
Oecetis ancorospina, Moura & Quinteiro, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5361.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:17E949E3-507F-4500-A0AA-AC06D2BA6451 |
|
DOI |
https://doi.org/10.5281/zenodo.10166858 |
|
persistent identifier |
https://treatment.plazi.org/id/A8F733BB-EA1E-4E71-997A-F64CA6F4DE70 |
|
taxon LSID |
lsid:zoobank.org:act:A8F733BB-EA1E-4E71-997A-F64CA6F4DE70 |
|
treatment provided by |
Plazi |
|
scientific name |
Oecetis ancorospina |
| status |
sp. nov. |
Oecetis ancorospina , n. sp. ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 )
urn:lsid:zoobank.org:act:A8F733BB-EA1E-4E71-997A-F64CA6F4DE70
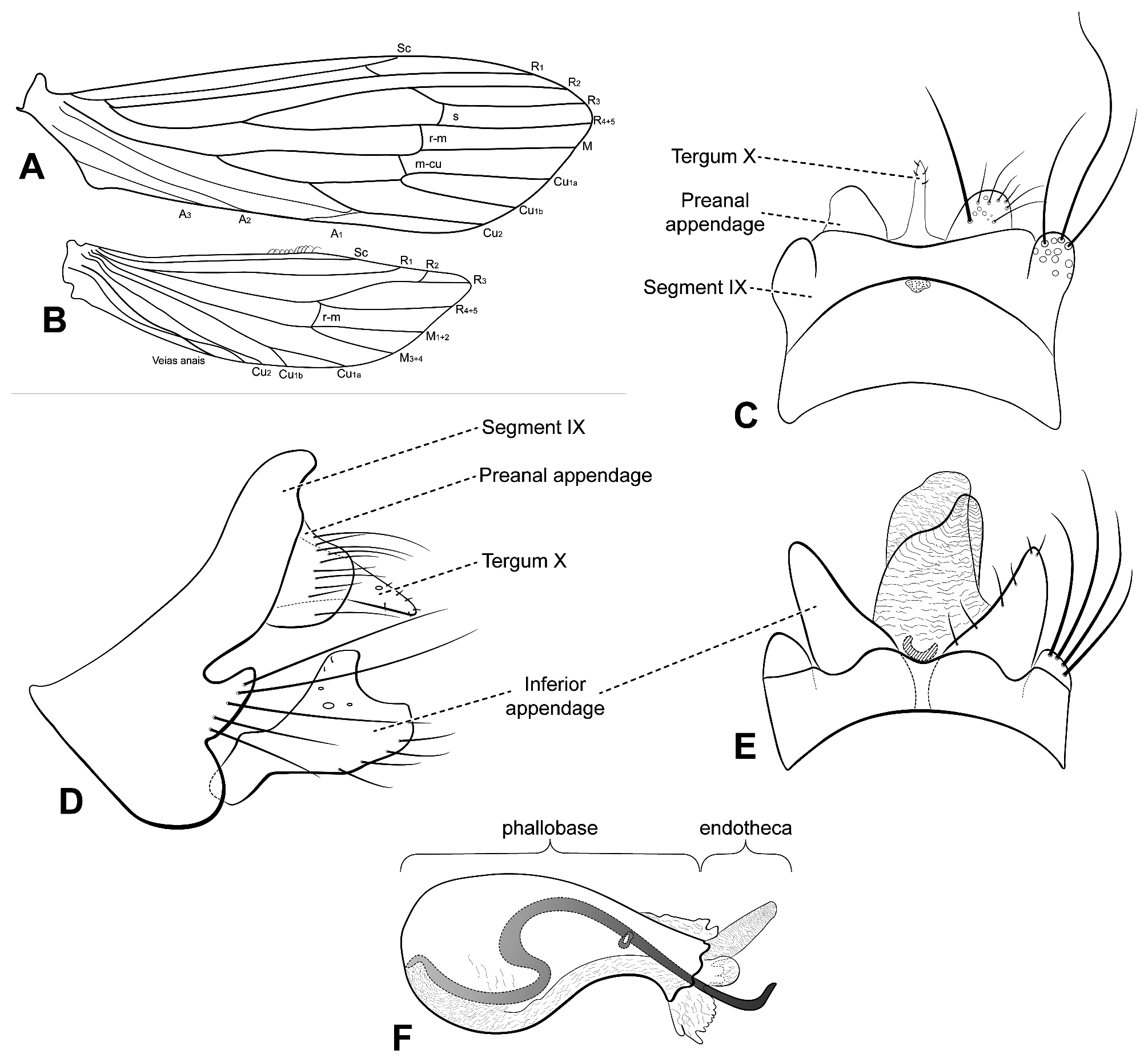
Diagnosis. This new species belongs to the Oecetis testacea Species Group due to the presence of abdominal terga V–VIII covered by honeycomb microstructure ( Figs 2A, 2B View FIGURE 2 ; Chen 1993; Quinteiro & Holzenthal 2017). It can be distinguished from the other species in the group by the elongate phallic spine with a curved tip ( Fig. 2C View FIGURE 2 ), the inferior appendages each with a broad distal lobe and rounded apex, and a discrete dorsal lobe with a truncate apex ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). The presence of a keel-shaped lateral process of segment IX with an acute apex can be helpful as an additional diagnostic character ( Fig. 2A View FIGURE 2 ).
Description. Male forewing average length 5.2 mm (5.0– 5.9 mm, n = 7), holotype 5.1 mm. Hind wing average length 4.1 mm (3.8–4.5 mm, n = 7), holotype 4.0 mm.
Head. Pale yellow (in alcohol). Antennae three times length of forewing; scape stout; pedicel short; maxillary palps pale yellow, 5-segmented; labial palps pale yellow, 3-segmented.
Thorax. Pterothorax yellowish brown. Forewings pale yellow; dark bands on wings absent; dark spots on forks, junctions, and vein tips absent; colored setae absent; forks I and V present (both rooted); sectoral crossvein (s) distal of with r-m; m-cu crossvein basal of with r-m ( Fig. 3A View FIGURE 3 ). Hind wings pale yellow, forks I (stalked) and V (sessil) present ( Fig. 3B View FIGURE 3 ). Legs pale yellow. Tibial spur formula 1,2,2; each foreleg tibial spur reduced.
Abdomen. Reticular patches (honeycomb texture) on terga V, VI, VII, and VIII ( Fig. 2 View FIGURE 2 ). Acrotergite present between segments VIII and IX ( Fig. 3C View FIGURE 3 ). Segment IX short, annular in lateral view ( Fig. 3D View FIGURE 3 ); posterolateral process present on each side, keel-shaped laterally, with long setae on posterior margin ( Fig. 3D View FIGURE 3 ). Preanal appendages short, apically round, broad basally, with apical setae ( Figs 3C, 3D View FIGURE 3 ). Tergum X undivided in dorsal and lateral views, longer than preanal appendages, tapering apically, with apex acute ( Figs 3C, 3D View FIGURE 3 ). Inferior appendages each 1-segmented, setose; dorsal lobe subquadrate, apex truncate, projecting dorsad ( Fig. 3D View FIGURE 3 ); ventral lobe absent ( Fig. 3D View FIGURE 3 ); distal lobe round ventrally, apex truncate; smooth concavity formed from apex of dorsal lobe to apex of distal lobe ( Fig. 3D View FIGURE 3 ). Phallic apparatus slightly asymmetrical, elongate, ventrally sigmoid; phallobase large basally, tapering posteriorly, slightly constricted subapically and flared apically ( Fig. 3F View FIGURE 3 ); one phallic spine present, long, helical, with apex hooked dorsad; phallotremal sclerite present, horseshoe-shaped ( Fig. 3E View FIGURE 3 ); endotheca multilobed ( Figs 3E, 3F View FIGURE 3 ).
Female and immatures. Unknown.
Holotype male: Brazil, Pará, Bragança, Jiquiri, Ramal Arauá . Sitio Cururutuia , 01°04’46.9”S, 46°44’17.6”W, 13.xii.2019, alt 19m., light trap, Quinteiro, F.B., Moura, L., Saldanha, G., Nunes, A. (alcohol, MPEG). GoogleMaps
Paratypes: Same data as holotype, 1 male (alcohol, UFBA) GoogleMaps . Same data as holotype except 26.ix.2020, Quinteiro, F.B, Costa, A.M., 2 males (alcohol, UFBA) GoogleMaps . Same data as holotype except 11.vii.2020, Quinteiro, F.B, Costa, A.M., 1 male (alcohol, UFBA) GoogleMaps . Same data as the holotype except 21.x.2022, bandeja, col. Quinteiro, F.B., Costa, A.M., 2 males (alcohol, UFBA) GoogleMaps . Same data as holotype except 11.vii.2021, bandeja, col. Quinteiro, F.B, Moura. L., Saldanha, G., 1 male (alcohol, MPEG) GoogleMaps . Same data as holotype except 13.viii.2021, bandeja, col. Quinteiro, F.B, Costa, A.M., 1 male (alcohol, MPEG) GoogleMaps . Same data as holotype, except Santa Luzia do Pará, Rio Aripé , Chácara Prof. Elizélia , 01°22’30.6”S, 47°03’18.5”W, 20.xi.2020, alt 50 m., lençol, col. Quinteiro F.B., Costa A.M., Moura, L, Saldanha, G., 2 males (alcohol, MPEG) GoogleMaps .
Distribution. Brazil (Pará state)
Etymology. Ancora (Latin) = curved, hooked; spina (Latin) = spine. The species’ name is a reference to the long, apically curved phallic spine.
Taxonomic discussion and remarks. Oecetis ancorospina n. sp. belongs to the Oecetis testacea Species Group due to the presence of reticulate microstructure on abdominal terga V–VIII, similar to honeycombs ( Quinteiro & Holzenthal 2017). There are only three valid species of the Oecetis testacea Species Group reported for the Neotropical Region: Oecetis iara Henriques-Oliveira, Dumas & Nessimian 2014 , Oecetis plenuspinosa Quinteiro &, Holzenthal 2017, and Oecetis meronai Gibon 2019 . The new species is characterized by the presence of a short, annular segment IX, with the ventral half enlarged and the anterior lateroventral margin strongly projected anterad with an acuminate apex ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). Oecetis iara has its segment IX somewhat falciform and O. plenuspinosa , despite having its segment IX slightly enlarged medially, the anterolateral margins are round. Oecetis meronai has its segment IX also annular and short but does not bear the acuminate anterior lateroventral margin with a distinct projection; instead, its anteroventral margin is smoothly convex ( Gibon 2019).
The posterolateral process on segment IX in O. ancorospina n. sp. is keel-shaped, pointing posterad, with an acute apex ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). In contrast, the posterolateral process of O. iara is digitate, with a round apex ( Henriques-Oliveira et al. 2014). The posterolateral process of segment IX in O. plenuspinosa is similar to that of O. ancorospina n. sp., in lateral view, since both of them have acute apices ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). However, the lateral process in the new species points posterad, while in O. plenuspinosa , the lateral process points dorsad. In dorsal view, the posterolateral processes of segment IX in O. ancorospina n. sp. are each apically round ( Figs 2B View FIGURE 2 , 3C View FIGURE 3 ), but in O. plenuspinosa it is acute. In O. meronai , the posterolateral processes of segment IX are in a different position than in O. plenuspinosa and O. ancorospina n. sp., in the dorsal half, beneath the preanal appendages ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ).
In lateral view, the preanal appendages of the new species are apically round and short, each with a broad base ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ), while those of O. iara , O. meronai , and O. plenuspinosa are digitate, with narrower apices than in O. ancorospina n. sp.
Tergum X of the new species is not divided into dorsal and ventral portions ( Figs 2A, 2B View FIGURE 2 , 3C, 3D View FIGURE 3 ), as in O. iara and O. plenuspinosa . However, the undivided tergum X of the new species is similar to the dorsal portion of tergum X in Oecetis iara , which is subtriangular and acute apically, in lateral view ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). In dorsal view, tergum X in all these species is broadly based, tapering distally, with an apex acute ( Henriques-Oliveira et al. 2014). In O. plenuspinosa , the dorsal portion of tergum X is elongate, at least three times longer than the preanal appendages, and cylindrical, with the apex distinctly clavate ( Quinteiro & Holzenthal 2017). In O. meronai , even though tergum X has a distinctly clavate apex, this structure is divided into dorsal and ventral portions, differently than in O. iara , O. plenuspinosa , and O. ancorospina n. sp.
The phallic apparatus of O. ancorospina n. sp. ( Fig. 3F View FIGURE 3 ), is distinct from that of O. plenuspinosa and O. iara since it has a much broader base than its apex. The phallic apparatus of O. iara is cylindrical; in O. plenuspinosa , it is broad apically with a small base. In O. meronai , the phallic apparatus is somewhat ovoid. All four species have phallic spines. In lateral view, Oecetis ancorospina n. sp. has a long helically curved phallic spine, with a tip hooked dorsad, U-shaped dorsally ( Figs 2B, 2C View FIGURE 2 , 3F View FIGURE 3 ). Oecetis iara has a short phallic spine, bent ventrad ( Henriques-Oliveira et al. 2014), and O. plenuspinosa has ten small falciform spines symmetrically distributed in two groups of five ( Quinteiro & Holzenthal 2017). The phallic spine of O. meronai resembles that of O. ancorospina n. sp, since both are long, helical, and curved ( Figs 2C View FIGURE 2 , 3F View FIGURE 3 ); however, the phallic spine in O. ancorospina n. sp. has a hooked tip, but in O. meronai the tip is straight.
The inferior appendages of the new species are also very distinct from those of other Neotropical species of the O. testacea Species Group ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ). Each has a projected dorsal lobe with a truncate apex, whereas in O. iara , the inferior appendages lack dorsal and ventral lobes; each of those in Oecetis plenuspinosa has only a round ventral lobe; and in O. meronai has a slightly quadrate ventral lobe. Concerning the distal lobe of the four species, O. ancorospina n. sp. has it short, and slightly enlarged apically, forming a C-shaped angle with the dorsal lobe. In O. meronai , the distal lobe is cylindrical with a round tip and an inner curve in ventral view. Each inferior appendage of O. plenuspinosa is characterized by having a long distal lobe with a large distal half and truncate apex and that of O. iara has a digitate, short distal lobe.

| UFBA |
UFBA |
| MPEG |
Museu Paraense Emilio Goeldi |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |