly without surface sensilla dorsally ( Fig. 2A View FIGURE 2 ) nor ventrally ( Fig. 2C View FIGURE 2 ), dorsally without ( Fig. 2A View FIGURE 2 ), ventrally with medial row of spinules ( Fig. 2C View FIGURE 2 ).
Anal somite 1.7 times as wide as long ( Fig. 2A, C View FIGURE 2 ), with lateroventral spinules close to joint of caudal rami, seemingly without sensilla, with two lateral and two ventral pores ( Fig. 2C View FIGURE 2 ); anal operculum inconspicuous.
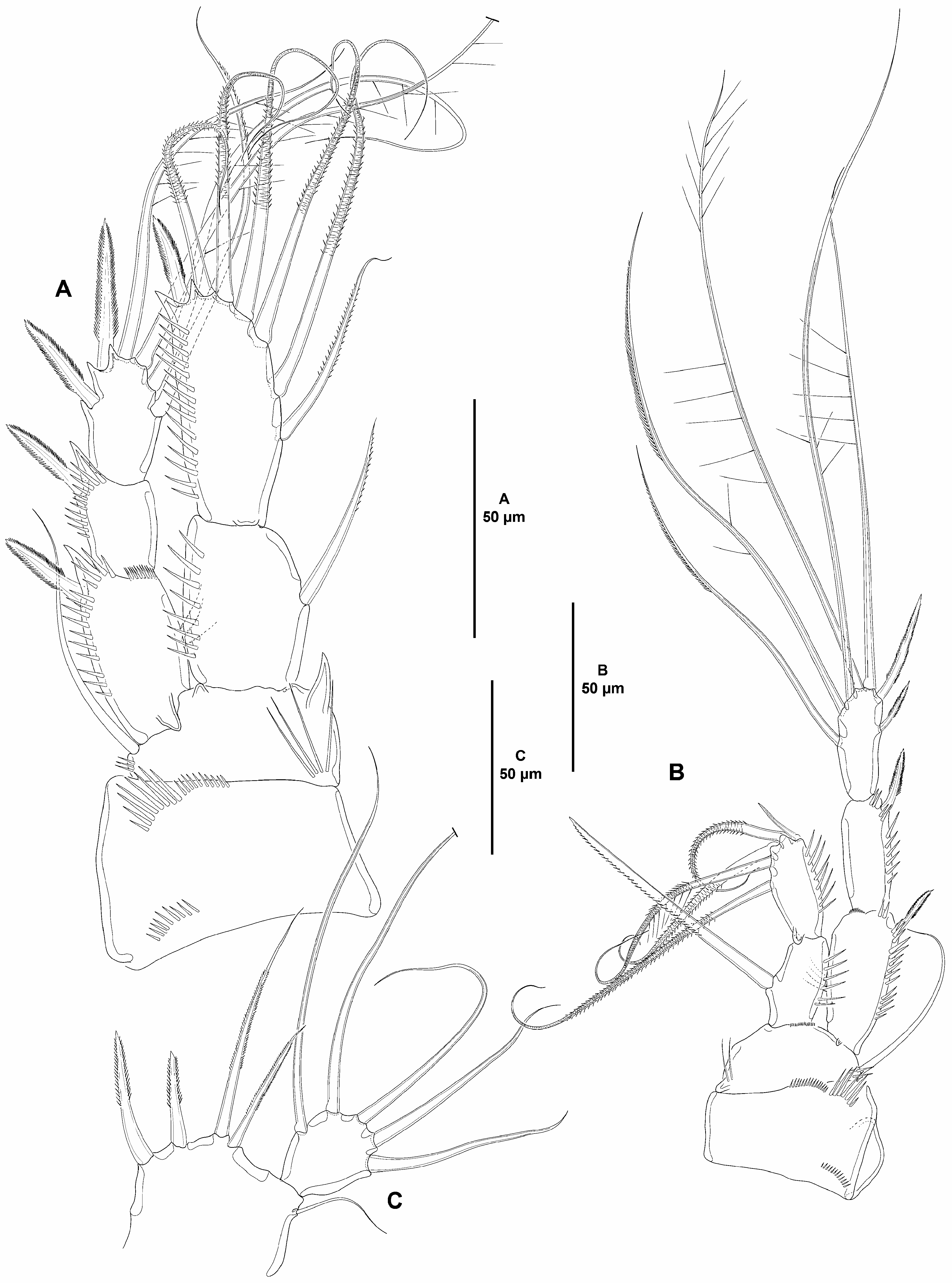
Caudal rami elongate, about five times as long as wide ( Fig. 2A–C View FIGURE 2 ) and about two times as long as anal somite, slightly divergent, each ramus with one subdistal pore ventrally, with dorsal spinules close to seta II, and on inner margin subdistally, with seven elements ( Fig. 2B–C View FIGURE 2 ); seta I and II arising subdistally, the former ventral to and half the length of the latter; seta III subdistal, arising ventrally ( Fig. 2B View FIGURE 2 ; lost in Fig. 2C View FIGURE 2 ); seta IV and V arising distally, both with fracture plane and rat-like, the former shorter; seta VI issuing at inner distal corner; dorsal seta VII triarticulate at base, situated subdistally close to inner margin.
Rostrum ( Fig. 3A View FIGURE 3 ) trapezoidal, not fused to cephalothorax, slightly bifid, subdistal sensilla lost during dissection.
Antennule ( Fig. 3A View FIGURE 3 ) six-segmented, all segments smooth except for spinular row on first segment; all setae smooth; second segment with two, third segment with one seta with fracture plane; sixth segment with two articulated setae. Armature formula: 1(1); 2(9); 3(7); 4(4 + (1 + ae)), 5(2); 6(11 + acro). Acrothek consisting of two setae and one slender aesthetasc fused basally.
Antenna ( Fig. 3B View FIGURE 3 ). Coxa short, with some slender outer spinules. Allobasis as long as free endopodal segment, with inner spinules proximally, with one slender abexopodal seta arising midway inner margin. Exopod threesegmented; first and third segments longest, each about three times as long as wide, and with spinules as shown; first and second segment with one pinnate seta each, third segment with one pinnate proximal and three smooth apical setae. Endopod with strong inner spinules, laterally with two spines and one slender seta, distally with six elements (outermost pinnate seta fused basally to strongly spinulose adjacent element), outer margin with two subdistal frills.
Mandible ( Fig. 4A View FIGURE 4 ) with relatively short coxa. Gnathobase wide, ventral distal corner produced into small sharp semi-hyaline process, with several acute teeth distally, and two smooth setae ventrally. Basis elongate, with narrow base, widest medially, with spinules as shown, with three subdistal setae. Exopod elongate, about four times as long as wide, tapering distally, with three lateral and three distal setae. Endopod recurved, twisted over exopod, with two lateral and six distal elements (two pairs of which fused basally).
Maxillule ( Fig. 4B View FIGURE 4 ). Praecoxa and arthrite lost during dissection. Coxa with one pinnate and one smooth seta. Basis with small inner spinules, and with seven elements of which two pinnate. Exopod and endopod onesegmented, confluent, not fused to supporting basis; endopod larger than exopod, the former with four, the latter with two setae.
Maxilla ( Fig. 4C View FIGURE 4 ). Large syncoxa seemingly without spinular ornamentation, with three endites; proximal endite smallest, with one seta; middle endite with one smooth and one spinulose seta; distal endite with one smooth and two spinulose elements. Basis drawn out into strong spinulose claw, additionally with strong naked spine and two slender setae. Endopod about 1.5 times as long as wide, with five slender setae (one arising basally, two medially, and two apically).
Maxilliped ( Fig. 4D View FIGURE 4 ) subchelate. Syncoxa elongate, rectangular; about 2.5 times as long as wide, with inner spinules subdistally, with two pinnate setae issuing subdistally at the same level, and one distal pinnate element arising from pedestal. Basis slightly shorter than syncoxa, oval, with arched row of outer spinules, with one anterior and one posterior inner spinular row as depicted, with two slender subdistal setae, one of which visibly longer. Endopod one-segmented, with one claw-like spinulose element and one smooth seta.
P1 ( Fig. 5A View FIGURE 5 ). Coxa massive, trapezoidal, with proximal and subdistal outer spinules. Basis with few inner slender spinules, and with stronger spinules medially, between rami, and at base of outer and inner spines, of which the former smooth, the latter pinnate. Exopod three-segmented, reaching middle of ENP2; first segment longest, second segment shortest, length ratio of EXP1–EXP3 1, 0.5, 0.8; all segments without outer nor inner acute distal processes; EXP1 with outer and distal spinules, with outer spine, without inner armature; EXP2 with distal spinules, with outer and inner rat-like setae; EXP3 with two outer, one apical and one inner rat-like element, with spinules at base of outer and distal element. Endopod two-segmented, segments without inner nor outer acute distal processes; ENP1 reaching middle of EXP2, 1.8 times as long as wide and slightly shorter than ENP2, with slender long inner spinules, with strong outer and with small distal spinules, with one long inner seta; ENP2 slenderer than ENP1, 4.5 times as long as wide, with strong outer spinules, with one inner and one distal rat-like seta, and one outer spine.
P2–P3 ( Figs. 5B View FIGURE 5 , 6A View FIGURE 6 ). Coxa shorter but wider than in P1, with one proximal and one subdistal row of spinules. Basis with acute inner process, with set of long inner spinules proximally, and with small spinules at base of outer seta. Exopod three-segmented, shorter than ENP; of P2 reaching middle, of P3 reaching distal third of ENP2; EXP1 longest, length ratio of P2/P3 EXP1–EXP3 1, 0.6, 0.7/1, 0.6, 0.8; EXP1 and EXP2 with outer and subdistal outer spinules, with outer acute process, EXP1 with, EXP2 seemingly without inner hyaline frill, EXP3 without spinular ornamentation; EXP1 and EXP2 without inner armature; EXP3 with two inner and two apical setae, and two outer spines. ENP arising from pedestal, at a higher level than EXP, two-segmented; ENP1 0.7 times as long as ENP2; ENP1 and ENP2 with longitudinal row of strong outer spinules; P2 and P3 ENP1 with one inner element, of P2 visibly shorter; P2 ENP2 with five, P3 ENP2 with seven setae/spines. No pores detected on EXP nor on ENP.
P4 ( Fig. 6B View FIGURE 6 ). Coxa largely as in P2 and P3. Basis largely as in P2 and P3, but without inner acute process, and with both rami arising at the same level. Exopod three-segmented, two times as long as ENP; EXP1 longest, length ratio of EXP1–EXP3 1, 0.9, 0.9; EXP1 and EXP2 with outer and subdistal outer spinules, without acute outer process; EXP1 with, EXP2 seemingly without inner hyaline frill; EXP3 without spinular ornamentation; EXP1 and EXP2 without inner armature; EXP3 with three inner and two apical setae, and two outer spines. ENP two-segmented; ENP1 slightly shorter than ENP2; ENP1 and ENP2 with longitudinal row of strong outer spinules; ENP1 with one strong and stiff inner element; ENP2 with five setae/spines. No pores detected on EXP nor on ENP.
Setal formula of swimming legs as follows:
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | 0,1,112 | 0,0,222 | 0,0,222 | 0,0,322 |
| ENP | 1,111 | 1,221 | 1,421 | 1,221 |
P5 ( Fig. 6C View FIGURE 6 ). BENP pentagonal, poorly developed, without spinular ornamentation; endopodal lobe with four setae as shown; first and second outermost setae close to each other. Exopod oval, nearly as long as wide, without spinular ornamentation, with five setae. No pores detected on BENP nor on EXP.
P6 lost during dissection.
Male. Unknown.
Remarks and discussion. The genus Pseudostenhelia accommodates three species, P. prima , P. secunda , and P. wellsi . Members of the genus have a preference for shallow brackish conditions with detritus-rich fine sediments (mud and fine sand). The discovery of the female of P. secunda in plankton samples from Lake Kolleru ( India) ( Ranga Reddy 1984) is, to the best of my knowledge, the only report of a species of this genus from freshwater habitats. Pseudostenhelia prima , the first species assigned to this genus, was found in fine sediments (mud or muddy sand) from Saco da Inhaca, a shallow inlet south of Inhaca Island ( Mozambique) ( Wells 1967). The male of P. secunda , the second species included in this genus, was found in muddy sediments from the estuary of the Vellar river near Parangipettai (formerly Porto Novo) in the Tamil Nadu State ( India) ( McIntyre 1968; Wells 1971), and the female was found in a freshwater lake (see above; Ranga Reddy 1984). Finally, P. wellsi was found in muddy intertidal sediments from South Carolina ( USA) ( Coull & Fleeger 1977).
The relationships between Pseudostenhelia and some other stenheliins are not clear. Dahms et al. (2005) proposed some apomorphies for Pseudostenhelia ( Dahms et al. 2005: 12, table 2) based on naupliar morphology. They also gave a list of characters shared by Stenhelia , Delavalia , and Pseudostenhelia , viz. the extremely foreshortened cephalic shield, the presence of a spinous process on the distal third (middle?; Huys & Mu 2008) of the second antennulary segment and a notched inner seta on the third antennulary segment, and a lobate outer protuberance of the first segment of the exopod of the antenna. Huys & Mu (2008) questioned the phylogenetic significance of these characters, but admitted that they appear to be unique within Harpacticoida .
Huys & Mu (2008) drew attention to a core group of Stenheliinae ( Onychostenhelia , Cladorostrata , and Delavalia ) sharing a synapomorphic confluent maxillulary exopod and endopod not fused to the supporting basis. This condition is present also in Wellstenhelia ( Karanovic & Kim 2014; Gómez & Cruz-Barraza 2021) and Lonchoeidestenhelia ( Gómez, 2020) and is confirmed here for Pseudostenhelia . Note that Ranga Reddy (1984) described the maxillulary rami as discrete and separated from basis, but this is most probably erroneous. The significance of this condition in the maxillulary rami is not clear but Huys & Mu (2008) suggested that the confluent maxillulary rami separated from the basis could be a strong synapomorphy for Onychostenhelia , Cladorostrata and Delavalia , but also for Lonchoeidestenhelia . However, its presence in some other stenheliin lineages ( e.g. Pseudostenhelia , and Wellstenhelia ) seems to indicate convergent evolution (see also Gómez & Cruz-Barraza 2021).
Pseudostenhelia shares a plesiomorphic anal operculum (semicircular, relatively small, weakly chitinized and flanked by a sensilla on each side) with Melima and Delavalia ( Willen 2003) , but also with Wellstenhelia (see also Gómez & Cruz-Barraza 2021), and it is completely absent in Stenhelia and Anisostenhelia ( Mu & Huys 2002) , and Onychostenhelia ( Huys & Mu 2008) .
Although its significance is not clear, the caudal seta I is either a spine-like element or a reduced seta in most stenheliins, but it is a well-developed seta (most probably plesiomorphic) in Pseudostenhelia and Onychostenhelia , and some species of Delavalia ( Gómez & Cruz-Barraza 2021) .
In addition to the synapomorphic nature of the structure of the maxillulary rami in Onychostenhelia , Cladorostrata , and Delavalia (see above), Huys & Mu (2008) suggested that the resemblance in leg segmentation and armature between Onychostenhelia and Pseudostenhelia observed by Itô (1979) had little phylogenetic significance. However, as noted above (see also Gómez & Barraza 2020), the phylogenetic importance of the confluent maxillulary rami separated from the supporting basis, the shape of caudal seta I, and anal operculum is questionable. On the other hand, the shape of P1 EXP (EXP3 smaller than EXP2 and EXP3), giving the entire ramus a characteristic appearance (except for the somewhat longer P1 EXP 3 in P. bathyalis sp. nov.), the exceedingly long outer elements of P1 EXP2 and EXP3, and the two-segmented endopod of P2–P4, seem to be strong synapomorphies for these two genera.
Both sexes are known for all Pseudostenhelia species except for P. bathyalis sp. nov. The female of the new deep-sea species can be readily separated from the females of its congeners by the segmentation of the antennule (with six segments in the new species, but with five segments in other species), and by its armature complement of P2 ENP2 and P3 ENP2, with five and seven setae instead of four and five setae respectively. The minute proximal endite of the maxillary syncoxa with one seta only, and the reduction of the armature complement on the P5 EXP from six to five setae seem to be the only potential apomorphies for the species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Stenheliinae |
|
Genus |