
Hadrocryptolarynx major Haran, 2023
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.877.2151 |
|
publication LSID |
lsid:zoobank.org:pub:65E8C3F7-5EA4-4013-A09E-37C96B4929FB |
|
DOI |
https://doi.org/10.5281/zenodo.8113786 |
|
persistent identifier |
https://treatment.plazi.org/id/55B2B09A-2A4F-4D0D-996F-8E93E874C0E9 |
|
taxon LSID |
lsid:zoobank.org:act:55B2B09A-2A4F-4D0D-996F-8E93E874C0E9 |
|
treatment provided by |
Felipe |
|
scientific name |
Hadrocryptolarynx major Haran |
| status |
gen. et sp. nov. |
25. Hadrocryptolarynx major Haran gen. et sp. nov.
urn:lsid:zoobank.org:act:55B2B09A-2A4F-4D0D-996F-8E93E874C0E9
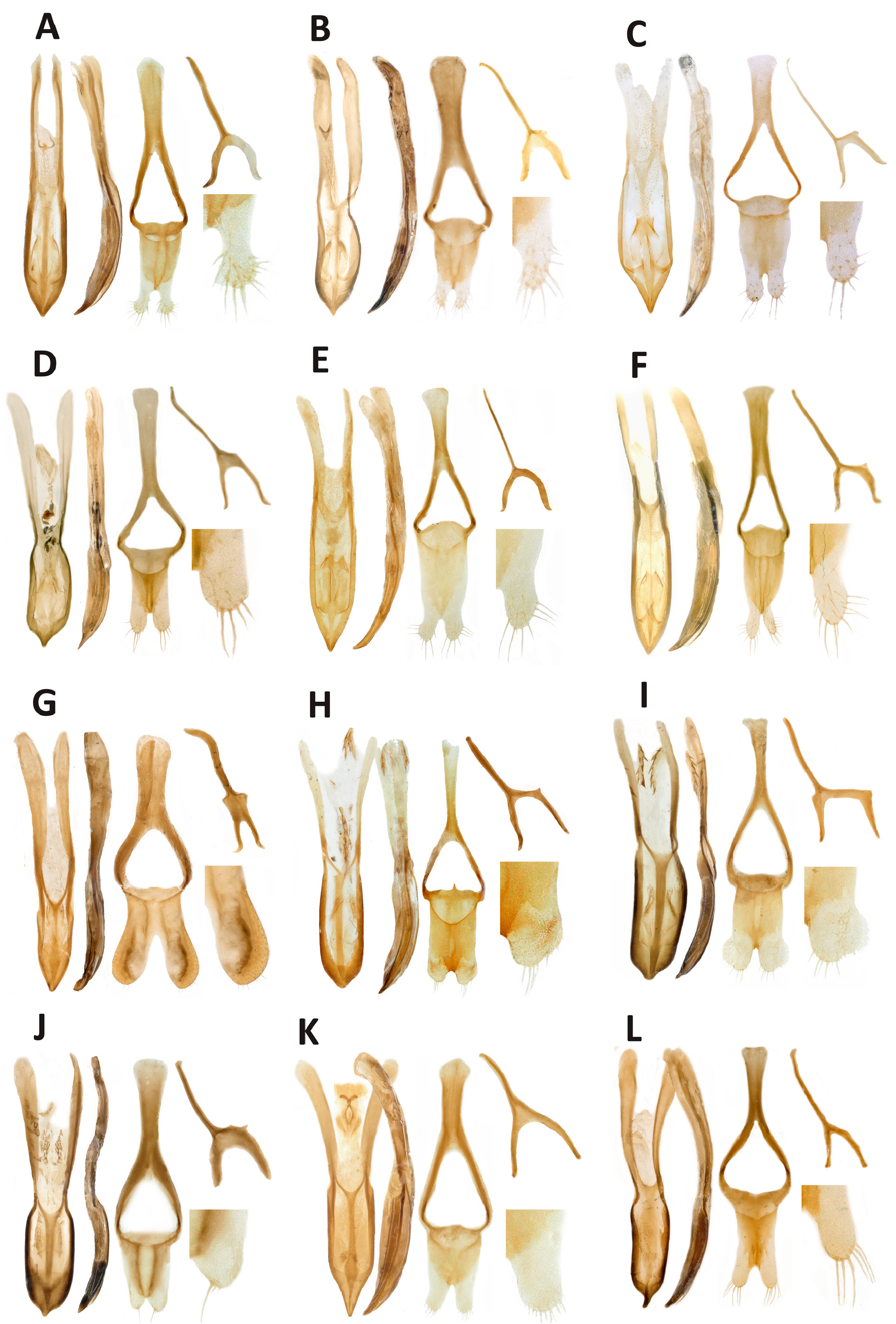
Figs 1X View Fig , 2X View Fig , 3X View Fig , 4X View Fig , 5X View Fig , 7M–P View Fig , 8F View Fig
Differential diagnosis
Hadrocryptolarynx major gen. et sp. nov. is at present the only species in its genus. See diagnostic characters in key to genera for differential diagnosis with species of Cryptolarynx .
Etymology
The species name major refers to the large body size of some specimens of the species, seemingly the largest among the Cryptolaryngini . The specific epithet is an adjective in the masculine form.
Material examined
Holotype REPUBLIC OF SOUTH AFRICA • ♂; “REPUBLIC OF SOUTH AFRICA. Western Cape Province, Klawer [13 km S]. 26.vii.2019. J. Haran leg.” “ 31.902° S 18.630° E. at base of Oxalis luteola . JHAR02464_0101. Cirad-CBGP coll.” “Holotype. Hadrocryptolarynx major . Haran 2023”; SAMC. GoogleMaps
Paratypes REPUBLIC OF SOUTH AFRICA – Western Cape • 1 ♂, 1 ♀; same collection data as for holotype; CBGP GoogleMaps • 1 ♂, 1 ♀; same collection data as for holotype; NHMUK GoogleMaps • 1 ♂; same collection data as for holotype; MNHN GoogleMaps • 4 specs (preserved in ethanol); same collection data as for holotype; CBGP GoogleMaps • 10 specs; Klawer 13 km S; 31.902° S, 18.630° E; 26 Jul. 2019; J. Haran leg.; at base of Oxalis luteola ; JHAR02486; CBGP GoogleMaps • 1 ♂, 2 specs; Velddrif , 32.722° S, 18.194° E; 27 Jul. 2019; J. Haran leg.; near stands of Oxalis pes-caprae ; JHAR02482; CBGP GoogleMaps • 1 ♂, 1 ♀; Worcester ; 33.650° S, 19.417° E; 23 Sep. 1973; L. Schulze leg.; collected from flowers in Karoo garden; E-Y:148; TMSA GoogleMaps • 1 ♀; 10 km WSW of Uitvlugt, Zandkraal Farm ; 31.700° S, 18.767° E; 12 Sep. 1987; S. Endrödy-Younga leg.; dry coarse-sandy flat; E-Y:2490; TMSA GoogleMaps • 2 ♂♂, 1 ♀; 5 km S of Vanrhynsdorp ; 31.600° S, 18.733° E; 26 Sep. 1973; L. Schulze leg.; collected from Aizoaceae flowers; E-Y:162; TMSA GoogleMaps • 1 ♂; Vanrhynsdorp S, Farm Wiedouw 309; 31.733° S, 18.783° E; 20–23 Sep. 1982; S. Louw leg.; NMBH 89711; SANC GoogleMaps • 1 ♀; Namaqualand, Koekenaap dunes; 31.533° S, 18.233° E; 23 Sep. 1994; S. Endrödy-Younga and C.L. Bellamy leg.; pitfall traps 3 days; red sand dunes with patchy dense bushy vegetation; E-Y:3032; TMSA GoogleMaps .
Description (♂)
MEASUREMENTS. Body elongate, length 2.0–6.0 mm.
COLOUR AND VESTITURE. Body integument black, antennae, tibiae and tarsi reddish. Dorsal vestiture (pronotum + elytra) consisting of overlapping, recumbent, parallel-sided or bilaterally convex clothing scales, each 2–5 × as long as wide, truncate at apex, not aligned on interstriae, concealing integument. Colour of scales black, dark brown to pale brown, grey or white; darker scales usually concentrated medially over pronotum and on elytral interstriae 1–3; paler scales concentrated laterally and on pronotum and elytra, usually forming a transverse spot near middle of interstriae 1–3. Scales arising from strial punctures not different from those of rest of vestiture.
HEAD. Forehead slightly wider than epifrons near antennal insertions, scales recumbent. Eyes convex, in dorsal view distinctly exceeding outline of head, surrounded by a ring of short scales, on forehead directed towards occiput; distance between eye and scrobe less than width of an eye. Epifrons flat, as wide as forehead, epifrontal scales suberect and orientated laterad, towards scrobes, overlapping. Frons with 3 pairs of long erect lateral setae. Epistome without median seta. Antennae with funicle segment 1 longer than wide, as long as 2 and 3 combined, 2–4 subcylindrical, slightly angular on inside, 5–7 globular or moderately elongate.
THORAX. Pronotum moderately transverse (W:L ratio 1.15), widest near midlength, sides arcuate; apex and base subequal in width; integument densely punctate, spaces between punctures narrow, dull, surface regularly convex.
ELYTRA. Broadly ovate (W:L ratio 0.76), sides arcuate, widest near midlength.
LEGS. Slender. Tibiae with apical mucro, inner margin bisinuate; protibiae curved in apical half; mesotibiae curved in apical third; metatibiae straight. Protarsi with segments 1 and 2 isodiametric; meso and metatarsi with segments 1 and 2 2 × as long as wide.
ABDOMEN. Ventrites 1–4 with pale, recumbent, plumose scales intermixed with white suberect setae; ventrite 5 with a few scales near base, elsewhere setose only; ventrites 1–2 slightly concave medially, 5 convex medially.
TERMINALIA. Body of penis elongate (W:L ratio 0.3), as long as temones, acuminate at apex, in profile downcurved in apical third. Tectum narrow but distinct; endophallus with copulatory sclerite divided into two symmetrical, elongate structures. Parameroid lobes of dorsal plate of tegmen fused and jointly evenly rounded at apex, devoid of seta. Spiculum gastrale asymmetrical; divergence of basal arms V-shaped.
Sexual dimorphism
The sexes can be distinguished by the shape of the elytra (elongate and widest near midlength in male, shorter and widest at base in female) and by ventrites 1–2 (concave in male, convex in female), and half of females examined bear a series of long, suberect setae on interstriae 1–3–5–7.
Life history
Adults of Hadrocryptolarynx major gen. et sp. nov. were at several localities found in dense stands of Oxalis luteola , most specimens collected from the base of the plants. As in Cryptolarynx , some specimens were observed to form small holes in the soil, below the leaves of various surrounding plants ( Fig. 7O View Fig ). The weevils are active during the day on open fields but may also climb onto vegetation. Adults were collected between July and September.
Distribution
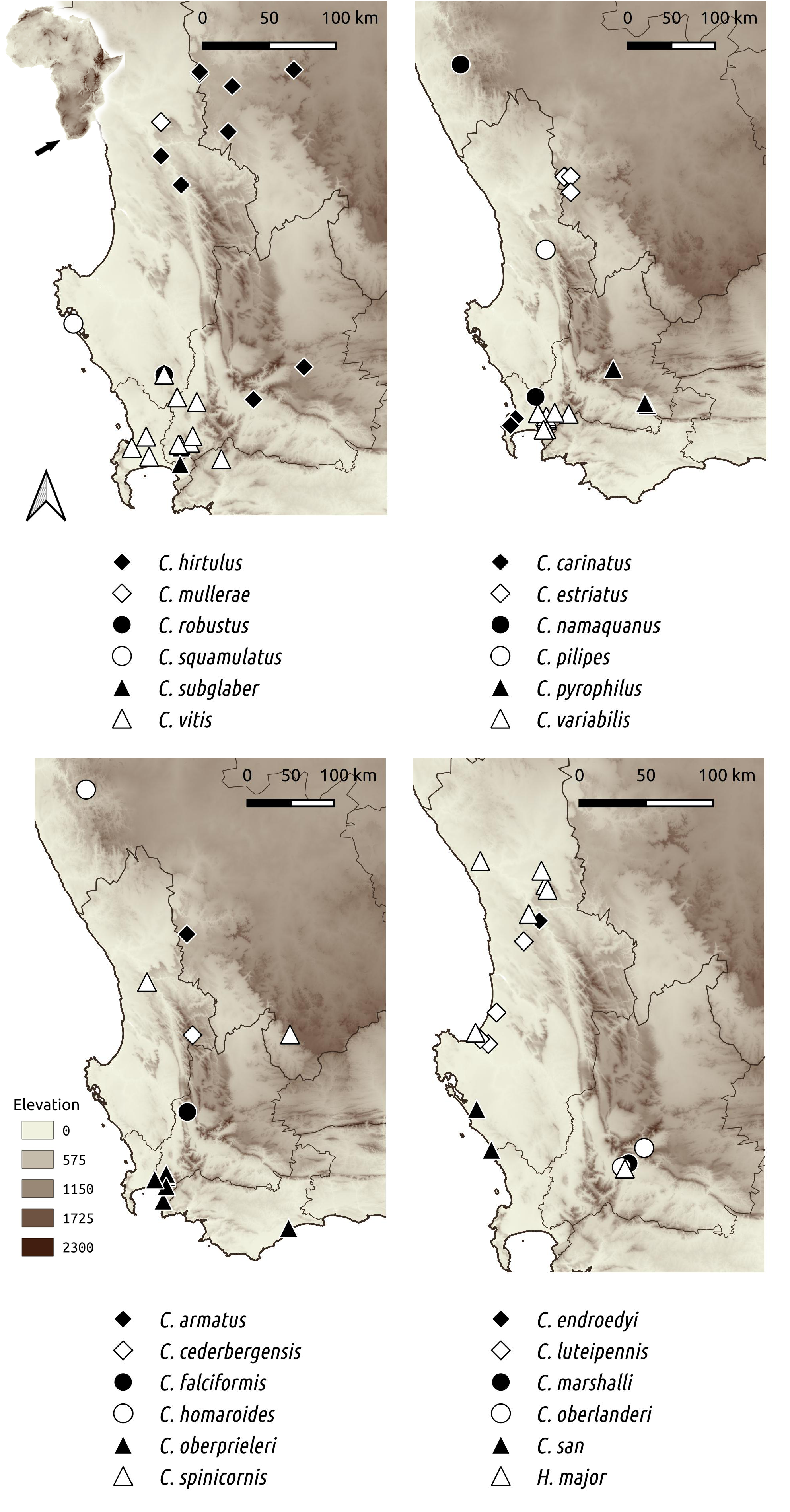
Hadrocryptolarynx major gen. et sp. nov. was mostly collected in the northwestern regions of the Western Cape province, from Koekenaap to Velddrif and inland to Vanrhynsdorp and Worcester ( Fig. 13 View Fig ).
Remarks
Specimens of Hadrocryptolarynx major gen. et sp. nov. were collected in sympatry with those of C. spinicornis sp. nov. Amplification of the standard mitochondrial barcode fragment failed repeatedly for this species, and the fragments of the only two specimens that successfully amplified (JHAR02464_0101 and JHAR02486_0101) yielded five regions with large nucleotide deletions. By contrast, all remaining nucleotides aligned well with those of the various species of Cryptolarynx sequenced and showed no traces of stop codons. This fragment is probably of pseudogene origin caused by a mismatch between the primers and the actual mitochondrial copy of the COI fragment.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Brachycerinae |
|
Tribe |
Cryptolaryngini |
|
Genus |