
Heterochterus timmsii Evans, 1971
|
publication ID |
https://doi.org/ 10.37520/aemnp.2020.003 |
|
publication LSID |
lsid:zoobank.org:pub:085DDB9B-0127-4EF6-9B32-826C0FB1219E |
|
DOI |
https://doi.org/10.5281/zenodo.4469547 |
|
persistent identifier |
https://treatment.plazi.org/id/43368780-FA48-1879-BAAD-FCF2FECBFB76 |
|
treatment provided by |
Tatiana |
|
scientific name |
Heterochterus timmsii Evans, 1971 |
| status |
|
Ƭ Heterochterus timmsii Evans, 1971
Heterochterus timmsii Evans, 1971: 149– 150 (original description, illustration). HOLOTYPE:?sex, wing, Australia: Queensland: ‘ Mt Crosby, coll. B. V. Timms, presented by Department of Entomology , University of Queensland, Q.M. F6473’ (QMBA).
Heterochterus timmsii : JELL (2004): 65 (redescription, illustrations).
Distribution. Australia: Queensland (Ipswich Coal Measures: Late Triassic) ( EVANS 1971, JELL 2004).
Discussion
The first described species of velvety shore bugs was the widely distributed Acanthia marginata Latreille, 1804, discovered in southern France, and soon after accommodated in its own new genus, Ochterus Latreille, 1807 ,
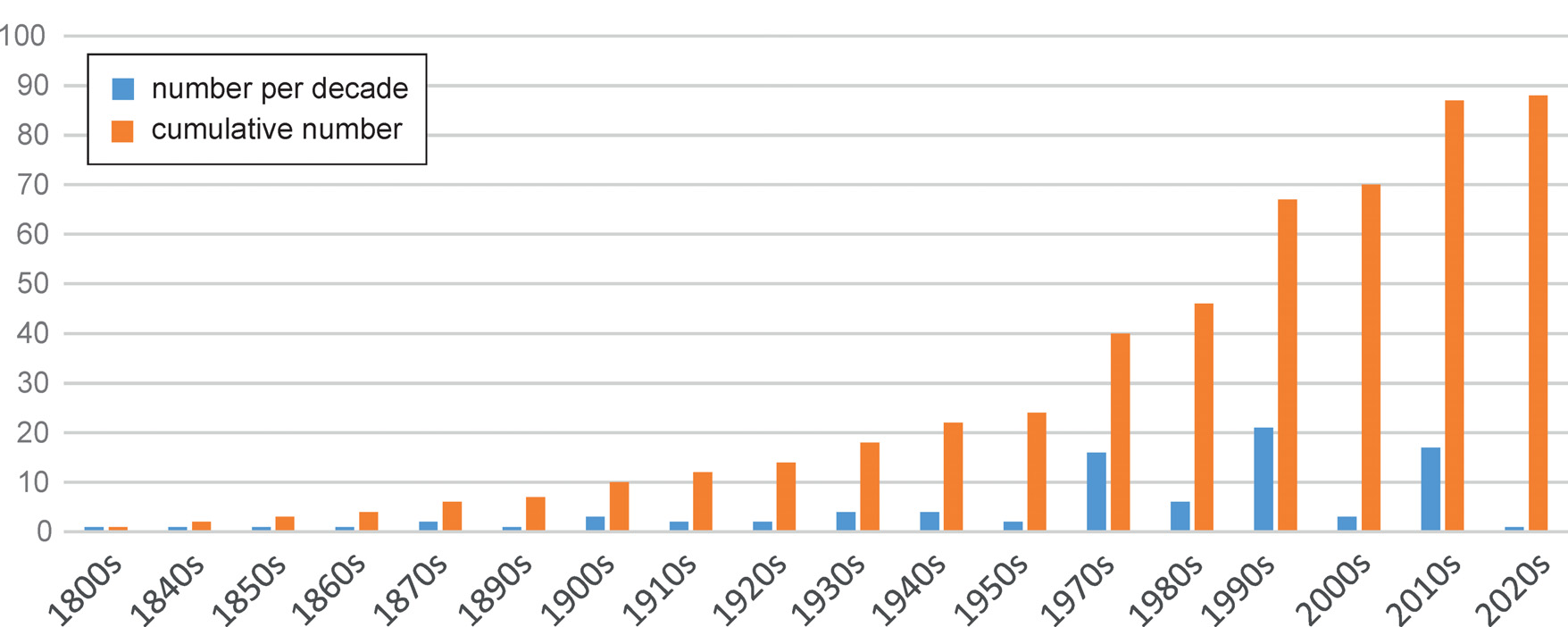
and new family, Pelogonida Leach, 1815 (based on an unnecessary new replacement name Pelogonus Latreille, 1809). Additional two taxa were described by GUÉRIN- MÉ- NEVILLE (1843), Pelogonus indicus (junior synonym of O. marginatus marginatus ) from India, and the first American species, Pelogonus perboscii from Mexico. Until the end of 19th century, altogether eleven species of Ochteridae were described, seven of which still remain valid. Among them were also two species which were in the future designated as types of new genera, Ocyochterus Drake & Gómez-Menor, 1954 (Pelogonus victor Bolívar, 1879) and Megochterus Jaczewski, 1934 (Pelogonus nasutus Montandon, 1898 ). Between 1901 and 1970, 17 additional species of Ochteridae were described, mostly from the American continent where important contributions were published by CHAMPION (1901), BARBER (1913), HUNGERFORD (1927), and especially the revision by SCHELL (1943a,b). In the Old World, only five species were described in this period ( HORVÁTH 1913; JACZEWSKI 1934, 1935, 1938; MANCINI 1939). SINGH- PRUTHI (1925) first described and illustrated male genitalia including the right paramere of Ochterus marginatus , a character soon applied in ochterid taxonomy by HUNGERFORD (1927) and JACZEWSKI (1934). Big progress in describing new Ochteridae started in 1970s with three revisional papers by Nicolas A. Kormilev for Australian, Oriental and Afrotropical taxa ( KORMILEV 1971, 1972, 1973), largely based on the structure of the right paramere. Soon followed the papers of NIESER (1975) ( Suriname), POLHEMUS & POLHEMUS (1976) (SW USA), RIEGER (1977) ( Australia, Philippines), and GAPUD & SAN VALENTIN (1977) ( Philippines). Victor P. Gapud continued his work on the fauna of Philippines also in the following three decades ( GAPUD 1981, 1995, 2003). In the same time, Martin Baehr, a renowned German coleopterist specialised in Carabidae (Coleoptera) , revised Ochteridae of the Australian Region in a series of four papers ( BAEHR 1989, 1990a,b,c). In 1990s, Nico Nieser and Ping-ping Chen described seven additional species mostly from Malesia, but also from southern China and South America ( NIESER & CHEN 1992, 1999). Finally, Dan A. Polhemus started describing new Ochteridae with Ochterus seychellensis in 1992 ( POLHEMUS 1992a), and continued, together with his father John T. Polhemus, with series of revisional papers on Ochterus of Mauritius ( POLHEMUS & POLHEMUS 2008), Malaysia and the Oriental part of Indonesia (2012, 2015), North and Central America ( POLHEMUS & POLHEMUS 2016), as well as a revision of the genus Ocyochterus with the description of one new species ( POLHEMUS & POLHEMUS 2014). Since 1971, 64 species of Ochteridae were described, nearly three times the number described before this date (see Table 1 and Fig. 25 View Fig ). The most prolific authors were D. A. and J. T. Polhemus, who described together 17 species of Ochteridae , followed by M. Baehr with 16 species and subspecies, N. Nieser with 9 species, and N. A. Kormilev and V. P. Gapud each with 8 species (cf. Table 2). The species inventory of Ochteridae is certainly far from complete, as it is documented by mentions of a probable new species of Ochterus from Palawan ( GAPUD 2003) and several undescribed species from New Guinea ( POLHEMUS & POLHEMUS 1998, 2012), and Oriental Region, as well as due to the probable polyphyly of O. marginatus ( CHEN et al. 2005). Also our unpublished results suggest existence of additional undescribed species, especially in the Oriental Region, the Afrotropical Region and Madagascar.
The extant members of the family Ochteridae are distributed in the tropical and subtropical regions of all continents. The present lack of published records from Lesser Sunda and Maluku Islands ( Indonesia) is probably due to sampling bias, as we have at hand unidentified material from Maluku, superficially resembling O. nicobarensis (ZJPC). Further Ochteridae inhabit temperate zones of Australia and Tasmania ( ANDERSEN & WEIR 2004), East Palearctic (north to the northernmost Japanese island, Hokkaido, and to Kunashir, the southernmost island of Kuriles) ( POLHEMUS 1995 a, KANYUKOVA 2006, HAYASHI et al. 2016, HAYASHI & MIYAMOTO 2018) and Eastern Nearctic Region (north to Massachusetts, Michigan, Minnesota and Ontario ( POLHEMUS & POLHEMUS 1988a, 2016; MAW et al. 2000). On the contrary, Ochteridae are missing in temperate Europe, continental regions of Central Asia ( POLHEMUS 1995 a, KANYUKOVA 2006), from the Appalachian Mountains and the Ozark Plateau in the eastern part, as well as from most of the western part of USA ( POLHEMUS & POLHEMUS 1988a, 2016), from New Zealand, and from temperate and subtropical South America (the most southern species being O. foersteri reaching the Misiones province of Argentina – MELO 2015, BACHMANN & MAZZUCCONI 2017). The zoogeographical pattern of Gelastocoridae resembles that of Ochteridae , with three exceptions: i) Gelastocoridae in South America occur further south, into central Chile; ii) in the Eastern Palaearctic they extend northward only to central China and the southern Japanese islands of Kyushu and Shikoku; and iii) they are completely absent in the Western Palaearctic (except one record from coast of Sinai, Egypt) (e.g., TODD 1955, 1961; POLHEMUS & POLHEMUS 1988b; POLHEMUS 1995b; ANDERSEN & WEIR 2004; CHEN et al. 2005; KMENT & JINDRA 2008; FAÚNDEZ & ASHWORTH 2015; FAÚNDEZ & CARVAJAL 2017; NIESER et al. 2020).
Concerning the biodiversity pattern, we can recognize two distinct biodiversity centres, the Australasian Region with 29 species-group taxa and the Neotropical Region with 28 species-group taxa. Within the Neotropical Region, most of these taxa (i.e. 17) are known from Central America, while only 14 species and subspecies are known from South America (see Table 2). This situation may be influenced by the existence of the recent excellent revision of Central American fauna by POLHEMUS & POLHEMUS (2016) and future research will elucidate the precise number of species actually occurring in South America. In the Caribbean, so far only one endemic species is known ( O. hungerfordi from Cuba) and two widely distributed species shared with the continent ( O. aeneifrons aeneifrons and O. perbosci ), which is in contrast with high number of island endemics occurring in Malesia and islands of the Indian Ocean. In the Australian Region, 5 species occur in Sulawesi (including the widely distributed O. marginatus marginatus and O. noualhieri shared with Sundaland), 11 species in New Guinea, Vanuatu and Solomon Islands, 13 species and subspecies in Australia, and 2 species in New Caledonia (one of them endemic). The Oriental Region seems also species rich with 22 species-group taxa, but only 5 species are so far known in its western (mostly continental) part north of the border of Malaysia; 8 species occur in Sundaland and 11 species and subspecies are confined to the Philippines. The Afrotropical Region harbours 8 species, 6 of them occurring on the continent and 4 on the adjacent islands; O. caffer and O. papaceki sp. nov. occur in both the African mainland and islands ( Madagascar and Socotra, respectively), while Mauritius and Seychelles each harbour a single endemic species. The fauna of the Nearctic Region includes five species (with O. rotundus and O. perbosci confined to its southernmost regions and shared with Neotropical Region) ( POLHEMUS & POLHEMUS 2016). The least species rich is fauna of the Palaearctic Region, including only three described species, the widely distributed O. marginatus marginatus , O. strigicollis endemic to Israel, and O. breviculus confined to the transitional zone between the Palaearctic and the Oriental Region in Yunnan and Xizang (Tibet) in southern China ( NIESER & CHEN 1992, POLHEMUS 1995a). In alternative zoogeographical regionalisation, Malesia (including the Malay Peninsula, Sundaland, Philippines, Wallacea, New Guinea and Solomon Islands), would harbour 33 species and subspecies – see CHEN et al. 2005, ZETTEL & LANE 2010. POLHEMUS & POLHEMUS 2012). In contrast to the New World, we must stress the importance of island endemism for the species diversity in Old World Ochterus – among 55 Old World species, 33 (60 %) of them are confined to islands. Interestingly, also Gelastocoridae also share a similar pattern of species richness, with centres of biodiversity in Neotropical and Australian Region; the Oriental Region harbours only 9 species, 3 of them extending to Eastern Palaearctic, the Afrotropical Region has only 2 species, one of them marginally reaching the Western Palaearctic; 8 species-group taxa occur in Nearctic Region, though most of them are confined to its southernmost areas, Florida and along the Mexican border (e.g., TODD 1955, 1961; POLHEMUS & POLHEMUS 1988b; POLHEMUS 1995b; ANDERSEN & WEIR 2004; CHEN et al. 2005; KMENT & JINDRA 2008; FAÚNDEZ & ASHWORTH 2015; NIESER et al. 2020).
The phylogenetic relationships within Ochteridae were addressed by MAHNER (1993) and summarized as follows: ( Megochterus + ( Ocyochterus + (American Ochterus + Old World Ochterus ))). The monophyly of the Old World Ochterus is well supported by the apomorphic structure of the right paramere, provided with a rounded apical cap and two subapical appendages; however, the monophyly of American Ochterus with a simple right paramere remains questionable, as their paraphyly towards the Old World clade ( Ochterus s. str.) is also possible ( MAHNER 1993). The first cladistic analysis of relationships among genera of Ochteridae was performed by YAO et al. (2011), who included the three recent genera and three fossil genera from Late Jurassic and Early Cretaceous of China, suggesting the following relationships: ((† Floricaudus + † Pristinochterus ) + († Angulochterus + ( Megochterus + ( Ochterus + Ocyochterus )))). For review of the fossil taxa assigned to Ochteridae and Ochteroidea stem group see Table 3. On the other hand, relationships among species-group taxa within the diverse Ochterus have never been rigorously tested. Considering the morphological homogeneity of this group, the use of molecular phylogenetic methods will be necessary.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Heterochterus timmsii Evans, 1971
| Kment, Petr, Carapezza, Attilio & Jindra, Zdeněk 2020 |
Heterochterus timmsii
| JELL P. A. 2004: 65 |
Heterochterus timmsii
| EVANS J. W. 1971: 150 |