
Polybius dioscurus, García-Raso & Moukrim & Schubart & Cuesta, 2024
|
publication ID |
https://doi.org/ 10.5852/ejt.2024.930.2501 |
|
publication LSID |
lsid:zoobank.org:pub:3FA94665-E6B2-4455-B40D-4ACDB71AAA55 |
|
DOI |
https://doi.org/10.5281/zenodo.10979179 |
|
persistent identifier |
https://treatment.plazi.org/id/F730427D-FF98-FFF0-6244-FD67EF33FE63 |
|
treatment provided by |
Plazi |
|
scientific name |
Polybius dioscurus |
| status |
sp. nov. |
Polybius dioscurus sp. nov.
urn:lsid:zoobank.org:act:E3FB5F11-959A-43C0-85A4-9A9FB88ED7C8
Figs 3–5 View Fig View Fig View Fig , 6A, C View Fig , 7B View Fig , 8C–D View Fig , 9 View Fig
Portunus marmoreus ‒? Bell 1844–1853: 105, in part, not unnumbered fig. —? White 1857: 50, in part. —? Palmer 1927: 889, in part (second abnormal specimen). — Nobre 1931: 66, figs 31–32; 1936: 36, pl. 11 fig. 19.
Portunus barbarus ‒ Monod 1956: 173, 179, in part.
Macropipus barbarus View in CoL ‒ Forest & Guinot 1956: 37, in part (Atlantic specimens only), not fig. 5.
Macropipus holsatus View in CoL ‒ Cardona Bendito 1965: 152, pl. 2 fig. d.
Macropipus marmoreus View in CoL –? Bourdon 1965: 29 (in part).
Macropipus vernalis View in CoL ‒ Neves 1975: 32, in part, figs 10, 11c.
Liocarcinus vernalis View in CoL ‒ García Raso 1984: 107 in part. — González Gurriarán & Méndez 1986: 43 (key), 96, fig. 30, 210, foto 19. — d’Udekem d’Acoz 1989: 184, in part, fig. 16, not fig. 17; 1991: 84, fig. 1; 2011: 78, fig. 1. — Adema 1991: 111, figs 1–2. — d’Udekem d’Acoz & Rappé 1991: 95, figs 3, 6, 9, 12. — Fransen 1991: 124. — Ingle 1996: 57, 138, fig. 37. — Ingle & Clark 1998: 224, figs 1c, 2c, 2f. — Livory 1998: 49. — López de la Rosa et al. 2002: 87. — McCarthy et al. 2005: 20. — Ashelby 2006: 1341. — Martin 2011: 221, unnumbered colour photographs. — De Blauwe & Decleer 2017: 173, unnumbered colour photographs.
Liocarcinus sp. aff. holsatus ‒ d’Udekem d’Acoz 1986: figs 17, 20.
Polybius (Polybius) vernalis View in CoL ‒ d’Udekem d’Acoz 1999: 222, in part, unnumbered fig. p.1.
Liocarcinus cf. holsatus View in CoL – Moukrim et al. 2010: fig. 2f.
Diagnosis
Carapace glabrous, wider than long, smooth. Frontal region with three slightly protruding blunt triangular teeth, with a tuberculate rim, continued by the inner-orbital angle, middle tooth rounded, subacute and does not reach same proportions as the lateral ones, which are more developed (wider and rounded), the curvature between rostral teeth is rounded. Anterolateral margins of the carapace curved with five teeth (including the external orbital), the fifth and the fourth being somewhat larger, the third and fourth with a rectilinear outer face. Chelipeds (P1) subequal; propodus outer face of the palm with 3 longitudinal keels along entire length; dorsal edge with a smooth rounded keel ending in a blunt distal tooth; internal face without keels; dactyl externally with a central depression, between two rounded ridges; dorsally, three rounded longitudinal ridges with depressions between them; merus smooth; carpus with one strong and protruding tooth on antero inner dorsal margin, on the anterior outer upper part one well-marked and rounded projection and two others below. Walking legs with merus, carpus and propodus smooth; dactylus quite rectilinear dorsally (P2 wider, robust and with a slightly more curved tip), with a more or less quadrangular section, and with angles defined by rounded longitudinal edges, the dactylus of P4 has a dense row of short setae in the basal third (or a little more), in P5 the dactyl is strongly flattened, paddle-like, oval, ending in a point. First sexual male pleopod (gonopod) with curved distal part, which forms an angle of almost 90° (more curved in large specimens) and ends in a small point.
Etymology
The name of the species is derived from the Greek mythology. Castor and Pollux are twin half-brothers in Greek and Roman mythology, known together as the Dioscuri (plural). The singular form is Dioscurus. The name, which is a noun in apposition, refers to the great similarity between the new species and Polybius vernalis ( Risso, 1827) .
Material examined
Holotype SPAIN • ♂ (26.4 mm CW); outer Bay of Cádiz, Cádiz, Valdelagrana ; 36°43′ N, 06°14′ W; 3.3–6.6 m depth; 31 Oct. 1996; I. López de la Rosa leg.; fine sand bottoms; GenBank: OR555887 –16S, OR557369 – COI; CRUST_ICMAN/3759 . GoogleMaps
Paratypes BELGIUM – Southern North Sea • 3 ♂♂, 2 ♀♀; Groenendijk ; 51°08′30″ N, 2°41′ E; 1 m depth; 10 Nov. 1990; C. d’Udekem d’Acoz leg.; sandy beach, net refuse of shrimp fishermen; RBINS, INV. 187227 GoogleMaps • 1 ♂ (illustrated); between Nieuwpoort and Oostduinkerke ; 51°09′ N, 2°42′ E; 1 m depth; 13 Oct. 2018; d’Udekem d’Acoz leg.; sandy beach, net refuse of shrimp fishermen; RBINS, INV. 187223 GoogleMaps • 4 ♂♂, 6 ♀♀ (colour in life photographed); between Nieuwpoort and Oostduinkerke ; 51°09′ N, 2°42′ E; 1 m depth; 13 Oct. 2018; d’Udekem d’Acoz leg.; sandy beach, net refuse of shrimp fishermen; RBINS, INV. 187230 GoogleMaps • 20 ♂♂; between Oostduinkerke and Koksijde ; 51°08′ N, 2°39′ E; 1 m depth; 27 Sep. 2019; d’Udekem d’Acoz leg.; sandy beach, net refuse of shrimp fishermen; RBINS, INV. 187229 GoogleMaps • 1 ♂; same collection data as for preceding; RBINS, INV. 187226 GoogleMaps .
FRANCE – Brittany • 1 ♂; Saint Lunaire ; 48°38′ N, 2°06′ W; 18 Mar. 1988; d’Udekem d’Acoz leg.; RBINS, INV. 187222 GoogleMaps • 9 ♂♂, 7 ♀♀; W of Crozon Peninsula, Anse de Dinant ; 48°15′ N, 4°33′ W; 1 m depth; 30 Mar. 2010; d’Udekem d’Acoz leg.; net refuse of Donax fishermen; RBINS, INV. 187228 GoogleMaps .
MOROCCO – Agadir Bay • 1 ♂ (36.7 CW); from Pte. d’Anza to South of Oued Souss ; 30°26′ N, 09°40′ W – 30°18′ N, 09°37′ W; 6–25 m depth; May 1999; Gofas and Moukrim leg.; sandy bottoms; GenBank: OR555890 –16S, OR557370 –COI; CRUST_ICMAN/3762 GoogleMaps • 1 ♂ (40.6 mm CW); same collection data as for preceding; GenBank: OR555889 -16S; CRUST_ICMAN/3763 GoogleMaps • 6 ♂♂; same collection data as for preceding; CRUST_ICMAN/3777 GoogleMaps • 1 ♂, 1 ♀; same collection data as for preceding; MNHN-IU-2022-4081 GoogleMaps • 1 ♂, 1 ♀; same collection data as for preceding; NHMUK 2024.3 About NHMUK , 2024.4 About NHMUK GoogleMaps .
PORTUGAL – Algarve • 107 ♂♂ (one with Sacculina ); Praia de Cabanas ; 37°08′ N, 7°35′30″ W; 1 m depth; 20 Jul. 1988; d’Udekem d’Acoz leg.; net refuse of Donax fishermen; RBINS, INV. 187232 GoogleMaps • 67 ♀♀ (some ovigerous); same collection data as for preceding; RBINS, INV. 187233 GoogleMaps .
SPAIN • 1 ♀ (19.3 mm CW); same collection data as for holotype; GenBank: OR557368 –COI; CRUST_ICMAN/3760 GoogleMaps • 2 ♀♀, 1 ♂; inner part of the Cádiz Bay ; 1994 – 1995; fine muddy sand bottoms with Caulerpa prolifera ; CRUST_ICMAN/3778 • 8 ♀♀, 6 ♂♂ (30.0 mm max CW); Cádiz, Chipiona ; CRUST_ICMAN/3779 • 1 ♂; Gulf of Cádiz, Huelva, Isla Canela ; 14 Jan. 2021; Helena Marco-Herrera leg.; FEMP-04: stn 10C; CRUST_ICMAN/3772 • 1 ♀; same collection data as for preceding; CRUST_ ICMAN/3773 • 1 ♀; same collection data as for preceding; GenBank: OR555886 –16S; CRUST_ ICMAN/3764 • 3 ♀♀; Gulf of Cádiz, Huelva, Doñana National Park ; 12 Apr. 2021; FEMP-04: stn 6; CRUST_ICMAN/3774 to CRUST_ICMAN/3776 • 4 ♀♀, 1 ♂; same collection data as for preceding; CRUST_ICMAN/3780 • 6 ♀♀; same collection data as for preceding; 12 Jan. 2021; 0.5–1 m depth; FEMP-04: stns 10C, 6 and 2; fine sandy bottoms; CRUST_ICMAN/3781 • 1 ♀; W Mediterranean, Alboran Sea, Málaga, Marbella ; 2–4 m depth; 24 Mar. 2021; García Raso leg.; from Donax trunculus Linnaeus, 1758 fisheries; GenBank: OR555891 –16S, OR557371 –COI; CRUST_ICMAN/3765 • 1 ♂; Málaga, Fuengirola ; 2–4 m depth; Aug. 1983; García Raso leg.; from Donax trunculus fisheries; CRUST_ICMAN/3782 • 18 ♀♀ (smallest ovigerous ♀: 16.4 mm width), 14 ♂♂; Málaga ; 1979 – 1981, García Raso leg., from Donax trunculus fisheries; CRUST_ICMAN/3783 .
Description
MAXIMUM SIZE. CL/CW: 30.7/ 40.6 mm.
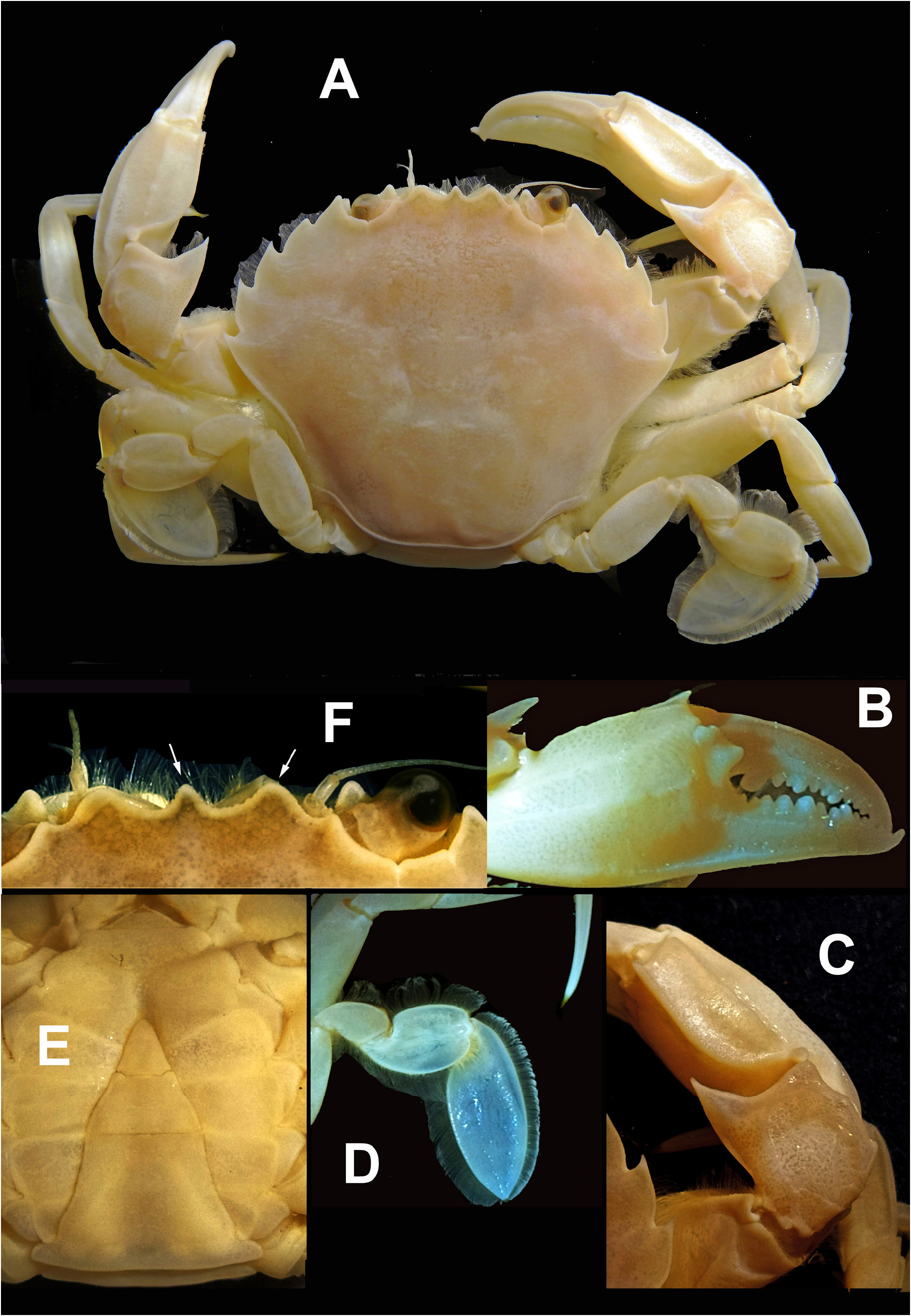
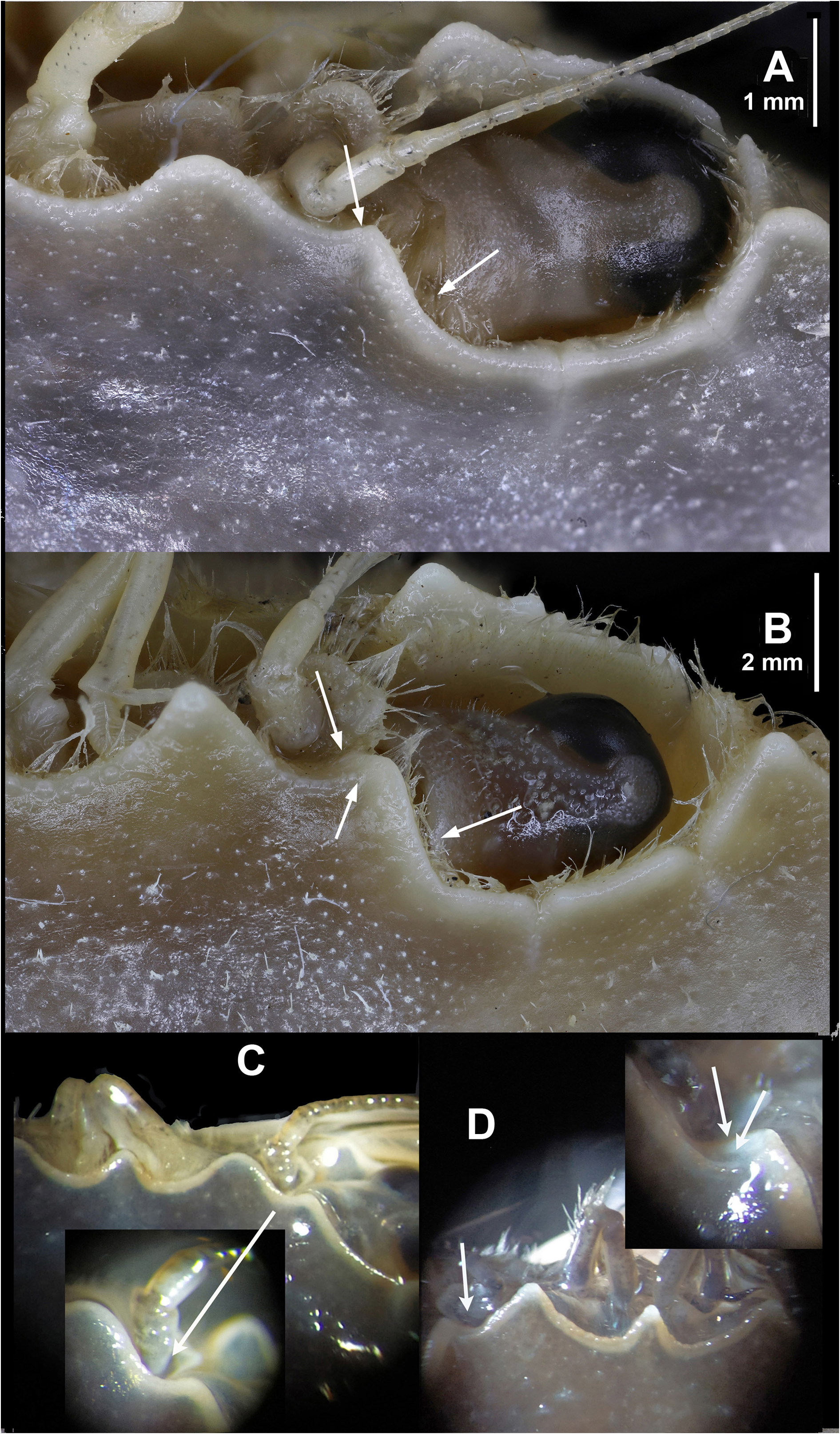
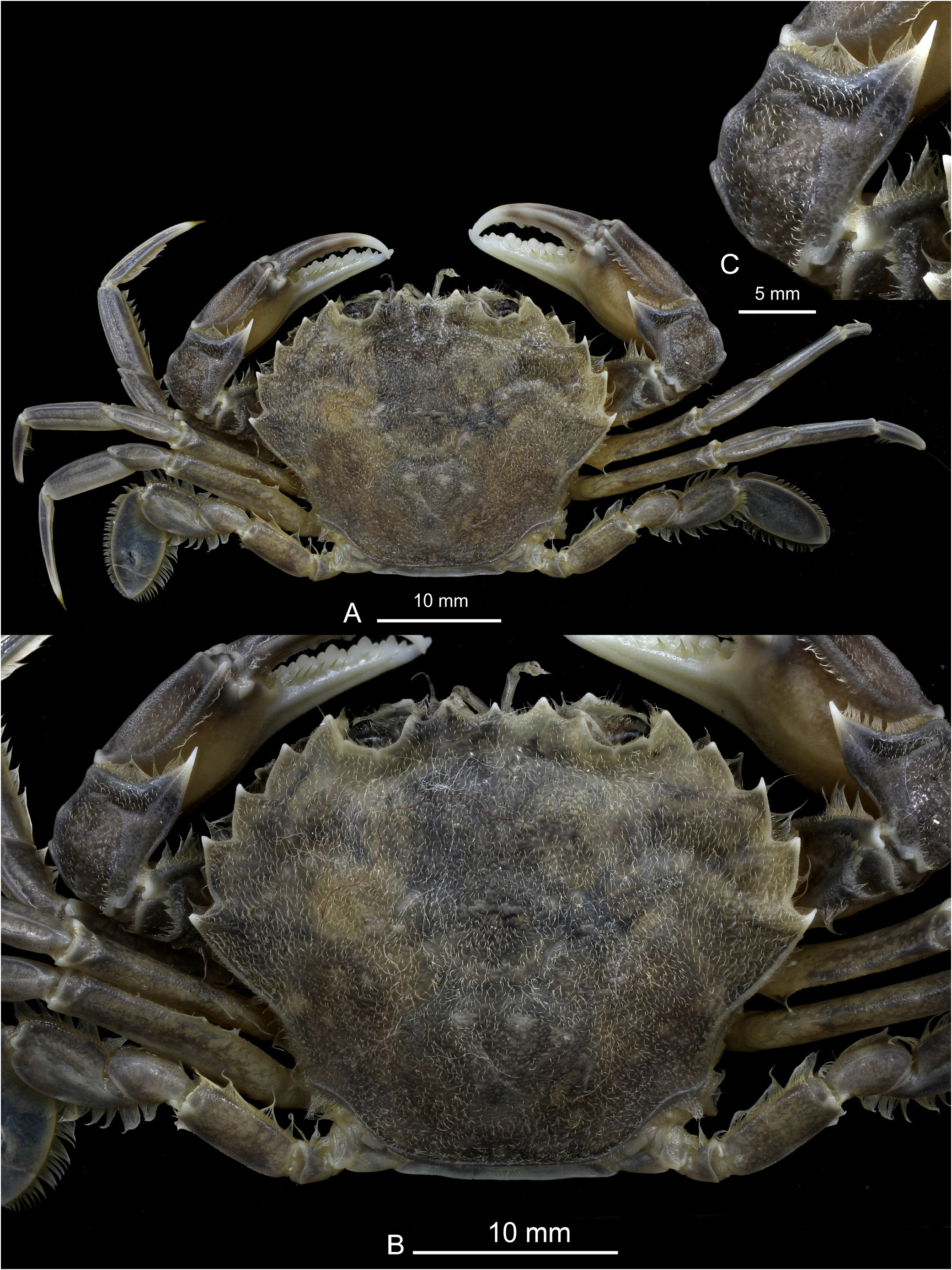
HABITUS. Carapace glabrous, wider than long (length/width relationships, CL/CW = 0.75 to 0.84 (holotype: 0.78) ( Figs 3A View Fig , 4A View Fig , 5A View Fig ), somewhat domed (convex), nearly smooth, but with some small tubercles especially located in the protogastric areas (in small aggregations) and in the anterior regions of the branchial areas (forming striae). Frontal region flat, with three slightly protruding blunt triangular teeth ( Figs 3A, F View Fig , 4A View Fig , 5A View Fig , 6A, C View Fig , 7B View Fig ), with a tuberculate rim, continued by the inner-orbital angle ( Fig. 6A, C View Fig ). This region is antero-dorsally delimited by a clear band; the middle tooth is rounded, subacute and does not reach the same proportions as the lateral ones, which are more developed (wider and rounded). Relative distance between external orbital teeth (apex to apex) and the distance between lateral rostral teeth (apex to apex) = 3.21 to 3.56 in males (holotype: 3.23; mean: 3.38), 3.44–3.56 in females (mean 3.50). Orbital area circular, concave, dorsally with two short longitudinal incisions, the internal longer ( Figs 3A, F View Fig , 4A View Fig , 5A View Fig , 6A View Fig , 7B View Fig ); ventral face with a very pronounced outer incision and a well protruding antero-internal lobe. Anterolateral borders of the carapace ( Figs 3A View Fig , 4A View Fig , 5A View Fig ) well curved, with a length more or less similar than the postero-lateral ones (0.95 to 1.17), which are converging. Anterolateral edges with five teeth (including the external orbital), the fifth and the fourth being somewhat larger, the third and fourth with a rectilinear outer face. Ventrally, the anterior margins (anterolateral teeth region) are delimited by a dense row of short setae; with a fringe of setae behind the pterygostomial sulcus. Branchial regions slightly domed, like the protogastric and centrogastric ones; concave urogastric region (depressed); cardiac region, with two tuberculate protruding areas anteriorly (and sometimes with two smaller ones behind) and flattened intestinal region.
EPISTOME. With two inclined longitudinal keels, one on each side.
ANTENNULE (A1). With a broad transversely folded basal segment in a fossa below the frontal area, with the ventral edge showing a projecting lobe located towards the third of the outer half.
ANTENNA (A2). Basal segments narrow, with a flagellum of about 26 segments, longer than the orbital concavity, reaching the basal half of the first anterolateral tooth (outer orbital).
CHELIPEDS (P1) ( Figs 3A–C View Fig , 4A–C View Fig , 5A, D View Fig ). Subequal. Propodus with a rounded ventral rim on its outer face, extending from the apex of the fixed finger to approximately the middle of the palm. Outer face of the palm with 3 longitudinal keels along entire length (one towards the middle zone and two in the dorsal zone, slightly more separated from each other, the upper one ends in the propo-carpal articular condyle); dorsal edge with a smooth rounded keel ending in a blunt distal tooth; internal face without keels, with the central area bulging (longitudinally) leaving two depressions (upper and lower). The palm (without the fixed finger) is 1.7 times as long as high and 1.48 times as long as the dactyl. Dactyl (mobile finger) with the distal part curved, externally presenting a central longitudinal depression, between two rounded and projecting ridges; dorsally, three rounded longitudinal ridges can be seen: an upper external one (which is the upper rounded edge of the external face), a central dorsal (somewhat more protruding, especially in its basal part) and an upper internal one, with depressions between them; inner face with a central longitudinal depression delimited by two rounded edges. Carpus ( Figs 3C View Fig , 4C View Fig , 5D View Fig ) with one strong and protruding tooth on antero inner dorsal margin; on the anterior outer upper part (coinciding with the superior articulation with the propodus) there is one well-marked and rounded projection and two others below: a not very prominent middle one and a bit more prominent posterior one. The external face ornamented with slightly raised areas and soft depressions. Merus smooth, with short setae along the entire upper edge (less in the most distal area) and inner edge of lower face (more developed in the distal area).
WALKING LEGS. Morphometric values: merus length (ML): P3 ≈ 2 ≈ 4> 5. The ML/MH (merus length/ height) ratio of P2–P3–P4 is between 2.9 to 3.1 and about 1.8 in P5. The ratio MLP2/P5, MLP3/P5 and MLP4/P5 is 1.82–1.88–1.8, respectively. Propodus length (PL): P3 slightly> P4 slightly> P2. Dactylus length (DL): P2 somewhat> P3 ≈ P4. The dactylus of all walking legs (P2 to P4) are longer than their respective propodus.
SECOND PEREIOPODS (P2). Merus smooth, with setae along the basal half of the dorsal margin, ventral margin without setae. Carpus and propodus smooth. Both segments with two dorsal ridges. On the propodus, external (anterior) face delimited ventrally by a row of very short setae, not extending onto carpus, and another row on internal (posterior) face, both separated by a dorsal longitudinal depression (wider in carpus). On the ventral edge of both segments, and along their entire length, there is a dense row of short setae. Inner face of carpus smooth and that of the propodus with a slight central longitudinal depression at anterior part. Dactylus of P2 quite rectilinear dorsally, and morphologically similar to those of P3 and P4, but wider, robust and with a slightly more curved tip; with a more or less quadrangular section, and with angles defined by rounded longitudinal edges, which leave a central longitudinal depression on dorsal, ventral and lateral sides (external and internal). Two dense rows of short setae are present on the anterior (external) face: one ventral (on the ventral edge) that exceeds half the length of the dactyl, up to approximately ⅔ basal, and another dorsal, somewhat shorter (on the dorsal edge) row, which runs from the basal part of the dactyl to half or little more.
THIRD AND FOURTH PEREIOPODS (P3 and P4). Similar to the second one (P2) and to each other. Both present setae on the basal half (P3), or on the entire (P4) dorsal (or superior) edge of the merus, and are glabrous on the ventral edge. Carpus and propodus smooth (without setae on the dorsal and ventral faces). The dactyli are morphologically similar to those of P2, but somewhat less robust and narrower (2 nd> 3 rd> 4 th). In P3 the row of short ventral setae goes from the basal part to half the length of the dactyl, or little more; while in P4 they are only found in the basal third (or a little more) ( Fig. 4D View Fig ), not reaching half of the dactyl, the row of dorsal setae is like that of P2.
FIFTH PEREIOPODS (P5). Merus smooth, with long setae all over the dorsal border, and in the distal fourth of the ventral outer part; ML/MH ratio: 1.63–1.85 (males), 1.76–2.0 (females). Smooth carpus, with long setae all over the dorsal and ventral edge. Propodus and dactylus flattened, with long setae on all edges (dorsal and ventral). Dactyl strongly flattened, paddle-like, oval ( Figs 3A, D View Fig , 4A, D View Fig , 5A, C View Fig ) (2.11 to 2.37 L/W), ending in a point.
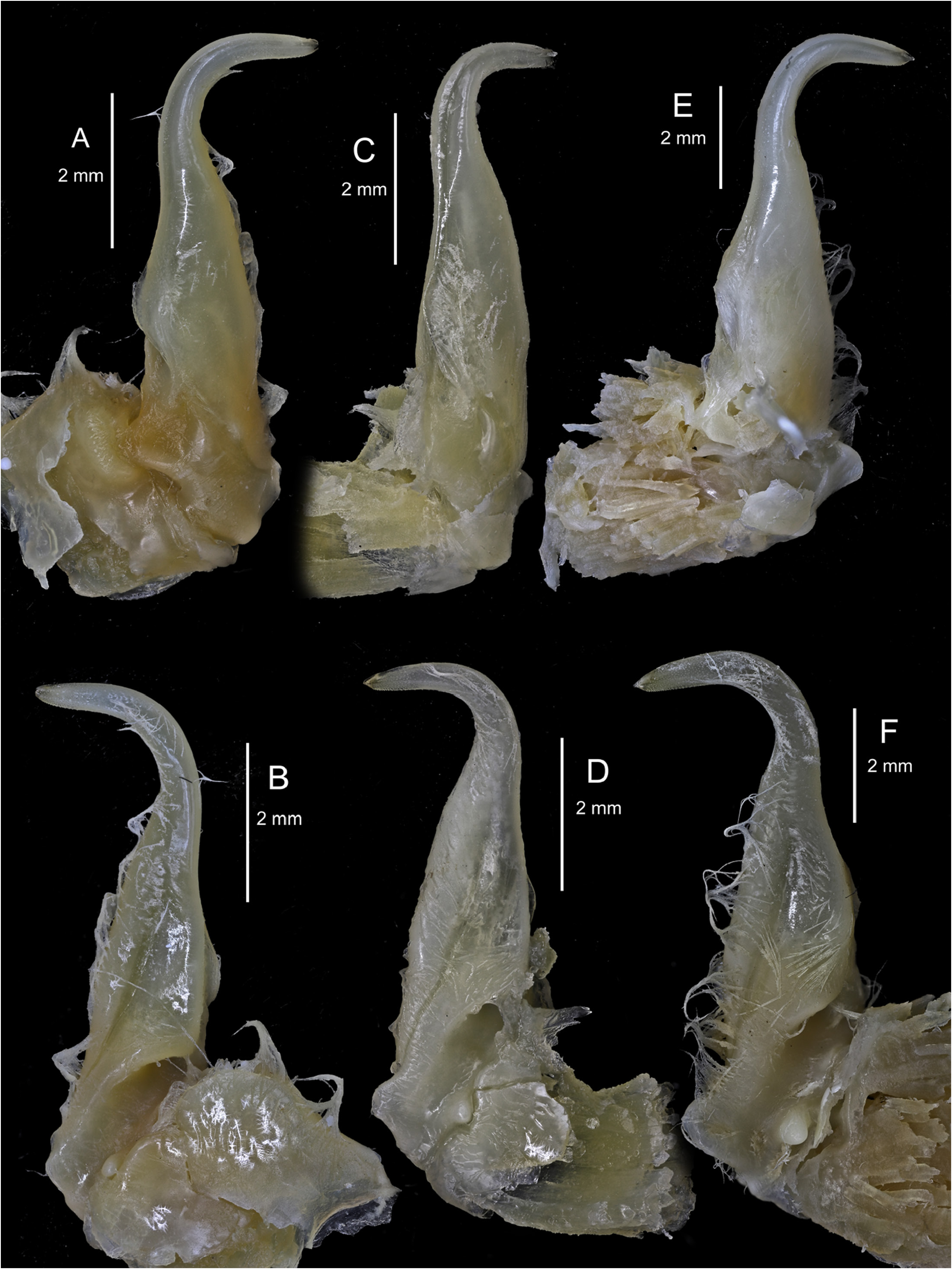
FIRST SEXUAL MALE PLEOPOD (GONOPOD). With curved distal part, which forms an angle of almost 90° ( Fig. 8C–D View Fig ), more curved in large specimen, and ends in a small point.
PLEON. Male pleon embedded in the sternum ( Figs 3E View Fig , 4E View Fig , 5C View Fig ), with the 3 rd to 5 th segments fused, narrowing strongly from the beginning of the 6 th and ending in a triangular telson. Rounded female abdomen.
COLOUR PATTERN ( Fig. 9 View Fig ). Body and legs usually greyish with a finely speckled motive; a large white, brown or black mark is occasionally present on the anterior part of carapace. Some specimens have a marbled colour pattern. The legs never exhibit the orange hue, which is usual in P. holsatus .
Distribution
Its exact geographical distribution is not well known, since in the papers (taxonomic, genetic and ecological) the identification of the species of Polybius is not at all clear (there is a lot of confusion between P. holsatus , P. marmoreus , P. vernalis and P. dioscurus sp. nov.). According to our data, it is present in the Atlantic Ocean: British Isles ( Ingle & Clark 1998; McCarthy et al. 2005; Ashelby 2006; d’Udekem d’Acoz 2011), the Netherlands ( Adema 1991), Belgium ( d’Udekem d’Acoz & Rappé 1991), Normandy ( Livory 1998), Bretagne ( d’Udekem d’Acoz 1986: fig. 10), Bay of Biscay (González Gurriarán & Ménez 1986), Portugal ( Nobre 1931, 1936; Neves 1975), Gulf of Cádiz ( López de la Rosa et al. 2002, as Liocarcinus vernalis ), Morocco: Bay of Agadir (Moukrim et al. 2010; fig. 2f), “ Maroc atlantique” (materiel examine by d’Udekem d’Acoz), Western Sahara ( Monod 1956 as Portunus barbarus ), Mauritania and Canary Islands ( Fransen 1991, as L. vernalis ?) and in the Mediterranean Sea: Alboran Sea, and Málaga (Marbella and Fuengirola) ( García Raso 1984 in part).
Habitat
It lives between 5 to 25 m, on bottoms of fine sand (range of median 0.17–0.32), with a low pelite content near the shore which increases to ca 10% at 20 m off Morocco, although at Oued Souss the pelite content was higher at a shallow depth (30% at 10 m). In Spain, it inhabits the Gulf of Cádiz on sandy bottoms with Callista chione ( Linnaeus, 1758) (0.5–10 m) and on fine muddy sand bottoms with Caulerpa prolifera (Forsskål) J.V.Lamouroux (3.3–6.6 m) and at Málaga it lives on sandy bottoms and has been caught during the harvesting of “coquina” ( Donax trunculus Linnaeus, 1758 ) at 2– 6 m. In Belgium, it lives on fine sand just below the tide marks and in shallow waters. So far, it has not been recorded in enclosed bays and environments of reduced salinity.
Remarks
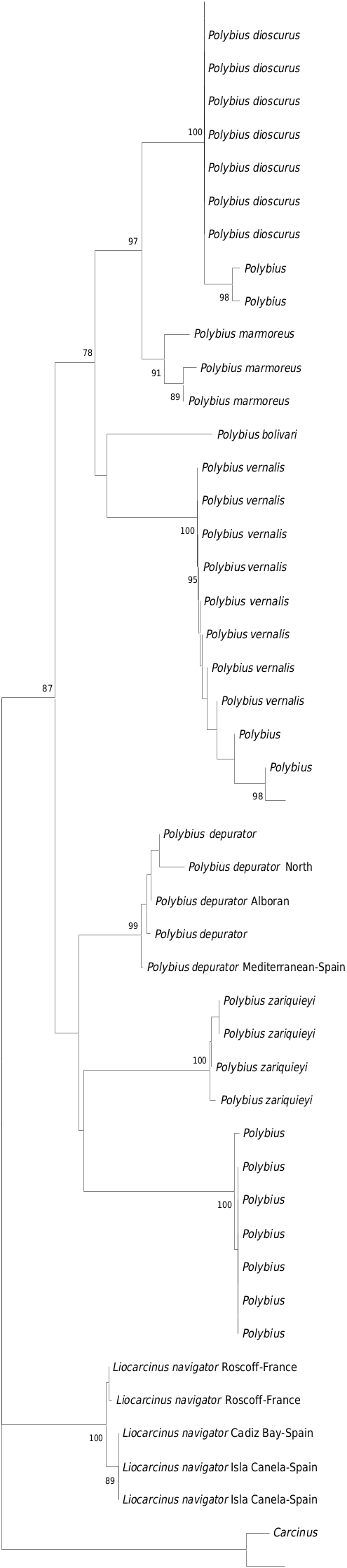
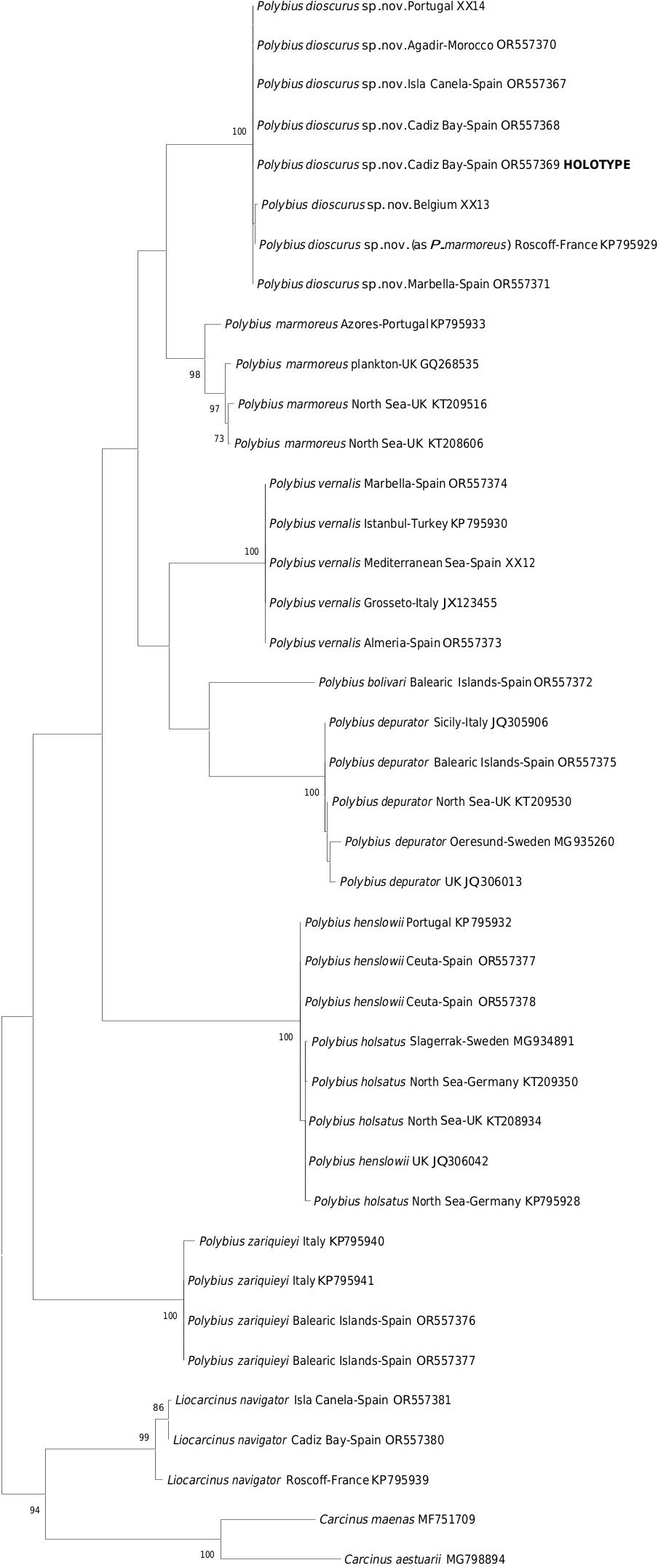
The new species, Polybius dioscurus sp. nov., belongs to the group formed by P. holsatus , P. vernalis and P. marmoreus , which are morphologically and genetically related ( Figs 1–2 View Fig View Fig ), but with significant differences. They have often been misidentified, because they share a combination of morphological characters, including some variability. Palmer (1927) analyzed the morphology of Polybius holsatus (as Portunus holsatus ) and compared it with that of P. marmoreus , showing a variability, which in some cases may correspond to characters of the new species. In 1986, d’Udekem d’Acoz raised the possibility that it was a new species, naming it Liocarcinus aff. holsatus , although he later considered it to be an Atlantic form of Polybius vernalis , with small population differences ( d’Udekem d’Acoz 1989). These considerations were not mentioned in later reports ( d’Udekem d’Acoz 1999), with the only exception of the existence of a glabrous versus a pubescent carapace. In the same broad period, d’Udekem d’Acoz & Rappé (1991) and Ingle & Clark (1998) carried out two very useful studies on the morphological differences between the three species considered valid in this group: P. holsatus , P. vernalis [in using material of P. dioscurus sp. nov.] and P. marmoreus (all as Liocarcinus ). This group was genetically validated by Passamonti et al. (1997), who also showed that there are some genetic differences among the populations of P. depurator and proposed the possible creation of subspecies. Ojeda et al. (2022) also cited differences between Atlantic and Mediterranean populations. Moukrim et al. (2010) captured specimens of the new species on bottoms of fine sand between 5 and 25 m, off Agadir ( Morocco), citing it as Liocarcinus cf. holsatus , since its morphology was considered close to P. holsatus , but still different.
This variability and the combination of morphological features is the reason for misidentifications, including records in GenBank and BOLD, where molecular sequences have been assigned to P. marmoreus , for example a specimen from Roscoff ( France) obtained by Plagge et al. (2016) (COI sequence KP795929) (see Fig. 2 View Fig ) and another from Portugal (sequence not available from BOLD, only early-release). Also, it has been assigned to P. holsatus , just like three specimens from Portugal (sequences not available from BOLD, only early-release); but all of them actually correspond to P. dioscurus sp. nov.
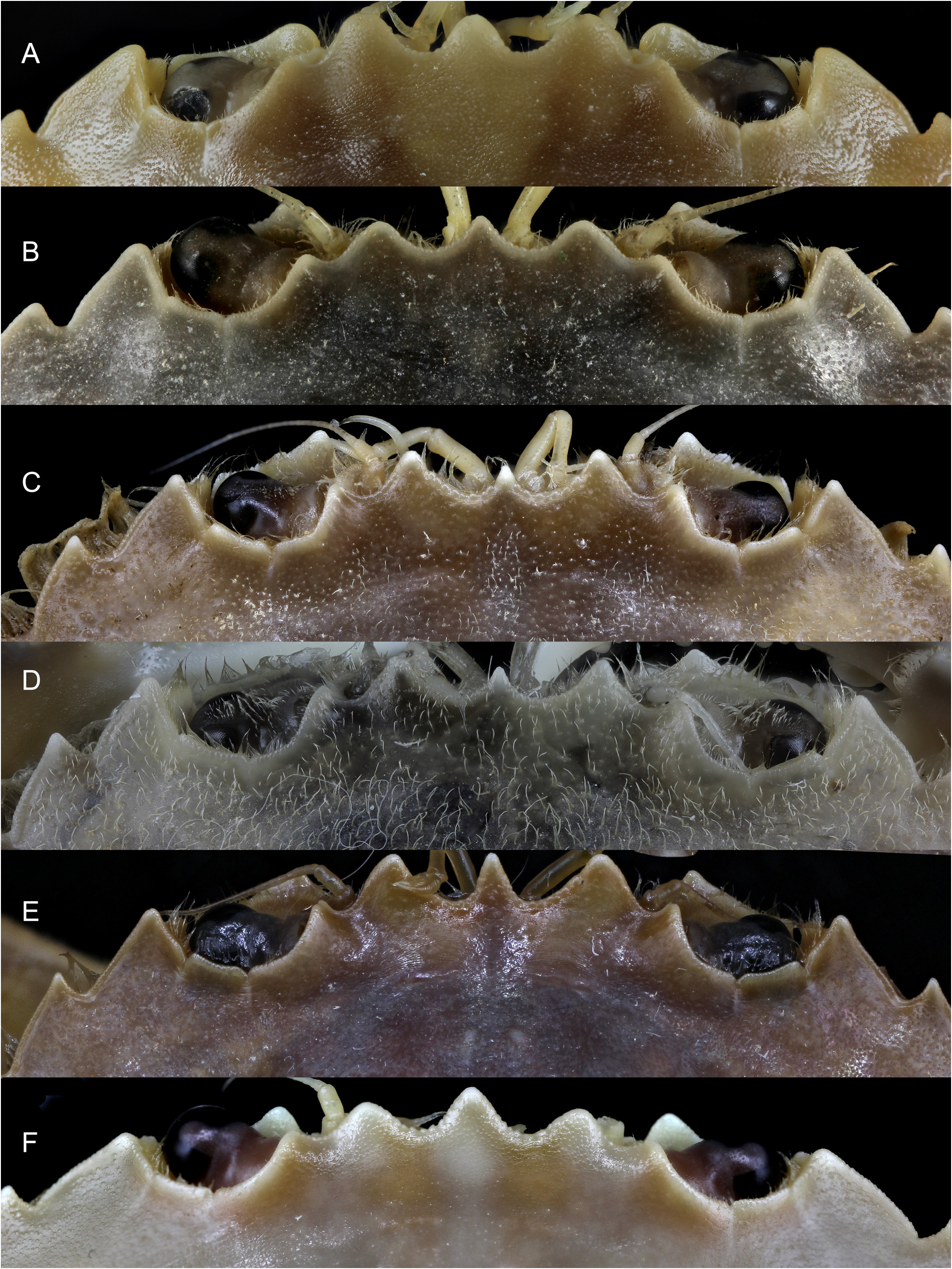
The rostrum of the new species ( Figs 3A, F View Fig , 4A View Fig , 5A View Fig , 7B View Fig ) is similar to that of P. vernalis , since the middle tooth is somewhat narrower either not, or only barely reaching the lateral ones, which are wider, rounded and more protruding; the central one is usually somewhat more rounded wide and somewhat longer in P. dioscurus sp. nov. ( Figs 6C View Fig , 7B View Fig vs 6D, 7C–E). In P. vernalis a greater variability is observed, mainly in specimens from the Black Sea in which the central rostral tooth is longer ( Figs 7E View Fig , 12 View Fig ). It differs from that of P. marmoreus because the middle tooth is similar to the lateral ones, rounded, and with equal length and width in this species ( Figs 7A View Fig , 13A View Fig ). In P. holsatus the median tooth overreaches the lateral ones ( Figs 7F View Fig , 14A View Fig ), as in P. henslowii ( Fig. 15A View Fig ). The rostral area of P. vernalis differs from that of P. dioscurus . The inner orbital angles are more projected in P. vernalis , while the curvature between the rostral teeth is more rounded in P. dioscurus (in P. vernalis the deepest zone is displaced towards the median tooth, not centred, Figs 6D View Fig , 7C–E View Fig vs 7B); however, there is some variability in this character. In addition, the tuberculate rostral rim is continuous with that of the inner orbital angle in P. dioscurus , while in P. vernalis it is not so clear because it is projected anteriorly and flexes slightly downward ( Fig. 6 View Fig ), and the inner side of the orbital socket shows a steeper slope in P. vernalis . The carapace in P. vernalis is frequently covered by short setae (e.g., Fig. 11 View Fig ), while in the others species it is glabrous, although we have seen specimens that were only almost glabrous. In addition, the carapace of P. vernalis from the Black Sea ( Fig. 12 View Fig ) is nearly smooth and hairless and their frontal teeth are a bit longer, different from those of the Mediterranean Sea ( Figs 10–11 View Fig View Fig ); but no genetic differences were detected, and the shape of their orbital region (the most important character separating them from P. dioscurus ) exhibits no difference in relation to the Mediterranean specimens. The anterolateral margin of the carapace is clearly curved, but in P. holsatus it is slightly shorter than the posterolateral one ( Fig. 14A View Fig ) (AB/PB: 1.17–1.23). In P dioscurus ( Figs 3A View Fig , 4A View Fig , 5A View Fig ) both are more or less similar in length (0.95–1.17) and in P. vernalis the posterolateral margin is slight shorter (1.07–1.17), but there are no clear differences between these two species. In P. marmoreus this ratio is 1.04–1.16. The anterolateral fifth tooth in L. vernalis projects slightly more outward than in P. dioscurus (with the tips pointing forwards) ( Figs 10A View Fig , 11–12 View Fig View Fig vs 3A, 4A, 5A). The posterolaterals borders are convergent, particularly in P. holsatus ( Fig. 14A View Fig ).
Other features that could be used are: the anterior outer border of the carpus of chelipeds, that in P. dioscurus sp. nov. shows protruding lobes that are slightly less marked than those of L. holsatus ( Figs 3C View Fig , 4C View Fig , 5D View Fig vs 14D), but closer to those of L. vernalis . However, in large specimens of P. dioscurus these lobes are less developed ( Fig. 4C View Fig ) (while in medium-small specimens they are more similar to those of P. vernalis ). In L. marmoreus the outer border is rounded and without any protrusions ( Fig. 13D View Fig ) (see Palmer 1927; d’Udekem d’Acoz 1986; d’Udekem d’Acoz & Rappé 1991). The ventral setae of dactyl P4 occupy approximately ⅓ basal in L. dioscurus and L. vernalis ( Fig. 4D View Fig ), while in L. holsatus they exceed half their length ( d’Udekem d’Acoz & Rappé 1991). In L. marmoreus they almost reach ½. However, this character remains somewhat variable. The merus of P5 is distinctly shorter in L. holsatus (ratio ML/MH: 1.1–1.3; 1.4–1.6 according d’Udekem d’Acoz & Rappé 1991) than those of P. dioscurus (1.63–1.89), P. vernalis (1.51–1.76) and L. marmoreus (1.65–1.87), whose ratio values overlap. D’Udekem d’Acoz & Rappé (1991) and d’Udekem d’Acoz (1991) found different ratios for the last two species (2.0–2.5, and 1.8–2.1, respectively). The datcylus and propodus are similar in P. vernalis and P. dioscurus but somewhat different from those of P. holsatus . The posterodistal lobe of the propodus is more developed and broader in P. holsatus than in other species ( Ingle & Clark 1998) and the dactylus is usually broader in P. holsatus .
The distal part of the first male pleopod is slightly more curved in P. vernalis ( Fig. 8E–F View Fig ; in larger specimens even somewhat hooked) than in P. dioscurus sp. nov., in which it tends to form an angle of approximately 90° ( Fig. 8C–D View Fig ).








| RBINS |
Royal Belgian Institute of Natural Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Polybius dioscurus
| García-Raso, J. Enrique, D'Udekem D'Acoz, Cédric, Moukrim, Abdellatif, Schubart, Christoph D. & Cuesta, Jose A. 2024 |
Polybius (Polybius) vernalis
| D'Udekem d'Acoz C. 1999: 222 |
Liocarcinus vernalis
| Martin J. 2011: 221 |
| McCarthy A. M. & McGrath D. & Allen B. M. 2005: 20 |
| Lopez de la Rosa I. & Garcia Raso J. E. & Rodriguez A. 2002: 87 |
| Ingle R. W. & Clark P. P. 1998: 224 |
| Livory A. 1998: 49 |
| Ingle R. W. 1996: 57 |
| Adema J. P. H. M. 1991: 111 |
| D'Udekem d'Acoz C. & Rappe G. 1991: 95 |
| Fransen C. H. J. M. 1991: 124 |
| D'Udekem d'Acoz C. 1989: 184 |
| Gonzalez Gurriaran E. & Mendez M. 1986: 43 |
| Garcia Raso J. E. 1984: 107 |
Macropipus vernalis
| Neves A. M. 1975: 32 |
Macropipus holsatus
| Cardona Bendito A. 1965: 152 |
Macropipus marmoreus
| Bourdon R. 1965: 29 |
Portunus barbarus
| Monod T. 1956: 173 |
Macropipus barbarus
| Forest J. & Guinot D. 1956: 37 |
Portunus marmoreus
| Nobre A. 1936: 36 |
| Nobre A. 1931: 66 |
| Palmer R. 1927: 889 |
| White A. 1857: 50 |