
Hydroporus galloprovincialis, Manuel, Michael, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3652.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:5BA188F9-51B0-4990-BBA4-62D7BE573FC7 |
|
DOI |
https://doi.org/10.5281/zenodo.6160339 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87B0-8D07-FFDB-48E0-DD1749CDF943 |
|
treatment provided by |
Plazi |
|
scientific name |
Hydroporus galloprovincialis |
| status |
sp. nov. |
Hydroporus galloprovincialis View in CoL sp. n.
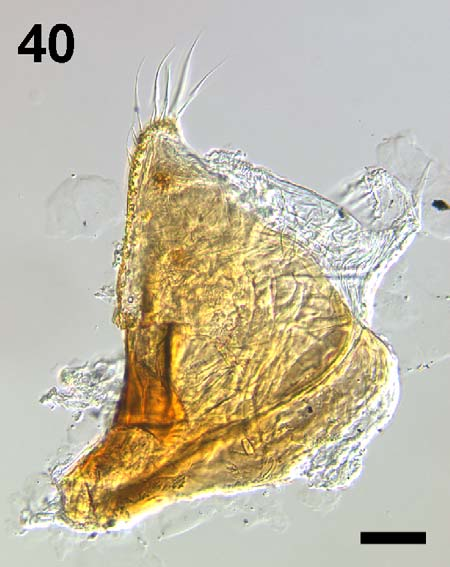
Figures 1–4 View FIGURES 1 – 2 View FIGURES 3 – 4 , 11 View FIGURES 11 – 13 , 14 View FIGURES 14 – 21 , 22–25 View FIGURES 22 – 39 , 40 View FIGURE 40 , 41–43, 54, 56.
Hydroporus normandi Régimbart, 1903 ; Guignot (1931–1933: 358; 1947: 102); Fery (1999: 237).
Type locality. Southern France, Provence, Pays d’Aix, Jouques, ca. 43°37’N 5°39’E, altitude ca. 280 m.
Type material studied. Holotype (male): “ 24.05.2011, France, ca. 2 km SE Jouques (13) [= département des Bouches-du-Rhône], spring with decaying leaves in a wet wood, Manuel leg.”, “ Holotype, Hydroporus galloprovincialis sp.n., Manuel det. 2013” [red] (MNHN). Paratypes: 36 exs, same data as the holotype (MNHN, ZSM, CMM, CHF, CLH). 32 exs, “ 23.05.2009, France, ca. 1 km SEE Jouques (13), little spring on the edge of a grassy marshland, Manuel leg.” (MNHN, CMM, CHF). 4 exs, “ 5.10.2009, France, ca. 1 km SEE Jouques (13), little spring on the edge of a grassy marshland, Manuel leg.” (two DNA-extracted vouchers, IBE-RA53 and IBE- RA54, deposited in the MNCN, and two additional ethanol-preserved specimens kept at the IBE). 2 exs, 9.10.2010, France, ca. 1 km SEE Jouques (13), little spring on the edge of a grassy marshland, Manuel leg.” (one DNAextracted voucher, IBE-RA289, and one additional ethanol-preserved specimen, both conserved at the IBE). 11 exs, “ 23.05.2011, France, ca. 1 km SEE Jouques (13), little spring on the edge of a grassy marshland, Manuel leg.” (BMNH, CMM, CLH). 1 ex. (male), “ 31.07.2011, France, ca. 2 km SE Jouques (13), headwaters of small stream in a wet wood, Manuel leg.” (CMM). The paratypes with the respective red label.
Other material examined. 1 female, “St. Maries [Saintes Maries de la Mer], détritus de la plage, 9.11.1926 ”, “Camargue, L. Puel”, female sex symbol, “Museum Paris, coll. R. DU BUYSSON 1900”, “ Hydroporus normandi normandi Régimbart Fery det. 1999”, “ Hydroporus galloprovincialis Manuel, 2013 . Manuel det. 2013”, coll. Guignot (MHNH). This specimen is thereafter referred to as the “Puel specimen”.
Habitus (holotype: Figs. 1–2 View FIGURES 1 – 2 ; other specimens: Figs. 3–4 View FIGURES 3 – 4 ; habitus of other H. normandi -complex taxa for comparison: Figs. 5–10 View FIGURES 5 – 10 ). Very elongate and parallel, flat dorsally, with large head and wide pronotum. Lateral outline continuous. Maximum width of body towards middle of total length, at about first third of elytral length. Maximum width of pronotum approaching maximum width of body (ratio between maximum pronotum width and maximum body width about 0.95); at posterior angles pronotum distinctly wider than elytra at shoulders. Head ferrugineous; pronotum and elytron dark brown, with their borders ferrugineous. Entire dorsal surface microreticulated but strongly shining, with short pubescence.
Head ferrugineous, with anterior part of clypeus and a broad transverse marking on vertex paler; between frons and vertex with two indistinct blackish spots (see Fig. 1 View FIGURES 1 – 2 ); lateral regions of vertex (behind eyes) dark brown to black. Antennae long, entirely testaceous; antennomere 2 almost as long and wide as antennomere 1; antennomere 4 distinctly shorter than antennomeres 3 and 5; length of antennomeres 5–9 slightly exceeding twice their width. Anterior outline of clypeus broadly sub-truncated. Eyes flat, their anterior outline continuous with that of head; interocular distance equalling half of pronotal width at posterior angles. Clypeus with two obliquely oriented and deeply impressed wide depressions, their central region with dense punctation; diameter of these punctures similar to that of frons punctures. Entire head surface (except along posterior margin) microreticulated with polygonal meshes, these more transverse near anterior margin of clypeus and on vertex behind eyes (not elsewhere on vertex). Punctures behind anterior margin of clypeus small and moderately dense; between clypeal depressions and on frons larger but irregular in size, their diameter equalling 1 to 2 meshes of microreticulation, with a few larger punctures reaching 3 meshes in diameter; punctures on frons rather regularly spaced with mean distance between two punctures about 2–3 meshes; vertex with small and sparse punctures; posterior margin of head with smooth surface between dense and small punctures; beside inner eye margin with impressed puncture lines and meshes of microreticulation smaller. Setae on head small and restricted to clypeal depressions and impressed puncture lines along inner eye margin.
Pronotum dark brown with anterior margin, posterior margin and sides ferrugineous. Lateral bead of pronotum rufo-testaceous. Pronotum width at posterior angles distinctly exceeding body width at shoulders. Posterior angles slightly obtuse. Sides of pronotum in posterior half almost parallel; in anterior half, pronotum sides regularly curved and weakly convergent. Pronotal bead thick, distinct from anterior to posterior angle, in lateral view equalling diameter of antennomeres 5–9 at their base. Surface uniformly microreticulated; meshes contiguous, polygonal and of same size as on head, anteriorly a bit smaller; before anterior line of large punctures, meshes with a transverse orientation. Near anterior margin, punctures very small and sparse (like on vertex); behind with a line of very coarse large punctures (their diameter about 2 to 3 meshes), distance between these punctures irregular, from less than one diameter to more than two diameters of a puncture. Laterally before posterior margin with a dense group of coarse and large punctures in a hardly perceptible linear-oblique depression; median half of pronotum without distinct puncture line before posterior margin. On disk with rather fine, moderately dense and regular punctation; in central disk region puncture diameter equalling 1 to 2 meshes of microreticulation and mean distance between punctures about 3–4 meshes; laterally, punctures slightly larger and coarser, but punctation not decisively denser than medially; in posterior third with punctures on average much larger, some reaching 3–4 meshes in diameter. Punctures apparently all bearing a distinct transparent seta; pronotum sides with longer setae.
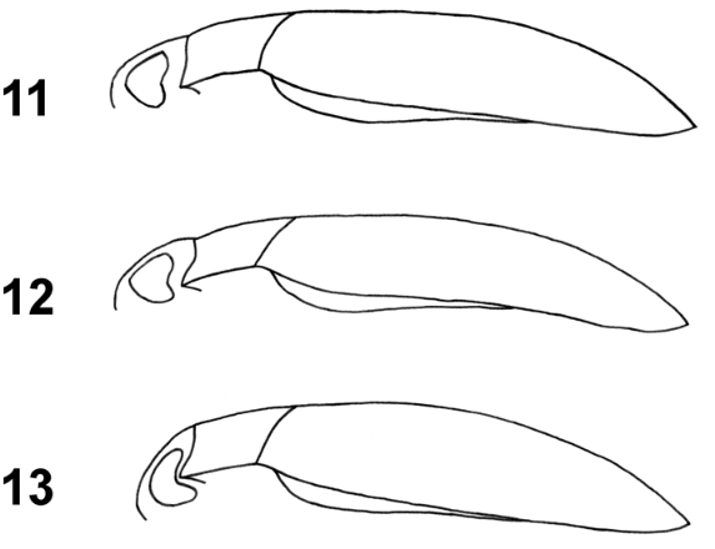
Elytra dark brown with shoulders, sides, apex, and a juxtasutural band ferrugineous; these paler areas not well delimited ( Fig. 1 View FIGURES 1 – 2 ). In dorsal view sides of elytra subparallel in their first two thirds; behind second third of elytral length sides strongly curved, subapically straight and then abruptly curved to apex, which is therefore rather pointed. In lateral view elytral margin distinctly ascending towards humeral angles; epipleuron not visible until humeral angle. Elytral bead much thinner than pronotal bead. Dorsal surface of elytra in anterior half very flat; in lateral view dorsal line of elytra continuous with that of pronotum and almost perfectly horizontal until slightly after elytral mid-length, then regularly curved towards apex ( Fig. 11 View FIGURES 11 – 13 ; comparison with other taxa Figs. 12–13 View FIGURES 11 – 13 ); in frontal view dorsal line of elytra in their anterior half horizontal between inner rows of coarse punctures; with a slight depression anteriorly next to suture (discernible using a horizontal light source). Microreticulation on elytra rather weakly impressed (at high magnification meshes appearing contiguous, Fig. 14 View FIGURES 14 – 21 ; comparison with other normandi -complex taxa Figs. 15–21 View FIGURES 14 – 21 ), more or less uniform, similar to that on pronotum but with meshes slightly larger. Punctation on elytra rather coarse, dense and regular, with punctures distinctly larger than on pronotum disk; punctures heterogeneous in size, their diameter varying from about one mesh to three meshes of microreticulation; diameter of largest punctures about 25 µm ( Fig. 14 View FIGURES 14 – 21 ); on elytral disk mean distance between punctures about two times diameter of a puncture. Inner and outer puncture lines distinct, their large punctures sparse and irregularly spaced; outer puncture line indistinct posteriorly. Elytron surface pubescent, each puncture apparently bearing a short seta; elytra bordered with very long setae (visible only in aqueous medium), these particularly conspicuous in posterior region (see Fig. 1 View FIGURES 1 – 2 ).
Ventral surface. Head reddish, genae blackish, gula rufo-testaceous. Palps entirely testaceous; last article of maxillary palps very long. Ventral surface of prothorax and elytral epipleura ferrugineous; posterior narrowed extremity of elytral epipleura darker. Rest of ventral surface black, except dark ferrugineous metaventral wings, antero-lateral part of metacoxal plates, posterior part of metacoxal processes, first and last abdominal ventrites, posterior and lateral parts of other abdominal ventrites. Genae microreticulated without punctures, gula smooth with small coarse punctures. Prosternal process ferrugineous, with blade lanceolate; lanceolate blade in cross section rather narrow, evenly convex (not roof-like and not carinate), medially smooth and laterally partly smooth and partly micro-granulose; margin distinctly beaded; apex bluntly pointed. Lanceolate blade with long golden setae on its margins. Between procoxae with an indistinct protuberance, and anteriorly with about 10 wellimpressed transverse ridges; surface between ridges smooth; ridge length decreasing anteriorly; first ridge placed slightly before procoxae; process not prolonged anteriorly as a narrow convexity onto prosternum. Antero-median region of prosternum ferrugineous (not distinctly darker than rest of prosternum), with many long golden setae; its surface rugose and finely granulose. Internal beading of elytral epipleura very thick; this beading not much thinner along metacoxal plate than along metepisternum. Posterior margin of metacoxal processes medially prolonged backwards, very slightly concavely sinuate along sides; midline of metacoxal processes deeply impressed over its entire length. Lateral lines of metacoxal processes distinct, elongate and weakly divergent, ending shortly behind posterior margin of metaventrite; inside metacoxal lines with strongly impressed grooves. Ventral surface smooth on metaventrite, internal half of metacoxal plates, between metacoxal lines, and in median region of abdominal ventrites 1 and 2. Elytral epipleura, metepisterna, lateral parts of metacoxal plates and of abdominal ventrites 1 and 2, and whole surface of abdominal ventrites 3–6 microreticulated; on elytral epipleura this reticulation weakly impressed, almost obsolete. On ventrite 6 microreticulation not distinctly more impressed than on ventrite 5. Sides of metaventrite with very large and coarse punctures; along internal margin of elytral epipleura, on whole surface of metacoxal plates and on sides of abdominal ventrites 1–2 with coarse punctures of medium size; on abdominal ventrites 3–6 punctation fine and sparse. Interlinear space of metacoxal processes with a few small punctures. Distinct and rather long setae present on metaventrite, on metacoxal plates, between lateral lines of metacoxal processes, on abdominal ventrites and elytral epipleura.
Legs uniformly rufo-testaceous; rather long and slender, particularly hind legs. Pro- and mesotarsomeres 1–3 ventrally densely covered with thick and short trumpet-like (distally enlarged) setae; central ventral surface of first protarsomere bearing 8 large sucker cups; on central surface of first mesotarsomere, sucker cups smaller and slightly more numerous (about 10). Protarsal claws small and simple, without apparent secondary sexual modification.
Note: The previous paragraphs described the morphology of the holotype. The other specimens examined are considered in the rest of the description (below).
Measurements. Holotype: total length (TL) 3.35 mm, total length without head 2.9 mm, maximum width (MW) 1.5 mm, ratio TL/MW 2.27. Paratypes: TL 3.15–3.7 mm (3.42 ± 0.12), TL without head 2.7–3.1 mm (2.92 ± 0.10), MW 1.4–1.7 mm (1.51 ± 0.05), TL/MW 2.18–2.36 (2.27 ± 0.04).
Male. Median lobe of aedeagus in lateral and ventral views as in Figs. 22–25 View FIGURES 22 – 39 (median lobes of other H. normandi -complex taxa shown for comparison in Figs. 26–39 View FIGURES 22 – 39 ); paramere as in Fig. 40 View FIGURE 40 .
Female. Gonocoxa (Figs. 41–43) with posterior half pigmented and rather thick; anterior half much paler, thin at its base then gradually thickened towards extremity which is club-shaped and often slightly bent (gonocoxa of H. n. normandi shown for comparison in Figs. 44–46). No conspicuous external differences to males.
Variation. Habitus moderately variable ( Figs. 1–4 View FIGURES 1 – 2 View FIGURES 3 – 4 ); in some specimens maximum body width slightly after mid-length; rarely pronotum at posterior angles not wider than body at shoulders. In some specimens (like holotype), sides of pronotum in posterior half parallel and maximum pronotum width at posterior angles ( Figs. 1, 2 View FIGURES 1 – 2 , 4 View FIGURES 3 – 4 ), but in other specimens sides distinctly converging to base before posterior angles (in this case, maximum pronotum width a little before posterior angles: Fig. 3 View FIGURES 3 – 4 ). Dorsal pigmentation strongly variable; brown discal area on pronotum and elytron present in darker specimens (like holotype), but in paler specimens indistinct on pronotum and sometimes also on elytra, in latter case dorsal pigmentation entirely ferrugineous ( Fig. 4 View FIGURES 3 – 4 ); uniformly pale specimens still found among beetles kept alive for six weeks after capture. In darker specimens intensity of central brown coloration and extension of surrounding paler areas highly variable on pronotum and elytra ( Figs. 2 View FIGURES 1 – 2 , 3 View FIGURES 3 – 4 ). Exceptionally, antennomeres 5–9 indistinctly darkened in their distal half. Average size of punctures on elytra somewhat variable; very small punctures (diameter equivalent to less than one mesh of microreticulation) rare or absent in most specimens but sometimes more numerous. Extension of microreticulation on metacoxal plate variable but in most specimens confined to its lateral half. Notable variation observed for shape of median lobe of aedeagus in profile and ventral views (compare Figs. 22–23 and 24–25 View FIGURES 22 – 39 ). In specimens from type locality, pubescence always distinct on whole surface of pronotum and elytra, but its development much variable (compare Figs. 2 View FIGURES 1 – 2 and 3 View FIGURES 3 – 4 ); length of setae on elytral disk up to 90 µm in most pubescent paratypes; in contrast pubescence much sparser and even indistinct on pronotum and elytron disks in the Puel specimen ( Fig. 4 View FIGURES 3 – 4 ); this specimen otherwise not showing any deviating character with respect to those of locus typicus.
Habitat. In Jouques, the species was found at two spots located at a distance of about 2 km from each other. In both situations, the beetles were strictly restricted to a small spring where clean cold water slowly flows out from the calcareous substratum; they were conspicuously absent from surrounding waters. Both spots were also characterised by an abundance of fine organic particles and decaying vegetal material (partially decomposed leaves), completely covering the calcareous substratum. These two habitats otherwise differ significantly. The first population lives on the margin of a grassy wetland ( Fig. 47 View FIGURES 47 – 50 ), in an open situation among large plants ( Carex acuta , Carex acutiformis , Cyperus longus , Typha sp., Eupatorium cannabinum , Epilobium hirsutum , Equisetum telmateia ). These plants provide shade to the spring, which is very small (diameter about 30 cm) and shallow (water depth less than 10 cm) ( Fig. 48 View FIGURES 47 – 50 ) and gives birth, when active in spring, to a minute brooklet slowly flowing among the grasses towards the stream of St Bachi, a few dozen meters downstream. Hydroporus galloprovincialis sp. n. was very abundant in this little spring on May 23, 2009 and was found in lower abundance on May 23, 2011; during visits in summer time the habitat was completely dry and the beetle could not be found; visits during October 2009 and 2010 when water level was extremely low yielded only very few specimens. Other dytiscid species observed at this place: (i) in the spring itself: Agabus paludosus (Fabricius, 1801) (abundant), Agabus bipustulatus (Linnaeus, 1767) , Hydroporus discretus discretus Fairmaire & Brisout, 1858 and Hydroporus planus (Fabricius, 1781) (not abundant); (ii) elsewhere in the nearby marsh (in addition to the already mentioned species): Hydroporus tessellatus (Drapiez, 1819) , Hygrotus impressopunctatus (Schaller, 1783) , Graptodytes bilineatus (Sturm, 1835) , Agabus didymus (Olivier, 1795) , Agabus brunneus (Fabricius, 1798) , Ilybius chalconatus (Panzer, 1796) , Dytiscus marginalis Linnaeus, 1758 . The second spot is a larger spring from which originates a rather important stream (about 2 meters wide 50 m downstream from the spring), affluent of the St Bachi stream. The spring is located in a small wet forest and is full of decaying tree leaves and branches ( Figs. 49 and 50 View FIGURES 47 – 50 ), with no aquatic vegetation and no surrounding hygrophilic plants. Hydroporus galloprovincialis sp. n. was found there in great abundance on 24 May 2011; at this time the diameter of the spring was about 1 meter and its depth about 10 cm; water was clear but the organic substrate was heavily covered with orange deposit due to the activity of ironoxydising bacteria ( Fig. 50 View FIGURES 47 – 50 ). During a subsequent visit on July 2011, the spring was completely dry and the stream was alimented by water flowing from the substrate about 20 meters downstream; at this level only a single specimen of H. galloprovincialis could be found despite intensive search. Other dytiscid species observed at this place were I. chalconatus , A. bipustulatus and A. paludosus .
At Saintes-Maries de la Mer, a single specimen (shown in Fig. 4 View FIGURES 3 – 4 ) was found in 1926 by L. Puel among “detritus of the beach” (previously this specimen had been identified as Hydroporus n. normandi , see Guignot 1931–1933 and Fery 1999). This certainly cannot represent the original habitat of the beetle, which was probably flowed out from an inland spring. At Saintes-Maries de la Mer there is a famous spring located in the crypt of the church. The latter was built on the burial place of the Holy women St Mary Jacobe, St Mary Salome and St Sarah, who according to the tradition arrived there after having been arrested in Jerusalem and set off on a boat then abandoned on a raft, together with Mary-Magdalene, Lazarus and other disciples of Jesus, towards the middle of the 1st Century. Perhaps H. galloprovincialis is an inhabitant of this spring and witnessed these events. Otherwise, the insect found by L. Puel might come from a more distant locality from which it would have been transported in the waters of the nearby Petit Rhône River, which ends in the sea a few hundred meters west from the Saintes- Maries beach.
Study area. The type locality is situated on the border of the Rhone valley, a few kilometres south from the Durance River (affluent of the Rhône), in the transition zone towards the Provençal Alps. The climate is of Mediterranean type but with relatively cold winters and rather high average annual precipitations (climatological data from Meteo France, station of Le Luc, ca. 60 km ESE Jouques at a similar altitude: average annual temperature 8.5°C–20.9°C, coldest month January 1.7°C–12.4°C, warmest month July 16.4°C–31.1°C; average annual precipitations 857.4 mm, 68 days per year with precipitations, highest in April–May and September– January with a maximum in October). The main geological substrata of the area are various kinds of Mesozoic and Cenozoic calcareous rocks (geological data from the BRGM). The springs where H. galloprovincialis sp. n. was discovered belong to an important system of resurgences (hydrological data from Geoportail) concentrated around a large geological fault line on the southern versant of the mountain of Vautubière, which culminates at an altitude of 658 m ca. 3.5 km ENE from the collecting sites. The southern extremity of this fault line is the place that the Romans chose to be the starting point of one of the main aqueducts of the region at that time, which alimented the city of Aix-en-Provence (the aqueduct of Traconnade, now almost entirely destroyed). The mountain of Vautubière itself is an anticlinal structure with Jurassic (Kimmeridgian and Tithonian) calcareous rocks in its centre, and on the south versant late Miocene molasse (west from the fault line) and early Cretaceous limestone and marl (east from the fault line) in abnormal contact. Among the two collecting sites described above (under Habitat), the first one is located ca. 200 m west from the fault line at the bottom of the southern versant of the mountain, in contact with the Holocene peat which forms the substratum of the marsh (a helocrene). The second spot is a small limnocrene located east from the fault line on Würm age colluvium.
