
Capitina Watson, 1934
|
publication ID |
https://doi.org/ 10.5733/afin.051.0101 |
|
persistent identifier |
https://treatment.plazi.org/id/110B87C2-FFEF-FFA4-D7A0-FCC2FE3CFEEC |
|
treatment provided by |
Felipe |
|
scientific name |
Capitina Watson, 1934 |
| status |
|
Genus Capitina Watson, 1934 View in CoL View at ENA
Capitina: Watson 1934: 153 . Type species: Helix schärfiae [= schaerfiae ] Pfeiffer, 1861, by original designation.
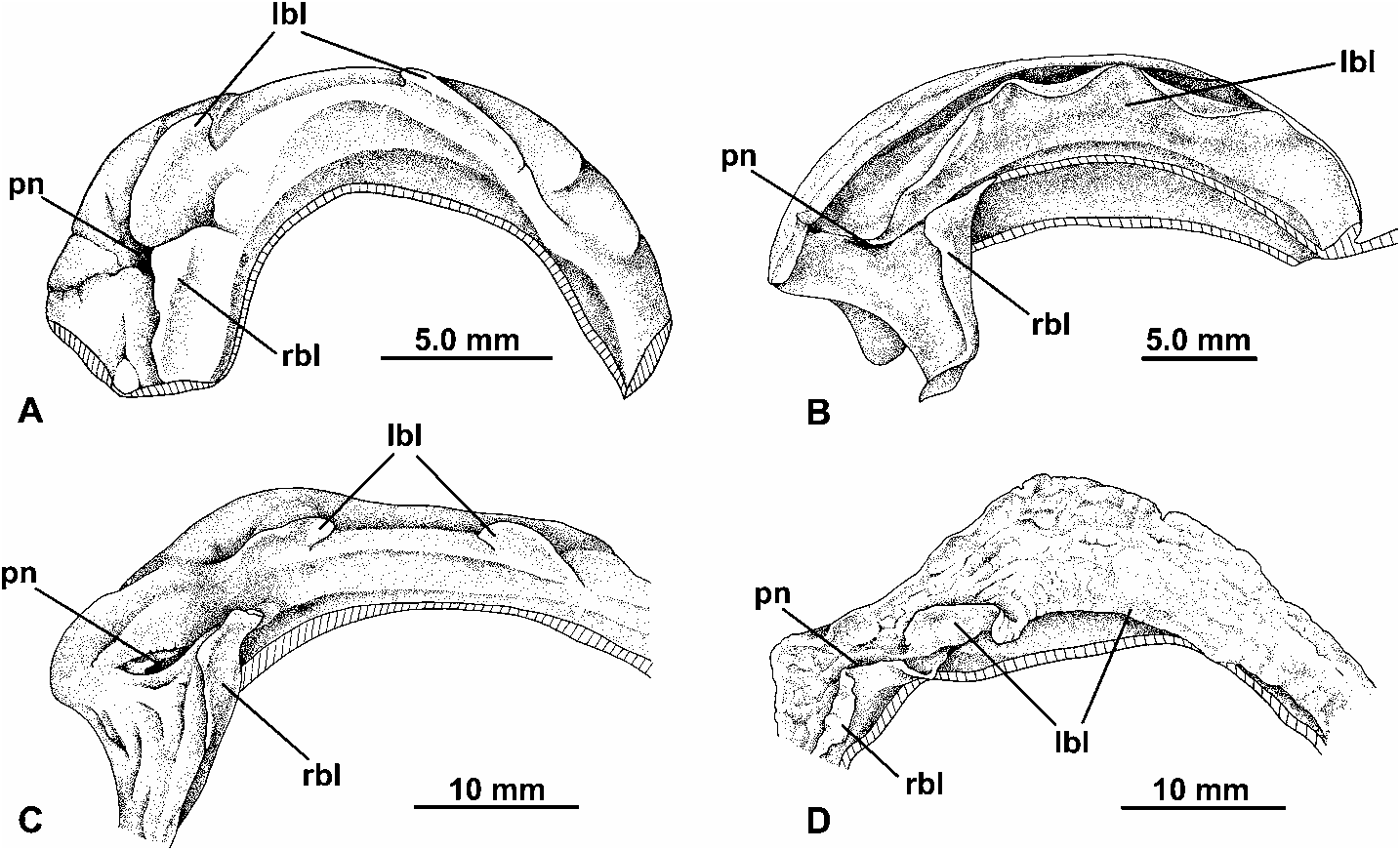
Diagnosis: Shell of moderate size (adult diameter up to 34 mm), discoidal to globose-lenticular, usually with distinct spiral colour pattern; sculptured by anastomosing axial riblets producing a somewhat pitted or wrinkled sculpture; outer lip slightly thickened and without periostracal fringe; peristome interrupted in parietal region; umbilicus open, width moderate; protoconch of 1.25–1.5 whorls, with close-set axial riblets ( Fig. 4C View Fig ) and often with traces of spiral sculpture, diameter 4.0–7.0 mm. Radula possessing rachidian tooth; no clear distinction between lateral and marginal series and no one tooth noticeably larger than the others. Mantle lobe to left of pneumostome undivided. Penis very long, situated to left of retractor muscle of right optic tentacle; epiphallus short (<25 % of penis length); epiphallus and lower vas deferens connected to penis by connective tissue web; vagina very long; oviduct caecum absent. Distal part of suprapedal gland sinuous but not convoluted, lacking a swollen terminal vesicle.
External anatomy ( Fig. 76 View Fig ): Left body lobe of mantle undivided, forming a continuous skirt over neck region ( Fig. 6B View Fig ); skin texture finely to moderately granular; labial palps present but not large; optic and inferior tentacles with a ventrally distended apical bulb; tail region of foot relatively short; genital pore ventral and just posterior to right optic tentacle; lung venation strongly delineated by black pigmentation.
Radula ( Fig. 73 View Fig ): Formula 1+(20–32); length up to 15.5 mm, with up to 84 V-shaped transverse rows of teeth, 5.0–6.6 rows/mm in adult; no clear distinction between lateral and marginal series. Rachidian present, with a relatively short, broad cusp, one quarter to one third length of base-plate; remaining teeth (laterals and marginals), elongate and slender, progressively (slowly) increasing in size, those in middle of lateromarginal series largest, then decreasing in size somewhat more rapidly toward edge of radula, the outermost tooth being minute; inner teeth claw-like, with a relatively short, weakly curved, broadly acuminate cusp; larger teeth in middle of lateromarginal series with a flatter, more elongate cusp, but lacking an enlarged quadrate base-plate; those peripheral to this becoming spathulate, with a rounded tip and anteriorly acuminate base-plate.
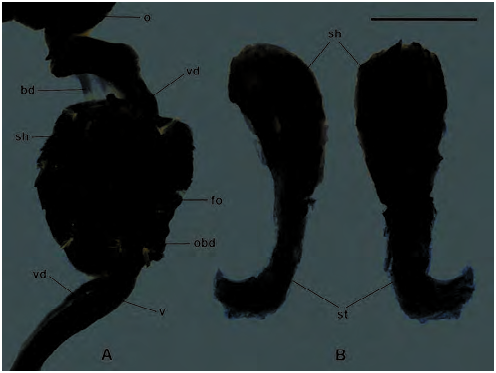
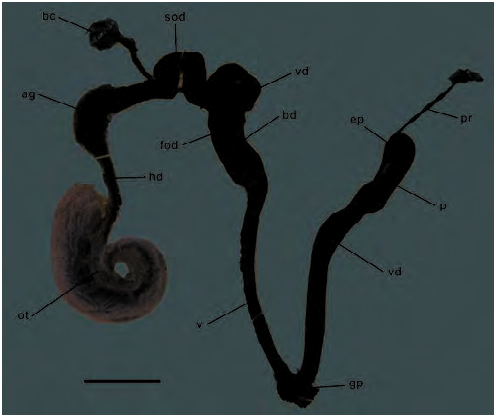
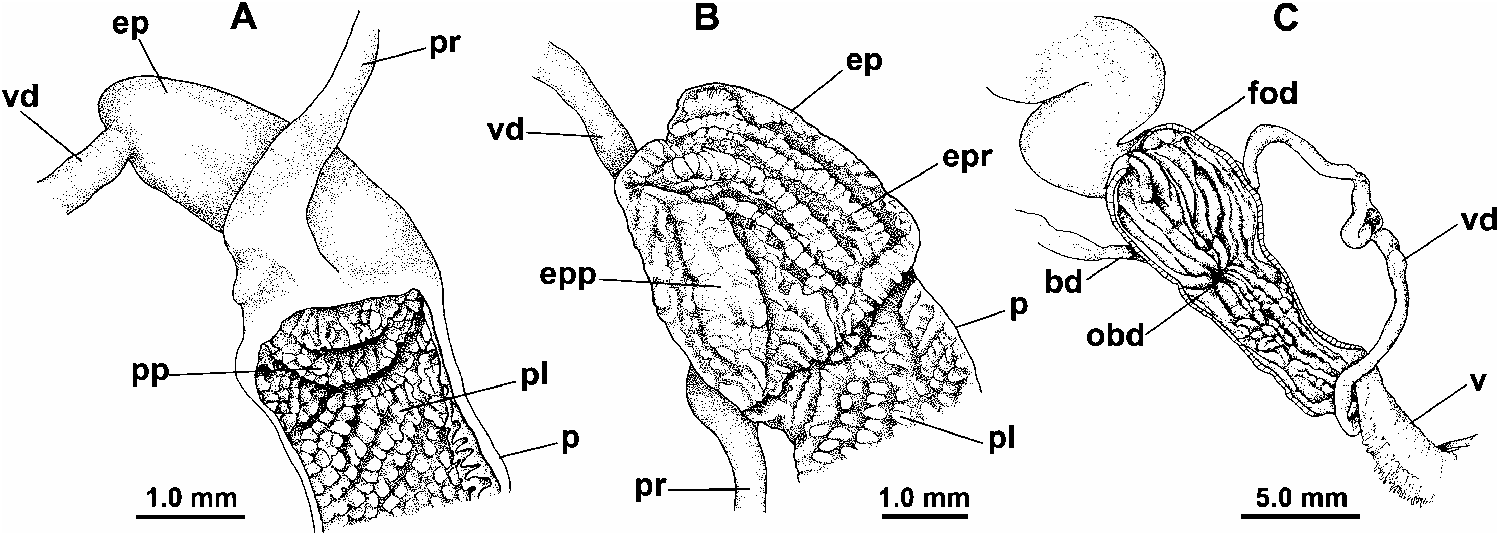
Distal genitalia ( Figs 9C View Fig , 74 View Fig , 75 View Fig ): Penis very long and slender, often sinuously folded in situ; its wall muscular, particularly towards base; internal lumen lined by papillate epithelium ( Fig. 75A View Fig ), papillae close-set and relatively coarse, somewhat larger toward penis base. Apex of penis with small, rounded penis papilla, through which epiphallus lumen connects with penis lumen ( Fig. 75A View Fig ); penis papilla itself covered with micro-papillae similar to those lining the penis lumen. Epiphallus short, <25 % of penis length; joins penis at U-shaped bend where penial retractor muscle inserts; interior with longitudinal ridges lining outer wall, lying opposite a broad, rugose, tongue-shaped pilaster on inner wall (adjacent to penis) ( Fig. 75B View Fig ). Vas deferens runs alongside penis, vagina and free oviduct, broader and thrown into a series of bends in the latter region, before it fuses with spermoviduct.A web of connective tissue joins lower vas deferens and epiphallus to penis.
Vagina very long and slender with muscular wall; attached laterally to body wall by a well-developed series of muscle fibres; merges with free oviduct at origin of bursa copulatrix duct; interior wall of vagina with weakly crenulate, longitudinal ridges. Free oviduct expanded and thick-walled (site of spermatophore receipt), lined with smoother longitudinal ridges ( Fig. 75C View Fig ). Oviduct caecum absent. Bursa copulatrix duct long and slender, but not as slender as that of Natalina , running beside spermoviduct rather than within its coils; bursa copulatrix thin-walled, more or less ovate, situated posterior to pericardium. Spermoviduct relatively short, the lower half convoluted; hermaphrodite duct highly convoluted; ovotestis comprising a loose assemblage of slender acini, embedded in apical whorl of digestive gland.
Spermatophore ( Figs 10A View Fig , 76 View Fig ): A single allospermatophore was found in a specimen of Capitina calcicola sp. n. collected in early October. It was in good condition, suggesting recent copulation and thus that mating occurs in spring, at least. Spermatophore tadpole-shaped (straightened length 8.2 mm) and situated in swollen, thick-walled, lower portion of free oviduct. Head of spermatophore foremost, lying close to origin of free oviduct from spermoviduct, tail curving downward into opening of bursa duct. Spermatophore head pale orange-pink and hard, with numerous irregular, sometimes sinuous, longitudinal ridges; underside of the head concave, corresponding to tongue-like pilaster in epiphallus; internal contents of spermatophore head whitish; tail considerably thinner and without pink coloration or internal contents. No obvious vent evident, but central portion of concave underside of head thin-walled and possibly representing rupture site.
Notes: The phylogenetic analysis of morphological characters and DNA sequence data published by Moussalli et al. (2009) has shown that Capitina represents a highly distinctive lineage within Natalina s. l. As a result, we have proposed that the taxon be ranked as a genus in its own right. In terms of shell features, Capitina is characterised by the brown spiral lines on the adapical surface, wrinkled sculpture and thickened apertural lip of the adult. In addition, Capitina exhibits a number of distinct anatomical character states associated with the mantle edge, radula and distal genitalia which clearly distinguish it from Natalina and Afrorhytida .
The distribution of the genus is confined to the southern part of the south-western Cape and appears not to overlap with that of either Natalina 2 or Afrorhytida . It is the only genus of relatively large rhytidids known from the south-western Cape, west of 20°E. This, combined with the basal split between Capitina and ( Natalina + Afrorhytida ) ( Moussalli et al. 2009) suggests that Capitina stems from an early divergence event which divided the ancestral Natalina s. l. stock into western and eastern lineages. Using a conservative mutation rate of 5 % pairwise, mtDNA sequence data places this basal split in the mid-Miocene ( Moussalli et al. 2009). However, Spencer et al. (2006) suggested an even slower rate for COI evolution within the New Zealand rhytidid radiation, and there is thus a possibility that the initial split within Natalina s.l. may date from as far back as the early Miocene ( Moussalli et al. 2009). It is interesting to note here in terms of an ancestral reconstruction, that the radula of Capitina is closer to that of the Australian Strangesta Iredale, 1933 (Smith 1979) and New Zealand Schizoglossa Hedley, 1892 ( Powell 1930) than it is to either Natalina or Afrorhytida . However, such similarities may be diet-related homoplasies and need to be interpreted with caution.
When proposing Capitina as a new taxon, Watson (1934) included in it only ‘ Natalina ’ schaerfiae , but noted the existence of light coloured specimens which Layard (in Benson 1864) had suggested were simply pale varieties or bleached specimens of the same species.Watson, however, observed additional differences in sculpture, shell proportions and protoconch size, and speculated that these specimens may ultimately prove to belong to a second and unnamed taxon.With considerably more material available and accurate locality data we are able to confirm Watson’s observations and here describe this paler lineage as a new species.
The Agulhas Plain, to which this new species is limited, has experienced repeated Neogene marine transgressions of up to 200 m, with sea levels retreating to present levels only as recently as 2 Mya ( Siesser & Dingle 1981; Linder 2003). Such transgressions may have constituted an effective isolating mechanism, restricting ancestral Capitina populations to refugial foci in the Riviersonderendberge and at higher elevations
2 Juvenile specimens of Trigonephrus species occurring in the Agulhas area can deceptively resemble juvenile specimens of Natalina .
on the Agulhas Plain, facilitating divergence within the ancestral stock. This is consistent with a Late Pliocene estimate for the divergence of the two lineages based on COI sequence data (unpubl. data).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |