
Crella (Pytheas) santacruzae Fernandez, Gastaldi, Thompson & Hajdu, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5052.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:FED635BA-B982-400E-B920-1DBA22025EA9 |
|
DOI |
https://doi.org/10.5281/zenodo.5572191 |
|
persistent identifier |
https://treatment.plazi.org/id/15003760-FF86-F66E-6591-FB6BFE6DFD22 |
|
treatment provided by |
Plazi |
|
scientific name |
Crella (Pytheas) santacruzae Fernandez, Gastaldi, Thompson & Hajdu |
| status |
sp. nov. |
Crella (Pytheas) santacruzae Fernandez, Gastaldi, Thompson & Hajdu View in CoL , sp. nov.
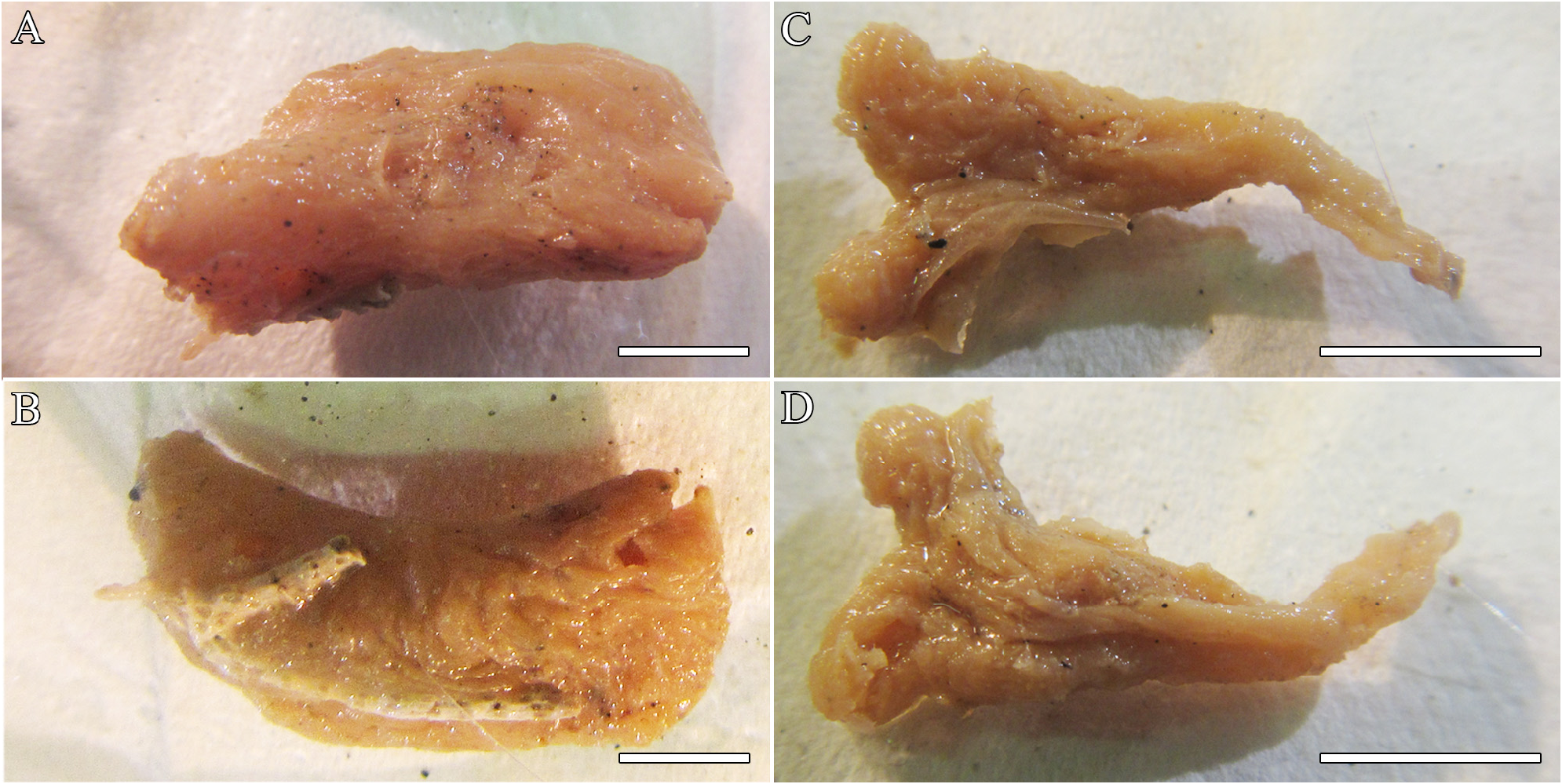
( Tables 2–3 View TABLE 2 ; Figures 6–7 View FIGURE 6 View FIGURE 7 )
Type locality. off Santa Cruz province, Argentina sea .
Material examined. Holotype — ZIN 12081, Stn. 128 (- 47.28333333 S, - 59.90000000 W; off Santa Cruz province, Argentine Sea ), 750 m depth, coll. RV “ Zund ”, 18 th March 1974. Fragment of holotype und MNRJ 22479 View Materials GoogleMaps ; Paratype — ZIN 12082, same data as the holotype. Fragment of GoogleMaps Paratype under MNRJ 22438 View Materials .
Diagnosis. Crella (Pytheas) occurring in the deep southern Atlantic, with tornotes (427–576/7.8–12 µm), two categories of acanthostyles (choanosomal, 375–563/9.1–13.5 µm; ectosomal, 92–119/4.3–7.1 µm) and a single category of arcuate isochelae (22–30.9 µm).
Description. Only two, small, preserved fragments ( Figs. 6A–D View FIGURE 6 ); holotype, 2.85 (L) x 1.28 (W) x 0.6 cm (H) and paratype, 2 (L) x 1 (W) x 0.8 cm (H). Surface irregular, slightly hispid, folded and wrinkled at parts, or smoother, with a thin, easily detachable membrane. Oscules and pores not evident in the fragments. Consistency compressible and resilient, texture soft. Color in spirit dirty beige.
Skeleton. Plumose architecture ( Fig. 7A View FIGURE 7 ). Ectosomal region with tornotes in bundles (up to 12 spicules) and in bouquets, which rarely protrude to the surface, up to 50 µm high (Figs. B–D). Ectosomal acanthostyles in tangential to paratangential arrangement at the surface making a continuous layer, ca. 50 µm thick ( Figs. 7D–F View FIGURE 7 ). Choanosomal region with tornotes in tracts (up to 12 spicules). Thick tracts of choanosomal acanthostyles (more than 15 spicules) at the base of the choanosome. Several ectosomal acanthostyles and arcuate isochelae occur scattered throughout the choanosome. All categories of acanthostyles (ectosomal and choanosomal) are distinguished by size (length and width) and patterns of spination. In spongin there are wide rounded subectosomal (150–700 µm longer length) and choanosomal channels (ca. 1000 µm longer length). No spongin fibers.
Spicules. Megascleres ( Tables 2–3 View TABLE 2 ). Tornotes ( Figs. 7G–J, 7O–P View FIGURE 7 ), slightly aniso, strongyloid, slightly fusiform, slightly curved to straight, smooth, sometimes with polytylote axis (a few or several tyles, occasionally verrucose); axis may bear very small spines spread all around, or at the tips, in a rough appearance; juvenile forms thinner and scarce: 393– 502.3 (41.3)–576/7.6– 10.1 (1)–12.2 µm. Choanosomal acanthostyles ( Figs. 7K–L, 7Q View FIGURE 7 ), uncommon, straight to slightly curved, bases blunt to slightly swollen (tyle), tips acerate or mucronate; rarely completely smooth, often variably spined; spines sharp but sometimes rounded, up to 5.4 µm high, mostly concentrated at the basal third, a few scattered elsewhere, straight or curved towards the tips: 372– 484.2 (53.2)–563/9.1– 11.6 (1.1)–13.7 µm. Ectosomal acanthostyles ( Figs. 7L–M, 7O, 7R View FIGURE 7 ), straight to slightly curved; isodiametric (seldom fusiform), fully spined, blunt bases (no tyle), tips acerate and sharp; spines variably abundant (more than in choanosomal acanthostyles), sometimes more concentrated at both ends, large (up to 6 µm high), straight and sharp: 92– 108.4 (7.5)–122/4.2– 5.4 (0.8)–7.1 µm. Microscleres ( Tables 2–3 View TABLE 2 ), arcuate isochelae ( Figs. 7N, 7S View FIGURE 7 ), axis slightly curved, alae rounded and slightly short; distance between opposite front alae, ca. 1/3 (or less) of maximum chelae length: 22– 25.2 (2.5)–31 µm.
Ecology. Deep sea species occurring with a diverse set of other sponges: Hymeniacidon sp. , Hymenancora sp. , Iophon sp. , Esperiopsis sp. , Phorbas sp. , Tedania sp. , Latrunculiidae sp., Myxillidae sp., Stelligeridae sp., Haplosclerida sp., Suberitida sp., Hexactinellida sp. The 750 m deep isobath is situated in the domains of the Antarctic Intermediate Water.
Distribution. Known only from its type locality, in the deep south-western Atlantic ( Fig. 1 View FIGURE 1 ).
Etymology. The name ‘santacruzae’ is a reference to the Argentine province Santa Cruz, off which the type locality of the new species is located.
Remarks. The new deep-waters southwestern Atlantic species is set apart from both new shallow-waters southeastern Pacific species described above, mainly in having a single category of choanosomal acanthostyles, much larger than those observed in both Pacific species ( Table 2 View TABLE 2 ).
Among additional Crella (Pytheas) spp., three appear close to C. (P) santacruzae Fernandez, Gastaldi, Thompson & Hajdu , sp. nov.: viz., C. (P.) basispinosa Burton, 1931 ( Burton 1931a) , from Norway, C. (P.) fristedti ( Dendy, 1924) , and C. (P.) novaezealandiae ( Bergquist & Fromont, 1988) , from New Zealand ( Table 3). These three species have large, paucispined acanthostyles similar to those of the new species. Moreover, C. (P.) basispinosa has subtylote to strongyloid tornotes that also approach the condition seen in Crella (P.) santacruzae Fernandez, Gastaldi, Thompson & Hajdu , sp. nov. Yet, the new species con be distinguished from C. (P.) basispinosa by the smaller length of its ectosomal acanthostyles (92–122 µm long vs. 250 µm, respectively), from C. (P.) fristedti by larger and stouter ectosomal diactines (393–576/7.6–12.2 µm vs. 212–242/3–4.5 µm, respectively), and from C. (P.) novaezealandiae by larger ectosomal diactines (393–576 µm vs. 240–300 µm, respectively) and chelae (22–31 µm vs. 11–20 µm, respectively). Furthermore, the immense distances between the areas of occurrence of these species make conspecificity unlikely.

| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heteroscleromorpha |
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Crella |