
Pseudophacopteron aspidospermi, Malenovský & Burckhardt & Queiroz & Isaias & Oliveira, 2015
|
publication ID |
https://doi.org/ 10.5281/zenodo.5303773 |
|
publication LSID |
lsid:zoobank.org:pub:1451DAF4-3F24-496C-86B5-3EEF4B9C5261 |
|
DOI |
https://doi.org/10.5281/zenodo.6401500 |
|
persistent identifier |
https://treatment.plazi.org/id/961D87B4-6C36-7B31-3018-AD1C6B97FAB7 |
|
treatment provided by |
Marcus |
|
scientific name |
Pseudophacopteron aspidospermi |
| status |
sp. nov. |
Pseudophacopteron aspidospermi View in CoL sp. nov.
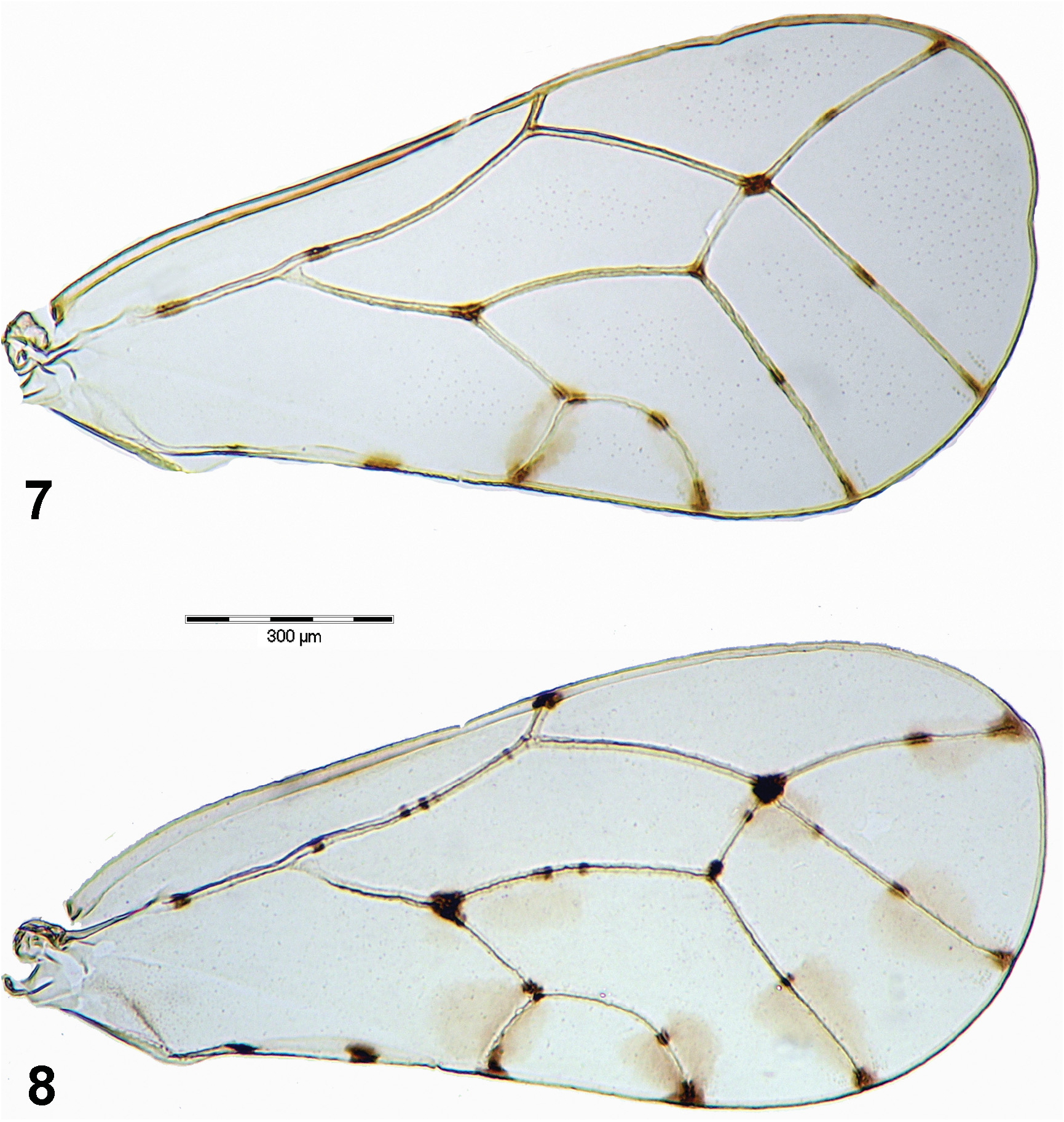
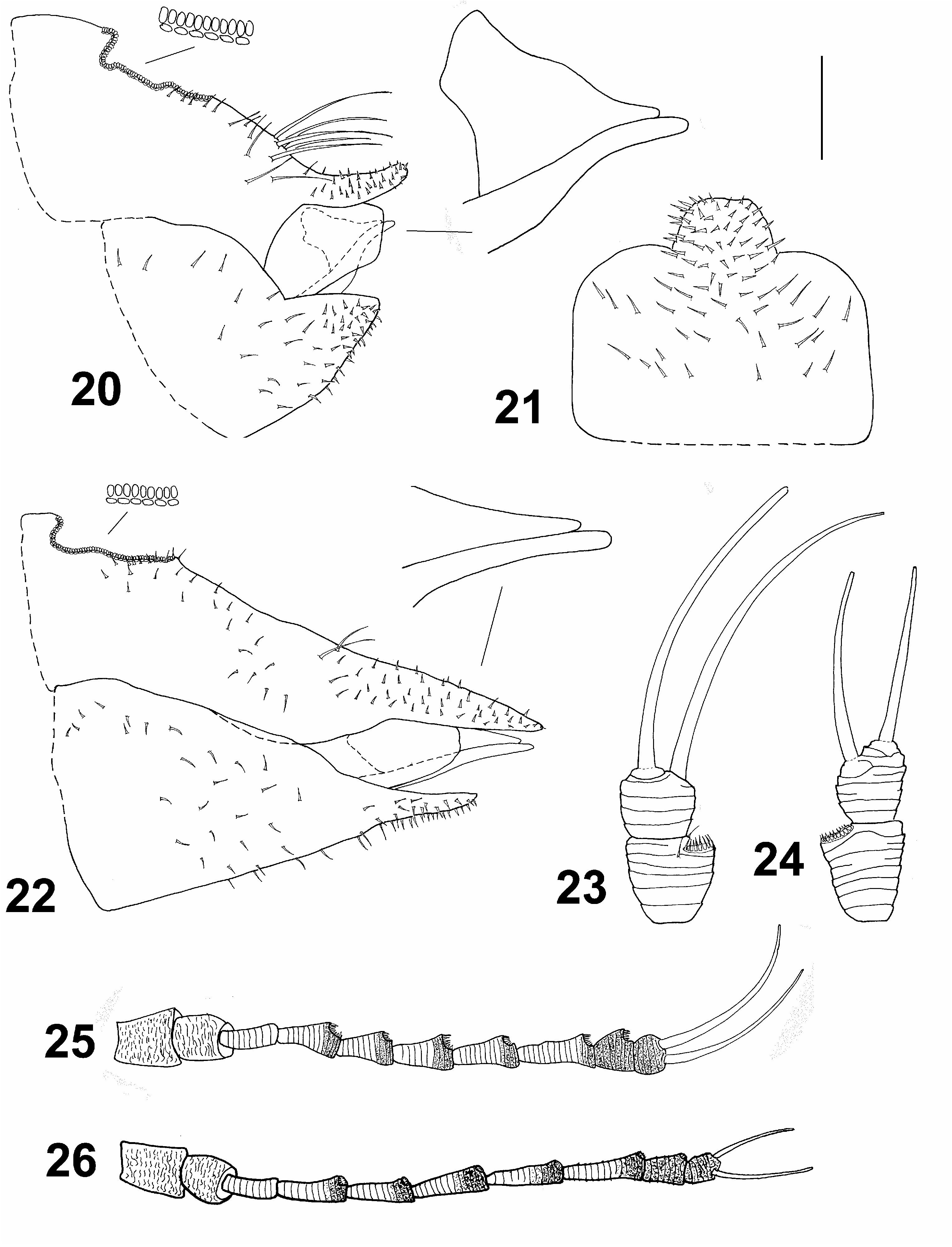
( Figs 1, 3, 5 View Figs 1–6 , 7 View Figs 7–8 , 9, 10, 12–16 View Figs 9–13 View Figs 14–19 , 20, 21, 23, 25 View Figs 20–26 , 27 View Fig , 29–35 View Figs 29–35 , 41–43 View Figs 41–43 )
“Psyllidengalle” [= psyllid gall]: RÜBSAAMEN (1907): 111.
Pseudophacopteron sp. : ISAIAS et al. (2000), CHRISTIANO (2002), CHRISTIANO et al. (2003), CAMPOS et al. (2010), OLIVEIRA & ISAIAS (2010), OLIVEIRA et al. (2011), BURCKHARDT & QUEIROZ (2012).
Type locality. Brazil, Minas Gerais: Belo Horizonte, Pampulha Campus of Universidade Federal de Minas Gerais, approx. 19°52′18″S, 43°52′00″W, 820 m a.s.l.
Type material. HOLOTYPE: ♂, BRAZIL: MINAS GERAIS: ‘ Belo Horizonte, Pampulha Campus of Universidade Federal de Minas Gerais , 12 April 1998, on Aspidosperma australe , J. C. S. Christiano leg.ʼ ( BMNH, dry-mounted) . PARATYPES: BRAZIL: MINAS GERAIS: 4 ♂♂ 6 ♀♀, same data as holotype ( BMNH, MMBC, NHMB, dry- and slide-mounted); 3 fifth instar immatures, same data but 17 June 2014; 2 fifth, 3 fourth and 3 third instar immatures, same data but 3 July 2014; 5 fifth instar immatures, same data but 10 September 2014, all R. M. S. Isaias leg. ( MMBC, NHMB, slide-mounted and preserved in ethanol); 1 ♀, Lavras, 21°14′S 45°00′W, 900 m, 1–6 June 2010, D. Burckhardt leg. #1 ( NHMB, dry-mounted). GoogleMaps SANTA CATARINA: 1 ♂, Nova Teutonia, 6 August 1943, F. Plaumann leg.; 1 ♀, same data but 29 June 1943 ( BMNH, dry-mounted).
Additional material examined. BRAZIL: RIO DE JANEIRO: Tijuca, July 1899, a twig of Aspidosperma australe with galls on leaves (coll. E. H. Rübsaamen, ZMHB, dry herbarium sample, Figs 41–43 View Figs 41–43 ).
Description. Adult. Coloration. Vertex dark brown with midline pale ochreous; genae, frons and clypeus uniformly dark brown ( Fig. 5 View Figs 1–6 ). Pronotum dark brown with midline and lateral tubercles pale ochreous; mesopraescutum orange brown with anterior margin dark brown or entirely dark brown with a pale ochreous patch at base medially; mesoscutum orange brown to dark brown with four pale ochreous stripes; mesoscutellum pale ochreous to dark brown; pleural sclerites dark brown with pale yellow markings ( Figs 1, 3 View Figs 1–6 ). Antenna off-white, segments 1–2 basally, segments 4–8 apically and segments 9–10 entirely dark brown ( Figs 1, 5 View Figs 1–6 , 25 View Figs 20–26 ). Legs off-white to pale ochreous; profemur with a dark brown transverse streak basally ( Fig. 5 View Figs 1–6 ), mesofemur with two dark brown streaks basally and subapically; metacoxa extensively dark brown on posterior and ventral margins, metafemur with a small dark brown patch on dorsal side subapically; metatibia with lateral and apical spurs dark brown; metatarsus infuscate ( Figs 5 View Figs 1–6 , 10 View Figs 9–13 ). Fore wing membrane clear with indistinct light brown infuscations along veins Cu 1b and apex of Cu 1a; veins off-white with well-delimited dark brown to black spots on apices and forks of all veins except R 1, one medial spot on each R+M+Cu 1, apical portion of Rs, M 1+2, M 3+4 and Cu 1a, respectively, as well as two medial spots on anal vein ( Figs 1 View Figs 1–6 , 7 View Figs 7–8 ). Hind wing membrane clear, vein C+Sc infuscate. Abdominal tergites orange brown, laterally and on posterior margin darker brown; sternites pale ochreous, dark brown laterally (especially on first visible sternite) to almost entirely dark brown ( Fig. 1 View Figs 1–6 ). Male terminalia uniformly pale yellow; female terminalia ochreous, apex of female proctiger dark brown ( Fig. 1 View Figs 1–6 ).
Morphology. Body with microsculpture, matt. Head, in frontal view, about twice wider than high ( Fig. 5 View Figs 1–6 ). Vertex dorsally with raised median ridge and two lateral bulges on either side in front; median epicranial suture completely reduced; lateral ocelli lying on small tubercles slightly above the plane of vertex. Eyes hemispherical; occiput and anteoccipital sclerite narrow. Genae small, weakly swollen, tubercle below torulus small and acute ( Fig. 5 View Figs 1–6 ). Antenna short, robust, serrate, segments strongly widening from base to apex ( Fig. 25 View Figs 20–26 ); one large elliptic rhinarium subapically on each of segments 4–9; rhinaria bordered with wreath of long cuticular spines; terminal setae subequal, more than twice longer than segments 9 and 10 together ( Fig. 23 View Figs 20–26 ). Fore wing ( Fig. 7 View Figs 7–8 ) pyriform, apex broadly rounded; costal break situated in distal fifth of vein C+Sc; membrane with fields of sparse surface spinulation in all cells except c+sc, leaving wide spinule-free bands along veins. Mesotibia with a comb composed of five stout setae on outer side subapically. Metafemur relatively long and slender, medially distinctly constricted, with a row of several evenly short setae along ventral margin ( Fig. 10 View Figs 9–13 ). Metatibia bearing open crown of ten tightly packed sclerotised spurs apically and two rows of seven and four similar, stout and sclerotised spurs laterally ( Figs 9, 10 View Figs 9–13 ). Metabasitarsus about 1.5 times longer than broad, conical, lacking sclerotised spurs laterally ( Fig. 10 View Figs 9–13 ). Abdominal tergites 3–4 with large dorsal tubercles ( Fig. 1 View Figs 1–6 ). Male subgenital plate with dorsal margin slightly convex. Male proctiger relatively slender, cylindrical ( Fig. 14 View Figs 14–19 ). Paramere shorter than proctiger; in lateral view, abruptly narrowing in apical fourth to a subacute apex; inner side bearing approximately 15 fine setae ( Fig. 15 View Figs 14–19 ). Distal segment of aedeagus with long shaft, apical dilation relatively short and broad, hooked; sclerotised end tube of ductus ejaculatorius relatively long and sinuate ( Fig. 16 View Figs 14–19 ). Female proctiger and subgenital plate with moderately long apical extensions covered with short and stout setae; dorsal margin of proctiger posterior to circumanal ring, in lateral view, strongly concave; circumanal pore ring with two rows of pores, pores of outer row contiguous; subgenital plate, in lateral view, with ventral margin convex, apex pointed ( Fig. 20 View Figs 20–26 ); in ventral view, broadly rectangular basally, with a truncate apical extension ( Fig. 21 View Figs 20–26 ). Dorsal and ventral valvulae lacking distinct lateral teeth ( Fig. 20 View Figs 20–26 ).
Measurements and ratios in Table 1 View Table 1 .
Fifth instar immature ( Fig. 27 View Fig ). Uniformly pale yellow, eyes grey. Body dorsally flat, ventrally strongly inflated. Body outline broadly ovoid, with convex lateral margins. Whole dorsal surface almost evenly covered with coarse microsculpture consisting of densely arranged small irregular (more or less round or square) cuticular granules; one larger quadrate cuticular scale on first visible abdominal tergite at posterior margin of hind wing pad on each side of midline. Whole body margin with pointed lanceolate setae in following numbers (one side only): head in front of insertion of antenna: 6–8, cephaloprothorax behind eye: 8–11, fore wing pad: 26–34, hind wing pad: 7–8, abdomen: (5–6) + (31–36); lanceolate setae absent from body dorsum except for a group of ca. 20 submarginal setae on each side of midline in subapical area of caudal plate. Antenna inserted on ventral side, oriented obliquely outwards and backwards over the body, extending beyond eye, gradually narrowing to a pointed apex, lacking distinct divisions and bearing two rhinaria posteriorly. Tarsal arolium membranous, hardly visible on slide-mounted specimens, sessile, pad-like, small, extending approximately to half of length of claws. Abdomen dorsally with four large free sclerites and caudal plate (incompletely fused in some specimens); caudal plate margin broadly rounded. Anus small, rhomboid, in ventral position. Circumanal ring moderately large, with fore and hind margins close together; outer ring composed of a single row of pores, hardly sinuate laterally. Measurements and ratios in Table 2 View Table 2 .
Egg ( Figs 12, 13 View Figs 9–13 ). Elongate, ca. 0.25 mm long, asymmetrical with subacute apex and reticulated surface.
Diagnosis. Adults of Pseudophacopteron aspidospermi sp. nov. are similar to P. longicaudatum sp. nov. in size and the absence of sclerotised lateral spurs on the metabasitarsus (the latter are well-developed in all other described Neotropical species of the genus). Pseudophacopteron aspidospermi differs from P. longicaudatum as well as from all the other known Neotropical Pseudophacopteron species in the robust antenna with very long terminal setae (more than twice longer than antennal segments 9 and 10 together), the dark coloured spurs on the metatibia, as well as the shape of the paramere, aedeagus and the female terminalia. It also differs from P. longicaudatum in the generally darker coloration of the body, particularly the head, the absence of oval light brown infuscations on the fore wing membrane on the apices of veins Rs, M 1+2, M 3+4 and the touching point of Rs and M 1+2, the smaller number of small dark brown spots on fore wing veins, the relatively broader fore wing with shorter apical section of Rs, the larger distance between apices of Rs and M 1+2, and the smaller (narrower) cell cu 1 of fore wing.
The fifth instar immature of P. aspidospermi generally resembles other Pseudophacopteron spp. living in intralaminar galls (see the Discussion). It is similar to P. longicaudatum in the unsegmented antenna oriented backwards and outwards over the body, the coarse microsculpture of the body dorsum, the presence of pointed lanceolate setae along whole body margin, and the structure of the circumanal pore ring. It differs from P. longicaudatum in the shorter and relatively broader body, the more evenly shaped and distributed granulous microsculpture on whole body dorsum, the more numerous and densely arranged lanceolate setae on the wing pad margins, the lack of isolated lanceolate setae on the abdominal dorsum, the presence of submarginal groups of lanceolate setae on the caudal plate subapically, the longer and less robust antenna, the larger circumanal pore ring, and the smaller tarsal arolium. Another species with similar fifth instar immatures are P. electum Capener, 1973 and P. sodalis Malenovský & Burckhardt, 2009 from Africa with which P. aspidospermi shares similar body shape, antenna, circumanal pore ring and pointed lanceolate setae on body margin but differs in the arrangement of the latter (both Afrotropical species lack the subapical groups of setae on caudal plate; P. electum also completely lacks lanceolate setae on cephaloprothorax and wing pad margins) and some other details ( MALENOVSKÝ et al. 2007, MALENOVSKÝ & BURCKHARDT 2009). The only other Neotropical species of the genus with described fifth instar immatures, P. punctinervis Brown & Hodkinson, 1988 , associated with Protium tenuifolium (Burseraceae) and known from Panama, differs from all the aforementioned species in the shape of the marginal lanceolate setae which are truncate, and the obliquely forwards-directed antenna with 3–5 indistinct divisions ( BURCKHARDT & BROWN 1992).
Etymology. Derived from the host plant genus Aspidosperma ; noun in genitive case.
Host plant and biology. Inducing intralaminar lenticular galls (sensu ISAIAS et al. 2013) on the leaf lamina of both young and mature leaves of Aspidosperma australe Müll. Arg. (Apocynaceae) ( Figs 29, 30 View Figs 29–35 ). Mature galls are green, glabrous, forming a slight projection of the adaxial surface and a more prominent projection of the abaxial surface of its host leaf, 5.0 ± 0.5 mm wide, with a central inner chamber lined with wax and enclosing one individual of P. aspidospermi from the first immature instar until the adult stage ( Fig. 31–35 View Figs 29–35 ). The gall is parenchymatic, with vascular tissues only at the top of the chamber, which is permanently opened through an ostiole to the abaxial leaf surface. Two tissue zones, an inner cortex, around the inner chamber, and an outer cortex with larger cells, can be distinguished within the gall ( ISAIAS et al. 2000, CHRISTIANO 2002, CHRISTIANO et al. 2003, OLIVEIRA & ISAIAS 2010, OLIVEIRA et al. 2011). Feeding of P. aspidospermi immatures alters the metabolism of the host plant during the gall development, particularly in both reserve substances (storage of starch in cells close to the inner chamber) and enzyme activity (related to carbohydrate metabolism and reactive oxygen species production), even though a nutritive tissue per se is not differentiated ( CAMPOS et al. 2010, OLIVEIRA & ISAIAS 2010, OLIVEIRA et al. 2011). Senescent galls open with a wide aperture on the gall surface ( CAMPOS et al. 2010; Fig. 43 View Figs 41–43 ). The galls can be frequent and numerous on the host; there are 5.35 ± 4.97 (mean ± standard deviation, n = 204) galls per leaf ( FERNANDES et al. 1988). Developing galls were observed over the entire year, indicating P. aspidospermi is a multivoltine species ( CAMPOS et al. 2010). In 2006, the highest percentage (80 %) of galled leaves was recorded in December, after the peak of leaf flushing, which occurred in September and when most galls were concomitantly induced on the young leaves. However, another peak of occurrence of galled leaves (77 %) occurred in July 2007, independently of new leaf production, which indicates P. aspidospermi might use mature leaf tissues as alternative oviposition sites ( CAMPOS et al. 2010).
Distribution. Brazil (Minas Gerais, Rio de Janeiro, Santa Catarina). The host plant, Aspidosperma australe , is widely distributed in the Midwest, Southeast and South of Brazil (Distrito Federal, Goiás, Mato Grosso, Mato Grosso do Sul, Minas Gerais, Rio de Janeiro, São Paulo, Paraná, Rio Grande do Sul, Santa Catarina), Bolivia, Argentina (Corrientes, Misiones), and Paraguay ( HASSLER 2014).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Psylloidea |
|
Family |
|
|
Genus |
Pseudophacopteron aspidospermi
| Malenovský, Igor, Burckhardt, Daniel, Queiroz, Dalva L., Isaias, Rosy M. S. & Oliveira, Denis C. 2015 |
Pseudophacopteron sp.
| BURCKHARDT & QUEIROZ 2012: 516 |
| OLIVEIRA D. C., ISAIAS R. M. S., MOREIRA A. S. F. P., MAGALHAES T. V. & LEMOS-FILHO J. P. 2011: 516 |
| CAMPOS P. T., COSTA M. C. D., ISAIAS R. M. D. S., MOREIRA A. S. F. P., OLIVEIRA D. C. & LEMOS-FILHO J. P. 2010: 516 |
| CHRISTIANO J. C. S., ISAIAS R. M. S. & KRAUS J. E. 2003: 516 |
| CHRISTIANO J. C. S. 2002: 516 |
| ISAIAS R. M. S., SOARES G. L. G., CHRISTIANO J. C. S. & GONCALVES S. J. M. R. 2000: 516 |
| OLIVEIRA & ISAIAS (2010) |
| RUBSAAMEN E. H. 1907: 111 |