
Galapagetula annikae, Reygel, Patrick C., Willems, Wim R. & Artois, Tom J., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.200746 |
|
DOI |
https://doi.org/10.5281/zenodo.6189707 |
|
persistent identifier |
https://treatment.plazi.org/id/F24C880C-1F4D-9D21-6093-A548FE59FE54 |
|
treatment provided by |
Plazi |
|
scientific name |
Galapagetula annikae |
| status |
gen. nov. |
Galapagetula annikae View in CoL n. gen. n. sp.
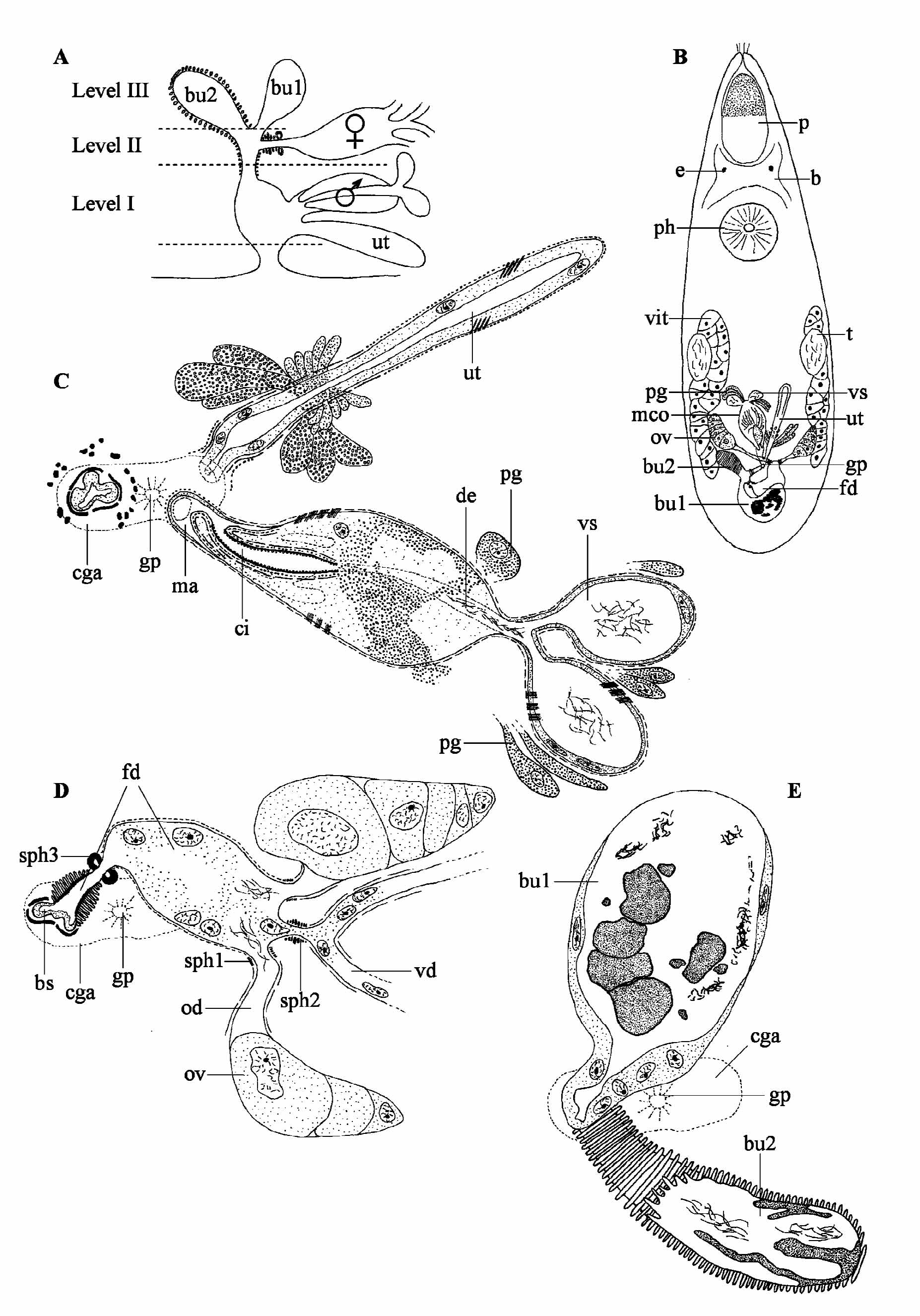
( Fig. 5 View FIGURE 5 )
Locality. Santa Cruz Island, station IX6 b, rock pools (type locality).
Material. One specimen studied alive and one serially-sectioned specimen designated holotype ( ZMUG 23251).
Etymology. The genus name is a contraction of “Galapagos”, and “ Getula ”, a resembling taxon. Species name dedicated to Annika Willems, daughter of the second author.
Diagnoses. Galapagetula n. gen. Koinocystididae with proboscis of the typical koinocystidid construction without proboscis juncture sphincter; pharynx without a mouth sphincter; testes at mid-body; copulatory organ pearshaped, distally cylindrical with a long, folded cirrus, armed with small spines; female duct proximally widened, functioning as seminal receptacle, distally highly muscular with strong sphincter; muscular bursa and resorptive bursa opening separately into the female duct.
Type species (by original monotypy): Galapagetula annikae n. sp., provisionally with the same diagnosis as the genus.
Description. Live specimen grey, spindle shaped, with two eyes. It is ± 1.2 mm long. The syncytial epidermis (± 10 µm thick) has a prominent basement membrane and contains numerous oviform rhabdites (2−4 µm long). They are absent from the anterior tip to half way along the proboscis. The animal is ciliated over its entire body surface (cilia ± 6 µm long), with some sensory bristles around the proboscis opening. The proboscis is about 1/5 of the body length. It has the characteristic traits of a koinocystidid proboscis (see Brunet 1972; Karling 1980). As far as could be derived from the sections, a proboscis juncture sphincter is absent. The pharynx is situated at 35% and shows no distinct pharynx mouth sphincter. Unfortunately, the material does not allow a detailed reconstruction of the pharynx and the proboscis. Observations indicate that the pharynx contains a fine-grained eosinophilic and coarse-grained basophilic secretion. The lumen is lined with a membranous epithelium.
Male and female gonads are paired and situated in the posterior half of the body. The oval testes lie dorso-laterally, at the beginning of the second half of the body. The spindle-shaped ovaries are situated at ± 75%. The short vitellaria lie ventro-laterally and extend from the level of the testes to the ovaries. The common genital pore is situated at ± 85% and opens into the globular common genital atrium, which is lined with a nucleated epithelium and surrounded by an outer longitudinal and a less prominent, inner circular muscle layer. It receives the uterus and the male and female genital system at three different “levels” (see schematic overview in Fig. 5 View FIGURE 5 B). The elongated uterus enters the common genital atrium rostro-ventrally, with the male copulatory organ situated just dorsally to the uterus and also opening into the atrium from the rostral side ( Fig. 5 View FIGURE 5 A: Level I; Fig. 5 View FIGURE 5 C). The female system enters the atrium from the dorsal side ( Fig. 5 View FIGURE 5 A: Level II; Fig. 5 View FIGURE 5 D). The most distal part of the female duct, which is highly muscular, receives two different bursal organs Fig. 5 View FIGURE 5 A: Level III; Fig. 5 View FIGURE 5 E).
The male genital system consists of an oval to pear-shaped copulatory organ with an armed cirrus. The seminal vesicles are almost globular and lined with a low, nucleated epithelium and surrounded by a spirally–running, almost circular muscle layer. Distally from the seminal vesicles, the vasa deferentia fuse just before entering the prostate vesicle, forming an ejaculatory duct that contains sperm in its proximal part. This duct runs centrally through the prostate vesicle and is surrounded by several prostate ducts with coarse-grained basophilic secretion. The cell bodies of the prostate glands are visible beside the seminal vesicles and the prostate vesicle ( Fig. 5 View FIGURE 5 C: pg). The exact course of the prostate ducts is difficult to discern, but there are indications that they enter the prostate vesicle a little distally from the ejaculatory duct. The muscle layer surrounding the seminal vesicles continues around the prostate vesicle. In contrast to observations on the live animal, the trajectory of the individual prostate ducts and the ejaculatory duct are not clearly visible on the sectioned material. However, they most probably enter the proximal part of the cirrus, which is situated in the most distal half of the male copulatory organ. The cirrus is lined with a thick basement membrane, which forms numerous minute, scale-like hooks (0.1 µm or less). The cirrus proper is surrounded by an inner layer of circular muscles and an outer layer of longitudinal muscles. It is folded, making its course very hard to reveal. At its distal end it forms a penis papilla-like structure, which protrudes into the male duct.
The elongated, almost spindle-shaped ovaries are connected to the female duct by short oviducts, which are surrounded by longitudinal muscles and are provided with a weak distal sphincter ( Fig. 5 View FIGURE 5 D: sph1). An epithelium could not be observed. Both vitelloducts fuse into a very short single duct, which is provided with a strong sphincter, just before entering the female duct in between the two oviducts (see Fig. 5 View FIGURE 5 D: sph 2). The most proximal part of the female duct is surrounded by circular muscles. As for the oviducts, an epithelium cannot be seen. The female duct is clearly widened and contains many vitellocytes and some sperm close to the oviducts, functioning as a seminal receptacle. More distally it narrows and can be closed by a very strong sphincter (see Fig. 5 View FIGURE 5 D: sph 3). From this point the female duct is lined with pseudocuticle (dense and thickened basement membrane not covered by epithelium) and surrounded by a very thick circular muscle layer, before entering the muscular bursal stalk. The bursal system consists of a muscular bursal stalk, which proximally ends in a large bursa ( Fig. 5 View FIGURE 5 A, B, E: bu1). The bursa is lined with a nucleated epithelium and filled with a mass of different density, wherein several nodular structures and clusters of sperm can be recognised. The bursal stalk also forms a very muscular diverticulum which contains some sperm and is lined with a thick irregularly-folded pseudocuticle: the accessory bursa ( Fig. 5 View FIGURE 5 A, B, E: bu2). It is surrounded by strong circular muscles that stain rather weakly compared to those of the female duct.
The uterus is lined with a nucleated epithelium and surrounded by spirally-running muscle fibres, distally continuing as longitudinal muscles. At the distal end a second outer circular muscle layer is present around the stalk. Two types of uterus glands are present: fine-grained eosinophilic ones proximally and coarse-grained basophilic ones distally.
Discussion. The new species shows all features mentioned in the diagnosis of the taxon Koinocystididae , given by Karling (1980). However, the specific combination of features shown by this species is unique, and does not allow its placement in any existing genus. The only other taxa without a proboscis juncture sphincter within Koinocystididae are Axiutelga Karling, 1980 , Getula Karling, 1980 , Itaipusina Karling, 1980 , Leguta Karling, 1980 , Parautelga Karling, 1964 , Pontaralia Mack-Fira, 1968 , Rhinolasius Marcus, 1951 and Utelga . G. annikae clearly differs from all these taxa, and the other koinocystidids, in the presence of the accessory bursa. In a number of species (e.g. Itaipusa ruffinjonesi ) the bursal stalk is enlarged towards the entrance of the seminal receptacle, an enlargement called “receptacle bursa” by Karling (1980). The position of the accessory bursa in G. annikae is, however, clearly different from that of the “receptacle bursa” described by Karling (1980).
The fact that the epidermis is syncytial is not uncommon within the Koinocystididae . For a discussion on this topic, see the discussion on Itaipusa .
A cirrus provided with uniform, small spines is also rather exceptional within Koinocystididae . Either the cirrus is unarmed, or the spines are not uniform, or additional large hooks are present. Basically, only in Axiutelga does the cirrus show uniform, albeit much larger spines than in G. annikae (7−10 µm and 0.1 µm respectively). The unique construction of the female system combined with a uniformly-spined cirrus warrants the erection of a new genus for G. annikae .
| ZMUG |
Zoologisches Museum der Universitat Gottingen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Kalyptorhynchia |
|
Family |
|
|
Genus |