
Pygopleurus brullei taygetanus Bollino, Uliana & Sabatinelli, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4674.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:B6626FE9-6490-4BC9-BACB-A4437AACDF25 |
|
persistent identifier |
https://treatment.plazi.org/id/03F087D8-C175-8401-FF0C-F919FE528ADD |
|
treatment provided by |
Plazi |
|
scientific name |
Pygopleurus brullei taygetanus Bollino, Uliana & Sabatinelli |
| status |
subsp. nov. |
Pygopleurus brullei taygetanus Bollino, Uliana & Sabatinelli View in CoL , new subspecies
( Figs. 99–105 View FIGURES 99–105 , 111–116 View FIGURES 106–114 View FIGURE 115 View FIGURE 116 )
Material examined. 199♂♂, 76♀♀:
Holotype: ♂, Greece – Lakonia / Taygetos, Pende Alonia Pass / ~ 36° 59’ N 22° 22’ E / 30.IV.2013 – m 1300 / Leg. Bollino, presently in MBLI, will be deposited in MHNG. GoogleMaps
Paratypes:
Laconia: Greece – Lakonia / 2 km SW Polovitsa / ~ 36° 55’ N 22° 26’ E / 29.IV.2013 - m 600 / Leg. Bollino ( 3♂♂, MBLI) GoogleMaps ; Greece – Lakonia / 5 km S Anavriti / ~ 37° 00’ N 22° 23’ E / 30.IV.2013 – m 1100 / Leg. Bollino ( 9♂♂, GoogleMaps 3♀♀, MBLI ); Recko / Anogia // J Kritzbach lgt. / 30.4.1992 ( 1♂, 1♀, IMC) ; Greece – Lakonia / Gorani, Moni Go- las / ~ N 36° 55’ E 22° 24’ / 10.V.2004 - m 800-1200 / Leg. Angelini ( 2♂♂, 1♀, MBLI; 8♂♂, 1♀, CMU) ; Pelopon- neso / Mt. Taigeto / 17.5.85 Drioli ( 1♂, 1♀, CMU) ; Peloponneso / Mt. Taigeto / 17.5.85 Drioli ( 2♂, 2♀, DCMT) ; Mt. Taigetos / 1400 m Greece // 1981.V.15-16 / leg. Rozner I. ( 3♀♀, IRCB) ; Greece – Lakonia / near Anavriti / ~ 37° 01’ N 22° 23’ E / 30.IV.2013 – m 800 / leg. Bollino ( 1♂, MBLI) GoogleMaps ; Greece – Lakonia / near Moni Golas / ~ 36° 55’ N 22° 25’ E / 29.IV.2013 – m 800-850 / Leg. Bollino ( 38♂♂, 6♀♀, MBLI; 3♂♂, 1♀, CGS; 2♂♂, 1♀, CMU) GoogleMaps ; Greece, Lakonia, near Paleopanagia / ~ 36° 58’ N 22° 24’ E / 28.IV.2013 - m 650 / Leg. Bollino ( 1♂, 2♀♀, MBLI) GoogleMaps ; Greece Peloponnese / ( Lakonia) Oros Taygetos / Paleopanagia env., // loc. Nihori m 650 / 36°58’36”N- 022°25’15”E / 8.V.2011 A Liberto leg. ( 1♀, CAL) GoogleMaps ; Greece Peloponnese / ( Lakonia) Oros Taygetos / Sohas , Pende Alonia , m 1340 / 36°58.99N 22° 22.59E / 29.IV.2013 A. Liberto Leg. ( 4♂♂, CAL) GoogleMaps ; Greece Peloponnese ( Lakonia) / Sohas - Pende Alonia pass / m 1340 2 V.2017 A. Liberto leg. / 36°58.99N 22° 22.59E ( 15♂♂, 3♀♀, CAL) GoogleMaps ; Greece Peloponnese ( Lakonia) / Sohas - Pende Alonia pass / m 1340 28 IV.2018 A. Liberto leg. / 36°58.99N 22° 22.59E ( 15♂♂, 8♀♀, CAL; 6♂♂; 1♀, CMU) GoogleMaps ; GR Peloponneso / ( Messinia) [recte: Laconia] Sochas env. / 27 IV. 2001 m 1000 / legit D. Sechi ( 1♂, 1♀, DSCC) ; Greece Peloponnese / ( Lakonia) Spartià env. / m 940 29.IV.2018 A. Liberto Leg. / 36° 53.42 N 22°23.98 E ( 18♂♂, 21♀♀, CAL; 4♂♂, 3♀♀, CMU) GoogleMaps ; Grecia Laconia / Taigetos Paleopanagia / 28/IV./ 2001 m.1100 legit. D. Sechi ( 1♂, CMU) ; GR., Peloponnese, distr. / Lakonia, Mt. Taigetos, / Anavristi , mixed forest, / 37° 2’10.49”N // 22°22’16.76”E, swept & / beaten, 13. V. 2013., leg. / Kotàn, Nemes, Gy. & T. Németh, Szénàsi ( 1♂, LNCB) GoogleMaps ; Greece – Lakonia / Taygetos, Anavriti / ~ N 37° 02’ E 22° 23’ / 7.V.2012 - m 700 / Leg. Agnoli ( 1♂, 2♀♀, MBLI) ; Greece / prov. Lakonia / Mts. Taygetos // 500-1000 m / Xirokambi / 25-27.V.2004 / leg. I & Gy. Rozner ( 1♂, IRCB) ; Greece / prov. Lakonia / Mts. Taigetos ( 900m) // Gorani / 2004 05 27 / Leg. V & G Rozner ( 1♂, IRCB) ; Grecia Lakonia, Taygetos / Gorani Gole del Monaco [=Moni Golas] / 800-1200 m 10.V.2004 / leg. Angelini ( 1♂, CFA-MZUF) ; Grecia, Lakonia / Taygetos, Kastania / 1200 m, 8.V.2004 / Leg. Angelini ( 2♂♂, 1♀, CMU) ; Greece – Lakonia / Taygetos, Pende Alonia Pass / ~ 36° 59’ N 22° 22’ E / 30.IV.2013 – m 1300 / Leg. Bollino ( 42♂♂, 3♀♀, MBLI; 4♂♂, CGS; 3♂♂ CMU) GoogleMaps ; Grecia Lakonia / Taygetos Vasiliki / ca 900 m 9.V.2004 / leg. Angelini ( 4♂♂, 6♀♀, CFA-MZUF, 1♂, 2♀♀, CGS; 1♂, 2♀♀ CMU) ; Greece / prov. Lakonia / Mts. Taygetos // 800 m Anavriti / Mon. Faneromiris / 28.V.2004 / leg. I & Gy. Rozner ( 1♂, IRCB) .
Diagnosis. Characters of the P. viridisuturatus species group described above, plus the following characters. P. brullei taygetanus new subspecies differs from P. brullei brullei new species in always having brown unicolored elytra (while commonly bicolored in brullei brullei new species); occasionally with a faint metallic shine observed only in the female, more evident on distal part. Paramera not diagnostic, shape of everted endophallus ( Fig. 104 View FIGURES 99–105 ) not different from P. brullei brullei new species.
Description of the male holotype ( Fig. 99 View FIGURES 99–105 ). Body size: 13.05 mm from the margin of the clypeus to the apex of the elytra; 15.4 mm including the apex of the abdomen. Width across the humeri: 5 mm
Color of integuments: head, pronotum and scutellum metallic, copper red. Elytra pale brown, with suture and lateral margin black. Propygidium black in the proximal part, orange in the distal one fifth; pygidium orange.Antennal articles first and second black, with black metallic reflection, the rest of the antenna blackish red. Legs metallic, with golden green shining.
Setation. Head with dense, erect, soft, yellow-whitish hair; black hairs mixed to light hairs on the canthus. First antennal article with dense black hairs, second with few black hairs. Pronotum and scutellum similar to the head, with black setae, longer and stouter than the light hairs, present at anterior angles of pronotum. Elytra with adpressed black hairs; long erect stout black setae are present along the lateral, apical and inner apical third of the edge; sparse long erect ivory yellow hairs, those in presutural area forming a row. Abdomen with hairs of axillary sclerites black, those along the edges of sternites ivory yellow; pre-pygidium and pygidium covered by ivory yellow adpressed hairs. Ventrally, mouthparts and prosternum densely covered by black hairs; mesosternum to pigidium, coxae and femora covered with yellowish hairs. Foretibiae with a row of black hairs on the dorsal surface. Mid and hind legs with long, ivory yellow hairs; spines light brown with lighter apex; apical spurs dark brown with lighter apex.
Morphology: clypeus subquadrangular, slightly narrowed basally, anterior angles rounded, without medial carina, but with a small notch in the center. Integument of the clypeus and of the rest of the head covered by fine and dense piliferous punctuation; microreticulation clearly visible through the punctuation on the whole head. Pronotum sub-ovoid, larger ( 3.8 mm) than long ( 3.1 mm),with anterior angles visible, rounded and obtuse; posterior angles rounded but readily visible; pronotal surface covered by vermiculated wrinkles, those along a 0.15 mm wide median sagittal line progressively more dense, thick and depressed toward the base. Scutellum triangular; about as long as wide, with a sculpturing similar to that of the pronotum. Elytra slightly dehiscent, more rounded at the external side than along the suture, apex regularly rounded, flattened after the apical callus; surface with a complete longitudinal presutural depression, and a short basal median one; piliferous punctuation fine, well visible near the humeral callus, fading toward the apex. Claws of prothoracic legs long and moderately curved, protarsi short (combs of tarsomeres 1 to 5 respectively 12, 9, 7, 4, 3 toothed). Claws of mesothoracic and metathoracic legs similar to that of prothoracic legs, but a bit larger and slightly slender. Mesotarsi 1.8 times longer ( 4.3 mm) than mesotibiae ( 2.4 mm).
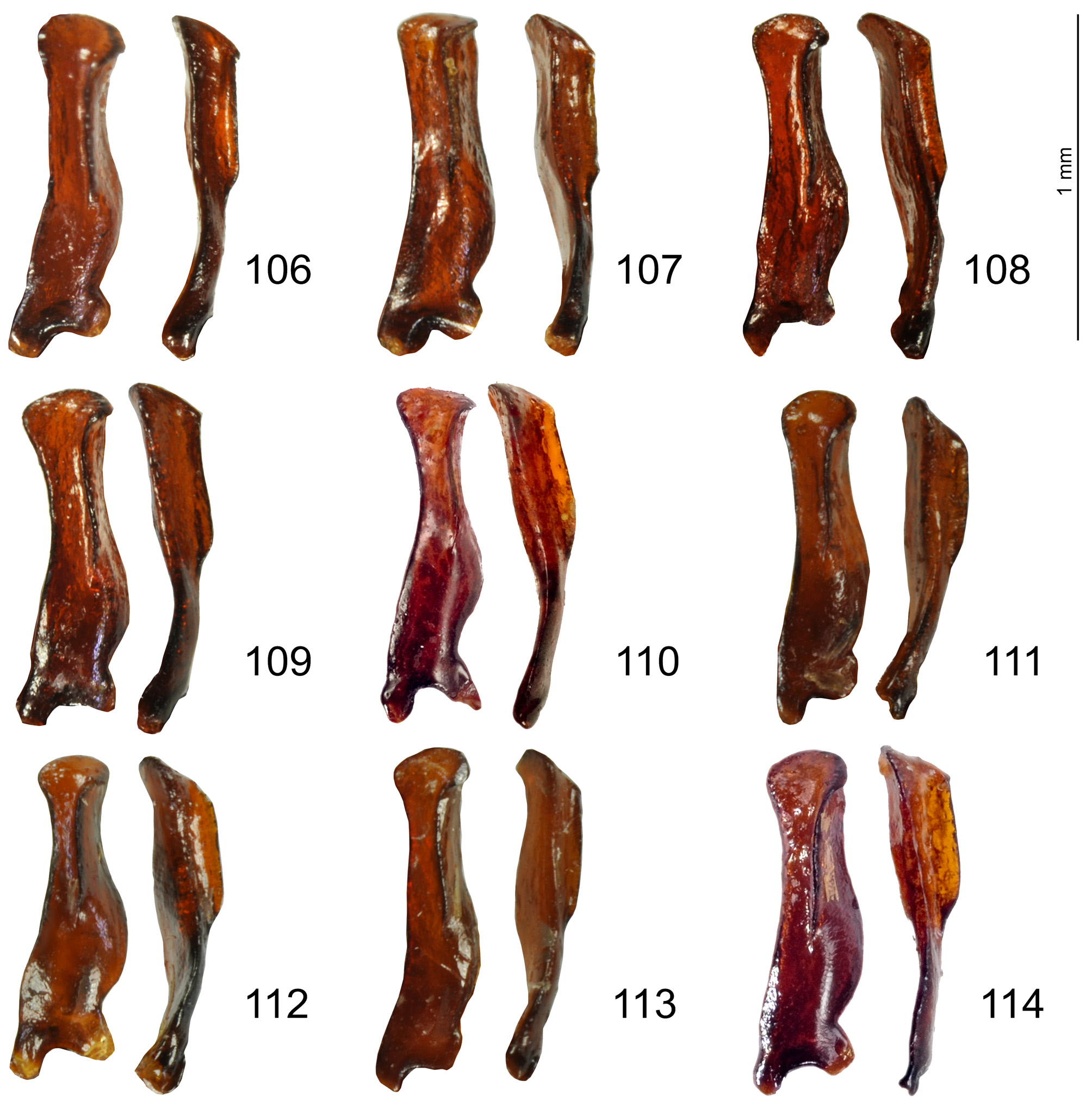
Paramera: Fig. 103 View FIGURES 99–105 .
Endophallus: Fig.104 View FIGURES 99–105 .
Relevant variability of males. None, except color of forebody going from green to copper-red or magenta-red.
Description of female paratypes. Body size: 11.5–13.1 mm from the margin of the clypeus to the apex of the elytra; 13.0– 14.3 mm including the apex of the abdomen. Width across the humeri: 4.5–5.0 mm.
Color: Androchrome, except elytra more shining than males, mostly with a diffuse, very pale cyan or green hue. Abdomen, including the pygidium, completely black, covered with ivory yellow hairs.
Morphology: similar to the male except the clypeus with a longitudinal carina, and Mesotarsi not longer than mesotibiae.
Genital sclerites: Fig. 105 View FIGURES 99–105 .
Etymology. The new subspecies is named after the mountain range of Taygetos in the Peloponnese peninsula, where the taxon is distributed.
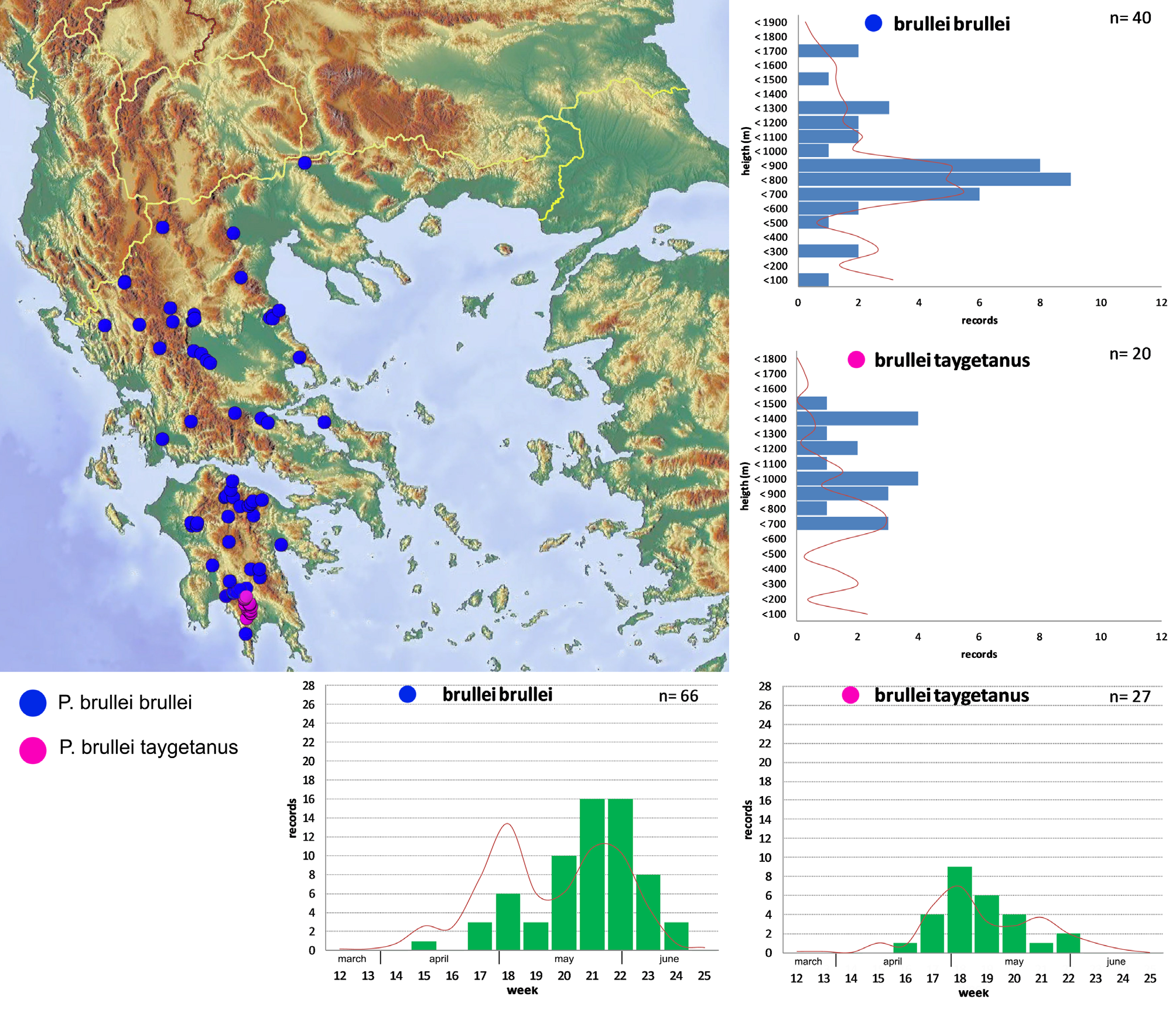
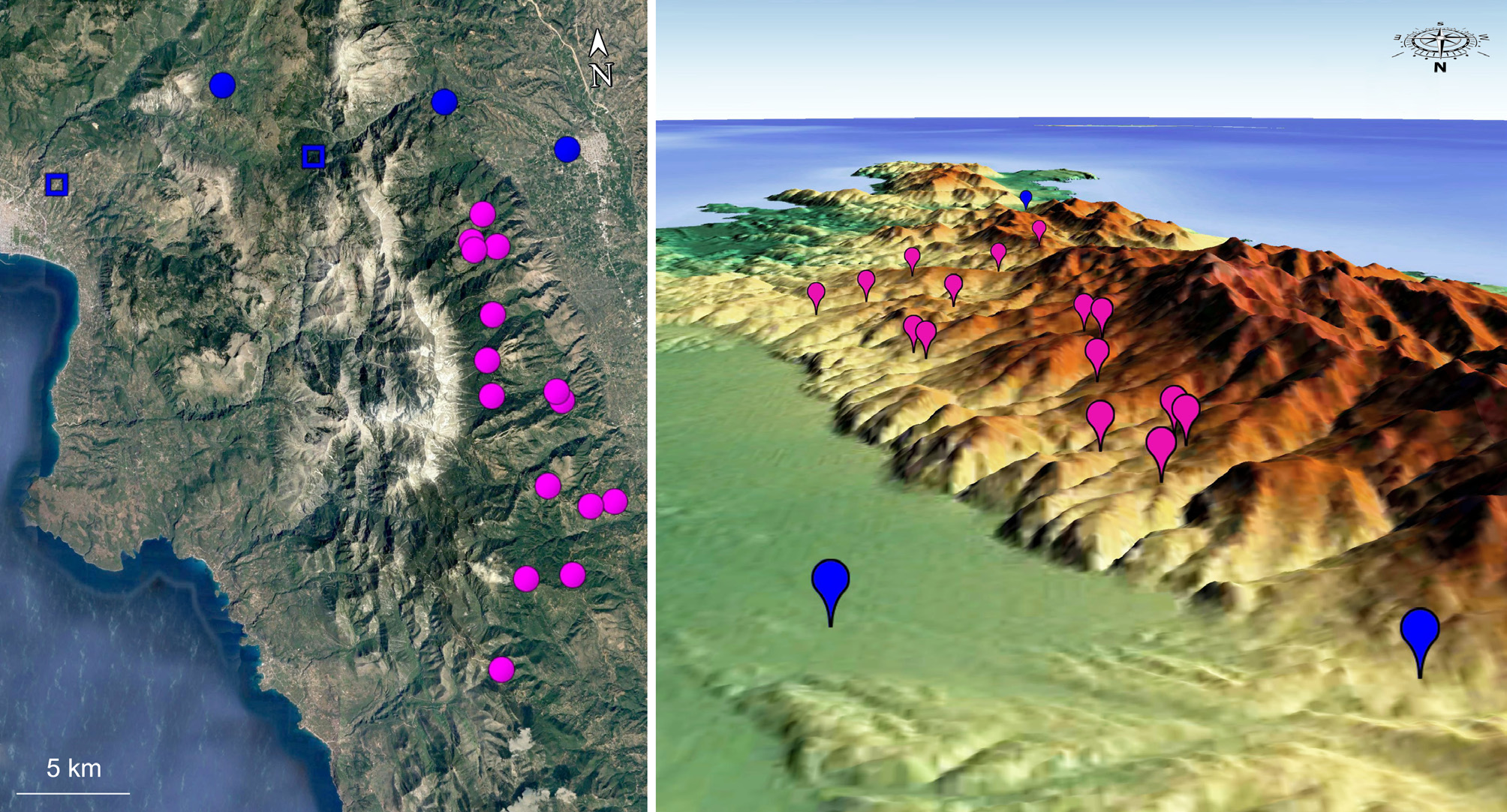
Distribution ( Figs. 115–116 View FIGURE 115 View FIGURE 116 ). Pygopleurus brullei taygetanus new subspecies seems restricted to the eastern slopes of the Taygetos mountain range, where it was recorded in several localities between the surroundings of Anavriti in the north and Kastania in the south. Its apparent absence from the western slopes of the mountain range may depend on lack of field sampling, or to the different biotopes apparently less suitable for Pygopleurus .
Eco-ethological notes ( Fig. 115 View FIGURE 115 ). The period of adult activity extends from the middle of April ( 19 th April, Sohas, Pende Alonia, 1340 m) to the end of May ( 28 th May, Monastery of Faneromenis, 800 m). The documented altitudinal range is narrower than that of the nominotypical subspecies, with an evident shift towards higher altitudes. Documented extremes are comprised from 600 m (SW of Polovitsa) to 1340 m (Pende Alonia Pass). Adults have a strong preference for yellow flowers of Asteraceae (Tubuliflorae) ; occasionally found feeding on yellow Ranunculus and Cistus incanus , while they are only very rarely found in Papaver .
Taxonomic remarks. Pygopleurus brullei sensu lato exhibits a relevant variability in two major external characters: color pattern of lateral hairs of abdomen, and color pattern of elytra (that is, presence and extension of the dark apical area).
Both characters vary quantitatively, and their variation is correlated to geography: outside the Peloponnese virtually all specimens have a well-defined bi-colored pattern on both abdominal hairs and elytra (figs. 86-88, 97), with scarcely significant variation. We examined a single specimen departing from this condition (fig. 93), having rather reduced elytral pattern and completely white abdominal hair. On the other hand, populations from Peloponnese vary significantly: within the same population, abdominal hairs can range from the “typical” bi-colored condition to completely white, including intermediate situations where dark hairs are progressively reduced towards the apex and mixed with white ones. The extension of the dark metallic area on elytra is variable in a similar way, and is often reduced to different degrees. Its disappearance is however uncommon, except on Taygetos, where its absence is observed on 100% of specimens. Populations from the Taygetos range are also characterized by abdominal hairs constantly white.
This situation suggests the existence of two separately evolving metapopulation lineages. We choose for them the rank of subspecies, rather than of species, as a gene flow is likely to happen between the two metapopulations. We infer this due to:
• the quantitative nature of the only diagnostic character identified by us (extension of the dark area on elytra), and the verified presence, although very rare, of specimens with the phenotype defining P. brullei taygetanus new subspecies in populations recognisable as P. brullei brullei new species
• the absence of morphological differences, including on paramera and everted endophallus, that would physically hamper hybridization.
• the strict geographic continuity
The two metapopulations do fit the two unanimously agreed conditions for the definition of subspecies, that is being (a) geographically defined and (b) diagnosable by at least one presumably heritable character ( Wallin et al. 2017), as we deem the presence/absence of dark elytral spots. In fact, the existence of other mountain populations exhibiting the ordinary bi-colored pattern suggests that the condition expressed by populations from Taygetos is not a case of phenotypic plasticity associated to mountain environment, but is rather linked to a different genetic (inheritable) condition.
Recognition as subspecies, based on inferred conditions above outlined, is therefore in agreement with (a) the definition of subspecies under the biological concept (we infer the absence of reproductive isolation), (b) with the definition of subspecies under the phylogenetic concept of species, as from the absence of reproductive isolation follows that P. brullei taygetanus new subspecies is not monophyletic with reference to nominotypical populations ( Patten, 2015), and (c) with the definition of subspecies under the unified species concept (USC), since they are potentially incipient species diagnosable by at least one heritable trait ( Wallin et al. 2017). The two taxa appears to fall within the “gray zone” for species delimitation under the USC (see de Quieroz 2007; Wallin et al. 2017), at least for the absence of reproductive isolation and for phenotipic distinction being partial (applicable to a large majority of individuals, but not to all).

| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Scarabaeoidea |
|
Family |
|
|
Genus |