Derivatio nominis. The specific epithet “ galloprovincialis ” refers to the known geographic distribution of the species. It is an adjective in the nominative singular.
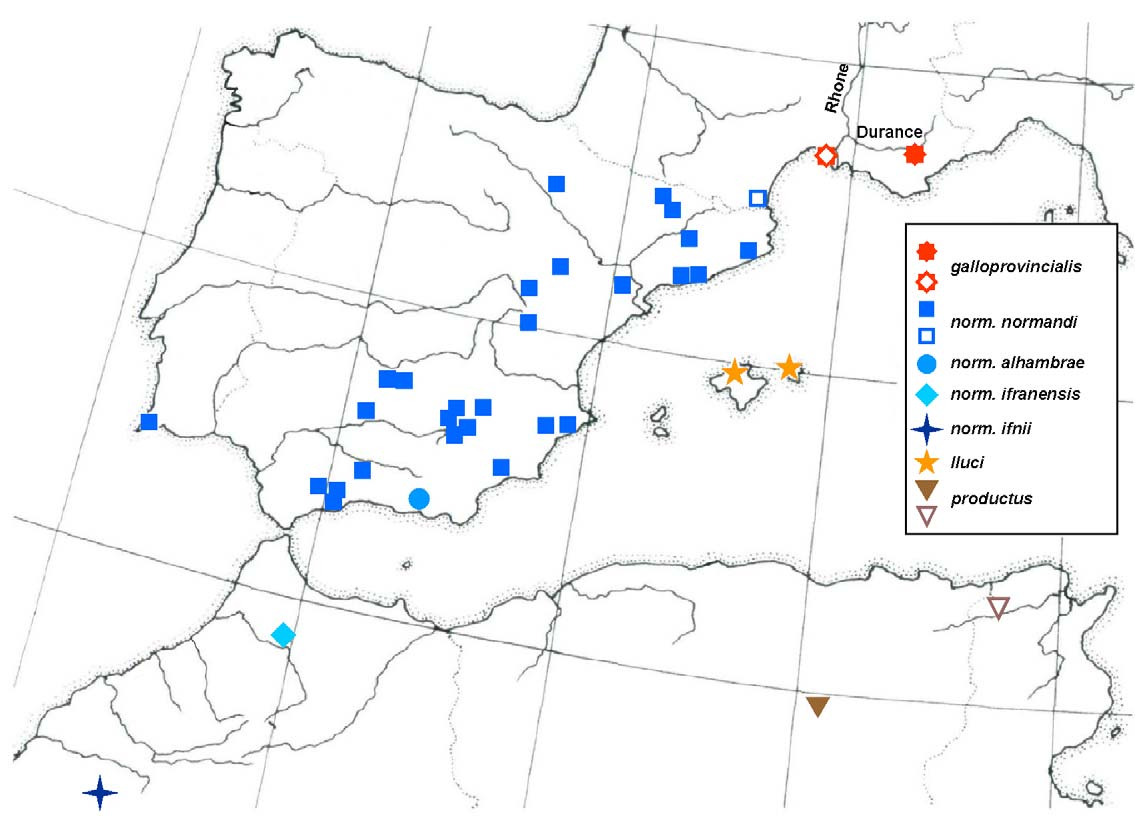
Distribution. So far the species is known only from two localities, both in the department “Bouches-du- Rhône” of the French region Provence ( Fig. 52 View FIGURE 52 ): Jouques (the type locality) and Saintes-Maries de la Mer in Camargue (a single observation in 1926). Hydroporus galloprovincialis sp. n. is therefore endemic to south-eastern France.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |