
Sauropleura scalaris ( Frič, 1875 )
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad039 |
|
DOI |
https://doi.org/10.5281/zenodo.10267826 |
|
persistent identifier |
https://treatment.plazi.org/id/E554DE7B-F560-FFB8-D54E-7674FE5467F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Sauropleura scalaris ( Frič, 1875 ) |
| status |
|
Sauropleura scalaris ( Frič, 1875)
Urocordylus scalaris Frič – Frič 1875: 73.
Urocordylus scalaris Frič – Frič 1879: 187.
Dolichosoma angustatum Fritsch – Fritsch 1880: 117–119, fig. 63; pl. 21: 1, 3 non pl. 21: 2.
Urocordylus scalaris Frič – Fritsch 1881: 129–136, figs 76–81; pls 25, 26.
Ptyonius cf. pectinatus Fritsch – Fritsch 1895: 3 .
Ptyonius distinctus Fritsch – Fritsch 1901: 90–92; pl. 164.
Urocordylus scalaris Frič – Stehlík 1924: 242–248, fig. 8.
Urocordylus scalaris Frič – Steen 1938: 206–213, figs 1, 2A–C, 3A, B, 4A–C; pls 2: 4, 4: 1, 2.
Urocordylus angustatus (Fritsch) – Steen 1938: 213, fig. 2D.
Sauropleura scalaris (Frič) – Baird 1964: 14.
Sauropleura scalaris (Frič) – Milner AC 1980: figs 1A–C, 3B, 5A. Sauropleura scalaris (Frič) – Roček 1988: 525.
Urocordylus angustatus (Fritsch) – Roček 1988: 525.
Sauropleura scalaris (Frič) – Bossy and Milner 1998: 114, figs 53A, B, 54A, B, 55C, 58A, 59D, 60A, 61A, I–M, 64C, D, 66C, 67C, 68C, 69A, 71A, 72 non fig. 59C.
Sauropleurascalaris (Frič)–ŠtambergandZajíc2008: 182–183, fig.272. Sauropleura scalaris (Frič) – Marjanović and Laurin 2019: figs 31, 32.
Lectotype: NMP M458 View Materials , the original specimen, with both dispersed cranial and postcranial remains and its two galvanotypes.
Specimen referred here: ÚGV PAL00158, a complete skull with an articulated right mandible, partial presacral and nearly complete caudal vertebral column, ribs and right clavicle ( Figs 1–7 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 ).
Locality and horizon: Nýřany, 13 km south-west of Pilsen, Czech Republic. Canneloid shales and mudstones of the Main Nýřany Coal Seam, the Nýřany Member of the Kladno Formation (formerly ‘ Lower Grey Formation’ ) in the Pilsen Basin. Recent radioisotopic dating resulted in an absolute age of 308.3–305.9 Myr for the Nýřany Member, corresponding to the upper Carboniferous (Pennsylvanian) Moscovian stage, Asturian substage ( Opluštil et al. 2016).
Remarks on the taxonomic attribution: The following slightly modified character list is based on the diagnosis of Bossy and Milner (1998) and supports the reference of ÚGV PAL00158 to the genus and species S. scalaris . For the full diagnosis, see Bossy and Milner (1998). An asterisk denotes newly added characters.
The following unique combination of characters permits aưributionofthespecimentothegenus Sauropleura : (1) thenarrowly pointed snout composed primarily of elongated premaxillae and nasals; (2) the unpaired, narrowly waisted frontal bone; (3) the skull table narrow and its postorbital length is about one-and-ahalf times as long as its widest part across the supratemporals; (4) the cheek meets the skull table at an angle of ~50–60° from the horizontal plane; (5) the maxilla participates in the orbital margin; (6) the maxilla contacts the quadratojugal posteriorly; (7) the supratemporal is long and splint-like; (8) the recurved premaxillary teeth are significantly larger than the teeth of the maxilla; (9) the pterygoids are long and plate-like; (10) one pair of overlapping accessory apophyses above the zygapophyses of caudal vertebrae; (11) the distal neural and haemal spines of caudal vertebrae are triangular in shape and extend from narrow bases; (12) the haemal arches are about one-third of the total height of the caudal vertebrae, with neural spines of equal height to the haemal spines or slightly lower; (13) the four to five anterior-most caudal vertebrae are rib-bearing; (14) the tail is longer than the precaudal region and is composed of> 60 vertebrae; (15) the height of a caudal vertebra is about three times the maximum length of the centrum; and (16) caudal centra are relatively uniform in length and height to about the 30th vertebra, only diminishing thereasser.
The following unique combination of characters permits the aưribution of the specimen to S. scalaris : (1) the narrowly pointed and only slightly elongated snout; (2) the snout to mid-orbit length is ~40% of the total skull length; (3) the premaxilla with five large, recurved teeth; (4) the lower jaw is long and markedly slender anteriorly to angular, mandibular length equals six times its maximum depth; (5) the distally expanded fan-shaped neural spines of adjacent caudal vertebrae do not overlap; and (6) haemal spines are expanded distally and approach closely to almost contact each other.
The following characters are, among urocordylids, unique to S. scalaris , and therefore we consider them to be autapomorphic: (1) the short medial branch of the capsular ridge on the parietal ventral surface*; and (2) pits and grooves of the lateral line system developed on the mandible*.
Description
Skull
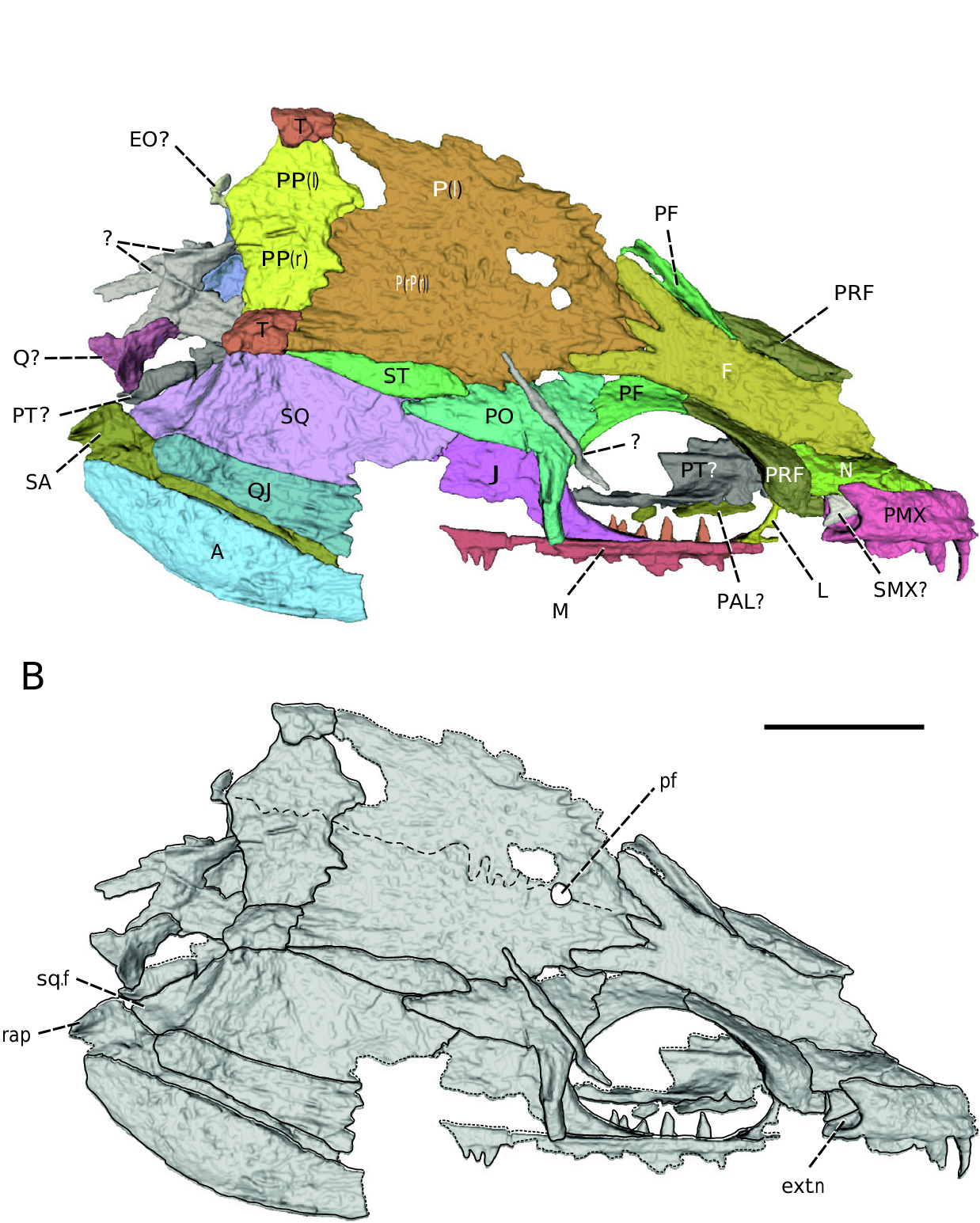
General aspect: The almost complete articulated skull is 15 mm long and heavily compressed in ventrolateral view ( Fig. 1A, B View Figure 1 ). Most of the dermal bones are present, although in some cases they are broken, and the overall shape can be inferred only from the partial imprints. Consequently, those parts could not be displayed through the µCT imaging, but imprints of their dorsal surface can still be observed on the skull in the ventrolateral view. Several bones on the ventral skull surface suffered from pyritic decay, and their original structure is no longer visible (e.g. maxilla, lacrimal, postorbital).
A
The dorsal skull roof is well preserved and shows only minor signs of deformation ( Fig. 2 View Figure 2 ). Its overall shape is subtriangular, with a narrowly pointed preorbital portion and wide, laterally expanded postorbital region. The length of the skull anterior to the mid-orbit represents ~40% of the total skull length. The widest part of the skull is behind the junction of the jugal and quadratojugal. From that point, the skull slightly narrows posteriorly for a short distance before reaching its strongly concave caudal border. Despite its current dorsolateral compression, the skull was apparently fairly high in life. As noted by Bossy and Milner (1998), the cheek region of Sauropleura slants laterally from the flat skull table at an angle of 50–60°, which was most probably true for the studied specimen. The orbit is large and circular, taking up one-quarter of the total skull length. The partly visible external naris was small compared with the orbits, suboval, and located close to the anterior margin of the orbit, far from the anterior tip of the snout.
The dermal sculpture is fine, most evident on the bones of the skull table (frontal, parietal, postparietal, supratemporal and tabular) and some elements of the circumorbital series (prefrontal, postfrontal and postorbital). It consists of small shallow pits and, to a lesser extent, elongated ridges and grooves. The second paưern is apparent especially on the quadratojugal and caudal area of the parietal. In other bones, the dermal sculpture
A is not well exposed, possibly because of the resolution limits of the µCT slices.
Premaxilla: The right premaxilla is well preserved and almost complete, whereas the imprint of the less element preserves only the posterior portion of the bone ( Figs 2 View Figure 2 , 3 View Figure 3 ). This elongated element forms, together with the nasal, the major component of the preorbital region of the skull. It is a relatively deep bone, dorsally contacting the nasal along a slightly oblique suture. Its posterior-most part is strongly concave and participates in the anterior border of the external naris. Two separate processes are developed at the caudal termination of the bone: the posterodorsal (or nasal) process and the posteroventral (or maxillary) process. The exact nature of the premaxilla–maxilla junction cannot be observed, because this area is not well preserved in the specimen. There are five irregularly spaced tooth positions on the premaxilla ( Fig. 3B View Figure 3 ). Only a single tooth is preserved completely, whereas others form natural moulds. The teeth are recurved, sharply pointed and rather large, with the third and fourth teeth being the largest in the row. The first tooth reaches only about half the height of the second one, which seems to be round in cross-section. The teeth are smooth and lack longitudinal grooves, i.e. labyrinthine infolding, as is typical for most other ‘lepospondyls’ ( Carroll et al. 1998).
Septomaxilla: We tentatively interpret the small crescent bone protruding in the lateral view from the anterodorsal border of the external naris as the septomaxilla ( Fig. 2 View Figure 2 ). Its location near the anterior, rather than posterior, border of the naris could possibly be explained by displacement owing to weak aưachment to the adjacent dermal bones, as was reported previously in some temnospondyls ( Schoch 2001, Witzmann 2005). The bone is subtriangular and smooth, and its concave margin would originally have been directed anteriorly.
Nasal: The right nasal is almost completely preserved and well exposed in the lateral view, whereas it is almost entirely overlain by the posterior portion of the vomer in the medial view ( Figs 2 View Figure 2 , 3 View Figure 3 ). This elongated, flat, slender element is one of the most significant bones contributing to the snout region of the skull. In lateral view, the anterior-most termination of the nasal extends slightly behind the rostral margin of the premaxilla to about two-thirds of its length. Posteroventrally, the nasal contributes to the dorsal border of the external naris. The caudal border of the nasal is strongly concave and contacts the prefrontal, and the nasal posteromedial process fits well between the anterior portion of the frontal and prefrontal.
Preflontal: Both prefrontals are present, but although the right prefrontal is nearly intact in lateral view, the less one is rather fragmentary and deformed ( Figs 2 View Figure 2 , 3 View Figure 3 ). The prefrontal forms the anterodorsal margin of the orbit and contributes to the posterodorsal edge of the naris. In dorsolateral view, the anterior portion of the prefrontal is slightly expanded and forms a rounded suture with the nasal. Its anterolateral section is rhomboid in shape and slopes ventrally, where it meets the lacrimal. The postfrontal process of the prefrontal is long and dorsoventrally flaưened. It sutures medially to the lateral edge of the frontal, whereas its lateral margin contributes to the sharp orbital rim. The suture with the postfrontal is V-shaped and placed approximately at the mid-level of the orbit.
Frontal: The frontal is a large, flat, unpaired dermal bone that occupies a substantial dorsal portion of the interorbital region ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is excluded from the orbital margin by the connection of prefrontal and postfrontal. In dorsal view, the anterior area of the bone is slightly widened and forms a large, subtriangular nasal process that protrudes between the posteromedial portions of the paired nasals. The frontal extends anteriorly, roughly at the level of the anterior margin of the naris. The smaller subtriangular naso-prefrontal process protrudes laterally from the less anterolateral area to contact the nasal and prefrontal. A similar process was probably also developed on the right side of the frontal, although it is broken off close to its base. The frontal is almost equally wide throughout its middle area. The posterior frontal expands laterally and forms three elongated, distinctly separated processes. The short posteromedial process is straight and wedged between the pointed frontal processes of the paired parietals. The posterolateral processes are much longer and reach the level of the posterior orbital edge. They contact the postfrontal and postorbital laterally, and posteriorly overlap the parietals strongly.
The ventral surface of the frontal is marked by a pair of conspicuous subparallel ridges, which are in the anterior area of the skull table termed the crista orbitotemporalis ( Figs 3B View Figure 3 , 4 View Figure 4 ; Gubin 1997, Damiani and Jeannot 2002). They emerge roughly from the level of the anterior orbital margin, immediately behind the anteriorly widest point of the frontal. At the mid-level of the frontal, they begin to diverge slightly from each other and continue onto the parietal bones posteriorly. The region between the orbitotemporal ridges is distinctively concave and probably accommodated the cartilaginous anterior portion of the braincase.
Postflontal: As with the prefrontal, both postfrontals are present, but only the right one is preserved well enough to enable a proper description ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is a boomerang-shaped bone that forms the posterodorsal margin of the orbit. Its dorsally flattened anterior process meets the prefrontal at the orbital mid-length, and together they form a V-shaped suture. The medial margin of the postfrontal is in contact with the frontal. Although only a small piece of the posterior portion of the bone is missing, the postfrontal was apparently, at least in part, overlapped dorsally by the anterodorsal process of the postorbital. As is seen in ventral view, the postfrontal is bent downwards posteriorly to meet the jugal in a short contact ( Fig. 3 View Figure 3 ).
Parietal: Both parietals are present and almost complete, although pieces of the broken bone are missing on the less side immediately behind the parietal foramen and posteriorly near the parietal–postparietal suture ( Figs 2 View Figure 2 , 3 View Figure 3 ). The plate-like parietals are the largest bones of the postorbital area and reach roughly the same length as the frontal. When joined together, their width exceeds that of the frontal. In dorsal view, each parietal forms a sharply pointed, subtriangular frontal process that is wedged between the medial and lateral parietal processes projecting from the caudal border of the frontal ( Fig. 2 View Figure 2 ). In ventral view, the anterior projections of the parietals are roughly quadrangular in outline and markedly underlie the posterior processes of the frontal in the same manner as was figured by Fritsch (1880: table 21; fig. 1) in ‘ Dolichosoma angustatum ’ ( Fig. 3 View Figure 3 ). The parietals extend laterally in the posterior direction along the medial postorbital margin slightly posterior to the level of the parietal foramen. Further posteriorly, they maintain a relatively constant width up to their caudal termination. The right parietal is laterally in contact with the postorbital in a complex stepped fashion, whereas its lateral contact with the supratemporal is only slightly curved and rather simple. The posterior area of the parietals is wing-like in shape and joins the postparietals in a strongly interdigitated suture medially. The posterolateral extensions of the parietals stretch further back to the firm sutural connection with the anterior margin of the tabulars. The parietal foramen is relatively small, rounded, and placed in the anterior quarter of the total parietal length. The interparietal suture is straight anterior to the parietal foramen, but posteriorly to it, it is strongly sinusoidal ( Fig. 2B View Figure 2 ). The caudal-most outline of the interparietal suture could not be traced on the µCT slices, but it was probably more or less straight (Bossy and Milner 1998). In ventral view, the suture is straight and only slightly undulated behind the parietal foramen ( Fig. 3B View Figure 3 ), similar to that of ‘ Dolichosoma angustatum ’ ( Fritsch 1880) .
The most distinctive feature of the parietal ventral surface is the paired orbitotemporal ridge, which continues from the posterior region of the frontal to the anterior area of the parietals ( Figs 3B View Figure 3 , 4 View Figure 4 ). The orbitotemporal ridges strongly diverge from each other at the level of the parietal foramen and run roughly along the midline of each parietal bone. A paired crista capsularis continues posteriorly to the parietal foramen ( Gubin 1997, Damiani and Jeannot 2002), where it is distinctively laterally expanded, and extends to the ventral surface of the tabular. A less prominent medial branch of the capsular ridge might be visible on the less parietal some distance behind the parietal foramen. It is short and disappears abruptly close to the interparietal suture ( Figs 3B View Figure 3 , 4 View Figure 4 ). The identical medial branch of the capsular ridge is present in ‘ Dolichosoma angustatum ’ (P.B., personal observation) and supports the previous reference of this specimen to S. scalaris ( Baird 1964, Bossy and Milner 1998).
Supratemporal: The right supratemporal is well exposed in dorsal view, but its ventral surface is almost completely overlain by a large, flaưened bone, the possible right pterygoid ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is a thin and long, splinter-like bone, about five times longer than its widest part. It is inserted between the large bones of the laterally situated squamosal and medially situated parietal. Anteriorly, it contacts the medial margin of the posterior process of the postorbital. Posteriorly, it tapers gradually to a thin strip of bone contacting the tabular, thus preventing the squamosal and parietal from contacting each other.
Postparietal: The paired postparietals are small, flat, polygonal bones contributing to the caudal margin of the skull table ( Figs 2 View Figure 2 , 3 View Figure 3 ). Although the exact course of the midline suture is poorly defined in the µCT scans, it was probably dorsally irregularly sinusoidal and, unlike the reconstruction of Bossy and Milner (1998), the bones were probably rather symmetrically developed. The anterior suture with the parietals is strongly interdigitated. The lateral joining of the right postparietal to the tabular is simple and straight, although the connection at the less side is rather concave. The caudal border of each postparietal is strongly concave, and together they form a wing-shaped structure exceeding the posterior margin of the lateral skull roof.
From the ventral view, only the less postparietal is visible, because the right one is covered by the bones of the braincase and palate ( Fig. 3 View Figure 3 ). It lacks a distinctly developed posterior occipital flange present in many early tetrapods ( Berman 2000). The posterior section of the postparietal bears a prominently protuberant, roughened ventral surface. A similar structure has been described in several other early tetrapods and probably represents the site for aưachment of the exoccipital or opisthotic (e.g. Bolt 1974, Dilkes 1990).
Tabular: Both tabulars are well preserved, but the right one is hidden from the ventral view by the overlapping elements of the braincase and palate ( Figs 2 View Figure 2 , 3 View Figure 3 ). The tabular is a small, subquadrangular bone forming the caudolateral margin of the skull table. It reaches only about one-half to two-thirds of the length of the adjacent postparietal. Its anterior margin broadly joins the posterolateral extension of the parietal, in addition to the posterior tip of supratemporal in the anterolateral point contact. Laterally, it meets the arched posterodorsal portion of the squamosal in a simple buư joint. A small protuberance extends from the caudal margin of the tabular beyond the adjacent lateral edge of the postparietal. In ventral view, the crista capsularis of the parietal continues posterolaterally to the tabular ( Figs 3B View Figure 3 , 4 View Figure 4 ).
Maxilla: Although incomplete, the middle and posterior portion of the right maxilla is preserved ( Figs 2 View Figure 2 , 3 View Figure 3 ). However, the anterior and posterior-most sections of the bone are entirely preserved as an imprint; therefore, they could not be displayed through µCT imaging. The maxilla is a long, slender, toothed element that contacts anteriorly the posteroventral portion of the premaxilla, and anterodorsally joins the small subtriangular lacrimal. It interrupts the connection between the lacrimal and jugal approximately at the mid-length of the orbit. In this region, the medial surface of the maxilla is overlain by the displaced small fragment of the less dentary ( Fig. 3 View Figure 3 ). The posterior-most termination of the maxilla is missing, and it is unclear whether it continued in a long, toothless, posterior process to contact the anterior margin of the quadratojugal, as reconstructed by Bossy and Milner (1998). In many specimens (e.g. NMP M391, NMP M865, ZP M122, ‘ Ptyonius distinctus ’), the maxilla clearly extends to the mid-length of the jugal, but its posterior portion is poorly distinguishable or seems to be absent. In ÚGV PAL00158, the maxilla displayed by μCT extends to two-thirds of the jugal length, although the crucial posterior-most part is missing. However, the imprint of an elongated rim extending along the ventral margin of the jugal is likely to represent the posterior-most portion of the maxilla contacting the quadratojugal. This region was probably fragile and thus broken off in many specimens. The posterior portion of the maxilla bears a flat, horizontal lamina for the reception of the jugal. All the maxillary teeth, with the exception of the three situated in the caudal-most position, are preserved only as imprints. The anterior teeth seem to be moderately large, but smaller than those of the premaxilla. The teeth become smaller and, possibly, more densely spaced in the caudal direction; otherwise, they conform to the premaxillary tooth positions.
Lacrimal: The right lacrimal is very incomplete and essentially preserves only its concave caudal margin contributing to the anteroventral portion of the orbit ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is a small bone, dorsally contacting the anteroventral portion of the prefrontal and ventrally siưing on the horizontal lamina of the maxilla. A deep depression is less in the place of the original bone and shows a roughly triangular shape of the element ( Fig. 1B View Figure 1 ). It could contribute, at least in part, to the posterior margin of the naris, as figured by Bossy and Milner (1998).
Postorbital: An almost intact right postorbital is well preserved on the dorsal skull surface in the original position and might be only slightly displaced ( Fig. 2 View Figure 2 ). It has a distinctive triradiate shape shared with other urocordylids (Bossy and Milner 1998). The subtriangular, anteromedial process of the postorbital overlaps the posterodorsal portion of the postfrontal. It also contacts the posterolateral process of the frontal, but it is unclear to what degree this connection is taphonomically influenced. Several subtriangular to semicircular lobes, located at the suture with the lateral margin of the parietal, project from the dorsomedial border of the postorbital. The posterior process of the postorbital is an elongated triangle. The pointed posterior end of the process contacts the supratemporal laterally and overlaps the squamosal anterodorsally and the jugal dorsally. The posterior process and the lateral stem of the postorbital form a sharp angle. The elongated, slender lateral process borders the posterior margin of the orbit and heavily overlaps the lateral surface of the jugal. It is unusually long and reaches up to the maxilla–jugal boundary, although this might not be the natural condition, owing to disarticulation. Unlike the rest of the bone, the lateral stem of the postorbital is smooth and not sculptured.
Jugal: The right jugal is essentially complete, but its posterior half is preserved only in the form of a bone imprint and therefore can be observed only on the ventrolateral side of the skull ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is a moderately large bone of the cheek region, contributing to the posteroventral margin of the orbit. Its anteroventral maxillary process is triangular in shape, with a sharply pointed anterior tip reaching halfway along the orbital length and resting on the horizontal lamina of the maxilla. In medial view, the anterior margin of the jugal is formed by a thick orbital rim, which is most pronounced at the contact with the maxilla. The dorsal process of the jugal is tongue-shaped and strongly underlies the ventral surface of the postorbital. The posterior and ventral portions of the jugal are preserved only as imprints. They rest on the horizontal lamina of the maxilla ventrally and contact the quadratojugal by the elongated caudal process posteriorly. Posterodorsally, the jugal is in contact with the squamosal along the slightly curved suture.
Squamosal: The squamosal is one of the largest bones of the cheek region and is well preserved on the right side of the skull ( Figs 2 View Figure 2 , 3 View Figure 3 ). The bone is roughly subtriangular in shape in the lateral view. Its posterodorsal edge is rounded and joins the tabular in a simple buư joint. This region is medially overlapped by the right pterygoid and cannot be observed in medial view ( Fig. 3 View Figure 3 ). The dorsal margin of the squamosal is almost straight along the suture with the supratemporal, and its pointed anterior tip fits between the posterior processes of jugal and postorbital in medial view. This anterior-most section of the squamosal is overlapped laterally by the posterior process of the postorbital. Its lateral surface slopes at an acute angle from the horizontal plain of the skull table and meets the dorsal border of the jugal and quadratojugal in a moderately curved suture. This connection is hidden from the medial view by the slightly displaced posterior portion of the mandible. The squamosal forms the caudolateral, concave border of the skull roof, and this prominent posterior edge delimits a strongly developed squamosal flange from the more anterior section of the bone ( Fig. 2 View Figure 2 ). The gently concave, smooth, subtriangular flange of the squamosal would form an incipient growth stage to the large extensions protruding from the caudolateral margin of the cheek region in larger specimens ( Steen 1938, Bossy and Milner 1998).
Quadratojugal: The complete right quadratojugal is elongated and contributes to the posteroventral margin of the skull ( Fig. 2 View Figure 2 ). It gradually widens to a broad, fringed anterior edge, which would probably be in slight overlapping contact with the posterior process of the jugal, if completely preserved. The maxilla–quadratojugal connection was probably present in the anteroventral part of the bone, although it cannot be seen clearly in the specimen. The anterolateral surface of the quadratojugal is formed by elongated subparallel ridges and grooves that become less pronounced towards the posterior part of the bone.
Quadrate: The displaced right quadrate is almost completely preserved posteroventrally to the skull roof ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is a relatively small, subtriangular element closely associated with the quadrate ramus of the right pterygoid and placed immediately posterodorsally to its articulation with the glenoid fossa of the articular ( Fig. 3 View Figure 3 ). The most pronounced feature of the bone is that its ventral surface is divided into two separated quadrate condyles by a deep groove. In ventral view, they are obliquely oriented to the longitudinal plane of the skull roof and directed medially. The quadrate has developed a flaưened, dorsally directed flange, which would be wedged between the quadrate ramus of the pterygoid medially and the quadratojugal laterally in occipital view.
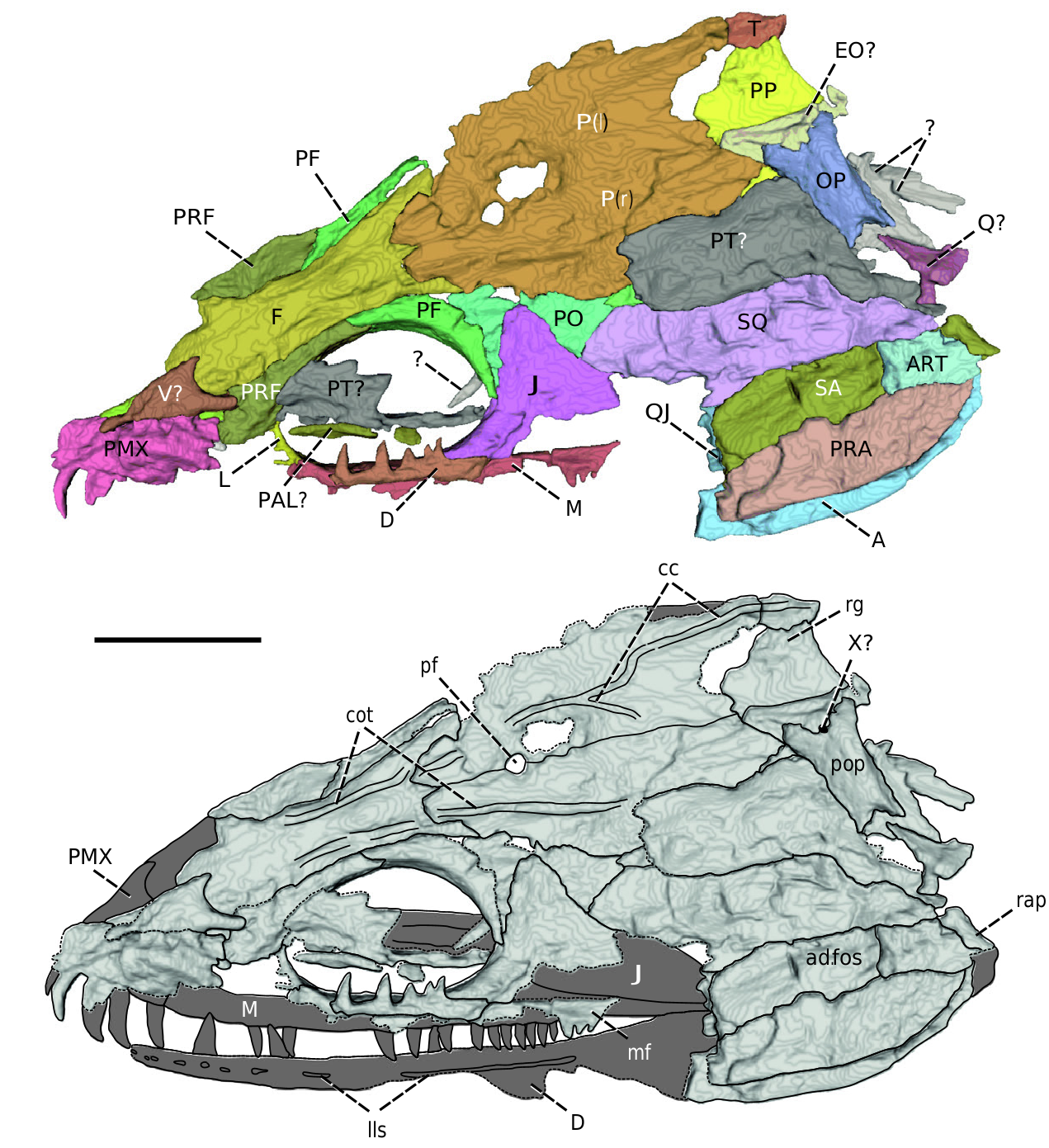
Vomer: A fragment of a bone overlying the right nasal in the ventral view is here interpreted as the possible posterior portion of the right vomer ( Figs 1B View Figure 1 , 3 View Figure 3 ). Its posterior-most section is well preserved and forms a symmetrical, strongly concave, crescentlike structure, which probably represents the anterior margin of the choana. Posteriorly, the vomer is divided into the shorter medial and a slightly more elongated lateral projection, the laưer with a rounded termination. However, the nature of the vomeral contact with other palatal bones remains uncertain.
Palatine: Fragments of a long and narrow bone that medially contacts the palatal ramus of the right pterygoid occur inside the right orbit ( Figs 2 View Figure 2 , 3 View Figure 3 ). Although the bone is almost featureless, based on its anatomical position and rough morphology it most probably corresponds to the palatine. Its anterior and posterior extent is incomplete, and no teeth or denticles can be observed from the µCT imaging.
Pterygoid: The right pterygoid appears to be preserved on the ventral side of the skull in two separated, fragmented parts ( Fig. 3 View Figure 3 ). The narrow anterior portion of the bone emerges from the orbital region and represents the palatal ramus of the pterygoid. In ventral view, it extends along the entire length of the orbit and laterally contacts the palatine. The abruptly terminated anterior portion is slightly curved medially and seems to be incomplete. The ventral surface of the palatal ramus is flaưened and lacks any certainly discernible denticles, which might, however, be too fine to be distinguished on µCT. The morphology of its dorsal surface is well exposed in the anterior section of the orbit in the dorsal view, and posteriorly in the form of an imprint from the skull ventral view ( Figs 2 View Figure 2 , 3 View Figure 3 ). The preserved medial margin of the palatal ramus is straight in the dorsal view and bears bulge-like thickening similar to that reported in an embolomere from Linton, OH, USA ( Witzmann et al. 2017). The central part of the bone is smooth and gently concave, whereas the lateral margin forms a sharp edge inclined at an acute angle from the horizontal plane.
In ventral view, the large, flaưened bone overlapping a large portion of the posterolateral skull roof is here less certainly interpreted as the posterior part of the right pterygoid ( Fig. 3 View Figure 3 ). Its anterior portion is incomplete and consists of a broad, rectangular base of the horizontal palatal ramus. The posteromedial part of the pterygoid immediately anterior to the opisthotic is bulged and might form the articulation place with the basipterygoid process, although none was found in the specimen. The long, slender quadrate ramus of the pterygoid extends from the posterolateral edge of the bone to contact the quadrate and appears to be deformed in shape. No ascending flange can be observed.
Exoccipital: Within urocordylids, the incomplete morphology of exoccipitals has been known only in Ptyonius Cope, 1875 until now (Bossy and Milner 1998), but a possible exoccipital is reported here, for the first time, also in Sauropleura . This element partly overlies the postparietals in ventral view and is closely associated with the right opisthotic ( Fig. 3 View Figure 3 ). It is subtriangular in shape and invades the medial head of the opisthotic in convex sutural contact. Its medial margin is straight and extends beyond the anterior and posterior terminations of the opisthotic. Posteriorly, the exoccipital forms a small, incompletely preserved projection surpassing the caudal border of the skull table, which is interpreted as a possible occipital condyle. The condyle is paired, as in all other nectrideans (Bossy and Milner 1998), and bears a flaưened posterior articulating facet.
Opisthotic: Among urocordylids, the opisthotic has been reported previously only for Ptyonius (Bossy and Milner 1998) and is here described in Sauropleura for the first time. It is well exposed in the posteroventral part of the skull roof, where it partly overlaps the right postparietal and several bones of uncertain identity ( Fig. 3 View Figure 3 ). In accordance with other nectrideans, it is a well-ossified and elongated element, with the expanded medial portion abuưing the exoccipital ( Steen 1938, Beerbower 1963, Milner 1996, Bossy and Milner 1998). The suture with the exoccipital is strongly concave and resembles that figured in Peronedon Olson, 1970 and Ductilodon Gubin, 1999 ( Milner 1996, Gubin 1999). A small jugular foramen for cranial nerve X, previously documented in few nectrideans ( Steen 1938, Beerbower 1963), might occur at the junction of the opisthotic and exoccipital in its central part. The paroccipital process of the opisthotic is elongated and robust. It would face dorsolaterally to join the roughened posteroventral surface of the postparietals. Unlike in Ptyonius , it tapers only slightly laterally before its abrupt V-shaped termination, which might have ended in cartilage.
Lower jaw: Only the right mandible is available for description, although a small, toothed fragment of the possible less dentary is preserved in the ventral margin of the orbit ( Figs 1B View Figure 1 , 2 View Figure 2 , 3 View Figure 3 ). The mandible is associated with the skull roof essentially in its original anatomical position. Only its deep posterior portion is somewhat displaced when seen from the lingual view and overlaps the posteroventral part of the squamosal, in addition to almost the entire quadratojugal ( Fig. 3 View Figure 3 ). The anterior two-thirds of the mandible is preserved only as a partial imprint of the labial surface, while the mineralized bone tissue of the posterior section enables it to be displayed through the µCT and description from both the labial and lingual views. The mandible overall is long and markedly slender anterior to the angular, whereas its posterior section is moderately deep. The length of the preserved portion is ~5.8 times its maximum depth at the level of adductor fossa. The mandible articulates approximately at the level of the caudal margin of the skull roof, but the conspicuously developed, posterodorsally directed retroarticular process extends posteriorly beyond it, although not as strongly as in urocordylines and diplocaulids (Bossy and Milner 1998, Milner 2019). No openings other than the adductor fossa are preserved in the specimen. It occupies about one-fissh of the total mandibular length. The dorsal margin of the prearticular forms the lingual border of the adductor fossa. It is set more ventrally compared with the dorsal edge of the surangular, which forms the labial border of the fossa. This gives rise to a prominent surangular crest and results in the dorsomesial orientation of the adductor fossa present in most other post-Devonian tetrapods ( Ahlberg and Clack 1998).
Dentary: With the exception of a displaced small fragment of the less dentary situated on the ventral edge of the orbit, the morphology of this bone can be inferred only from a partial imprint of the labial surface of the right element ( Figs 1B View Figure 1 , 3 View Figure 3 ). The anterior-most region of the bone is missing, as are its ventral part in the mid and posterior sections. The dentary, which gently widens posteriorly, is very long and slender, occupying ~67% of the length of the preserved mandible. The labial surface of the dentary is smooth and not sculptured, with the exception of the pits and grooves of the lateral line system (see below). The few preserved dentary teeth are smooth, pointed and slightly recurved, as documented by the toothed fragment of the less dentary. At least the anterior teeth are fairly large, although slightly smaller than the largest premaxillary teeth.
Angular: The angular is a large bone of the lower jaw well exposed in labial view ( Figs 2 View Figure 2 , 3 View Figure 3 ). It is almost complete, and only its anterior-most portion is broken off. In labial view, the angular forms the major component of the posteroventral area of the mandible. Its ventral margin is strongly convex and gives rise to the deepest part of the mandible. The angular dorsally contacts the surangular in an almost straight suture and exceeds its length more anteriorly, where it would probably have contacted the posteroventral portion of the dentary (Bossy and Milner 1998). The caudal border of the angular is rounded and seems to be complete, thus its contribution to the retroarticular process is not significant. The labial surface of the angular was certainly ornamented, but it is poorly recorded in the specimen. Unlike the reconstruction of Bossy and Milner (1998: fig. 60A), in the lingual view, the angular is exposed only in a thin strip of bone along the ventral margin of the mandible (mesial lamina sensu Ahlberg and Clack 1998) and does not contribute to the ventral margin of the adductor fossa ( Fig. 3 View Figure 3 ). Instead, the dorsal margin of the angular forms an elongated contact with the prearticular on the lateral surface, a condition which is present in almost all other early tetrapods, with the exception of the highly derived dissorophid Anakamacops Li & Cheng, 1999 ( Liu 2018) . Posterodorsally, the angular is in a short contact with the articular, and posteriorly with the surangular.
Surangular: Apart from the angular, the surangular forms the only other element exposed in lateral view in the posterior area of the mandible ( Fig. 2 View Figure 2 ). Its dorsal portion is markedly overlapped by the squamosal and quadratojugal, and only a thin strip of the bone is exposed laterally as contacting the dorsal margin of the angular in a straight suture. Posteriorly, the surangular constitutes the only element that builds the labial surface of the retroarticular process. Its preserved posterodorsally directed part is quadrangular in shape, but the process itself is incomplete ventrally and would have a broader base, as indicated by the impression on the lingual side ( Fig. 3B View Figure 3 ). The surangular embraces the posterior part of the articular and is exposed as a thin posterior flange located at the lingual side of the retroarticular process (contra Bossy and Milner 1998). In medial view, the surangular is a massively exposed bone contributing to the labial wall of the adductor fossa. Its slightly convex dorsal margin forms a high surangular crest that greatly exceeds the level of the dorsal edge of the prearticular.
Prearticular: The prearticular is a large bone of the posterior region of the mandible, visible only in medial view ( Fig. 3 View Figure 3 ). It might be incomplete anteriorly to some degree, but its original extent cannot be discerned from the state of preservation. The morphology of the prearticular differs markedly from that reconstructed by Bossy and Milner (1998). It is dorsoventrally high and occupies the majority of the posteroventral portion of the mandible, leaving a narrow mesial lamina of the angular exposed ventrally. Dorsally, it contributes to the entire length of the lingual border of the adductor fossa and prevents the angular from participation in it (contra Bossy and Milner 1998). Posterodorsally, the prearticular abuts to the lingual surface of the articular, whereas its ventral border is smoothly convex at the contact point with the angular.
Articular: In medial view, this small quadrangular bone is exposed in the posterodorsal part of the mandible ( Fig. 3 View Figure 3 ). It forms the caudal border of the adductor fossa, and it is surrounded anteriorly, laterally and posteriorly by the surangular. The ventral surface of articular is in contact with the prearticular and a small posterodorsal portion of the angular. The dorsal surface of the bone is poorly preserved, but it bears the articulating surface of the glenoid fossa, composed of a pair of shallow depressions separated by a low ridge for the reception of the double-headed quadrate condyle.
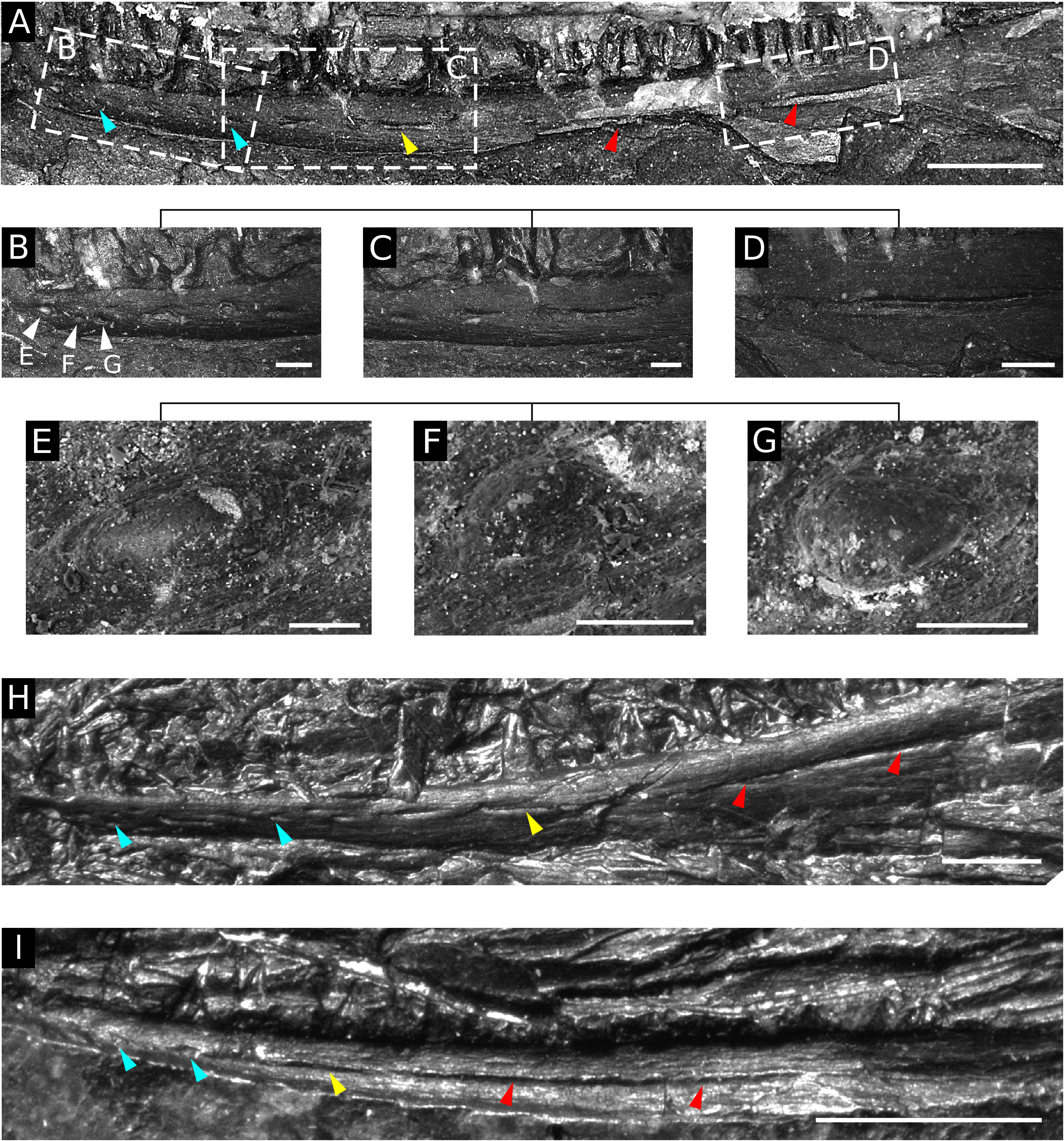
Lateral line system: According to Bossy and Milner (1998), the lateral line system is absent in the skull dermal bones of all urocordylids, including S. scalaris . However, Steen (1938) noticed a row of small pits in the anterior portion of the dentary in ÚGV PAL00158 and interpreted them as the lateral line pits, which were considered by her to be absent in other specimens of this species. A number of small pits occurring in the labial surface of the dentary were also noticed by other authors, who regarded them as the foramina for vessels and nerves ( Stehlík 1924, Bossy and Milner 1998). The imprint of the labial surface of the right dentary indeed possesses in ÚGV PAL00158 a series of small pits anteriorly ( Figs 3B View Figure 3 , 5A–G View Figure 5 ). However, these round–ovate pits (blue arrowheads) become more elongated posteriorly (yellow arrowhead), where they eventually form a previously unrecognized, long, continuous, open sulcus ( Figs 3B View Figure 3 , 5A, D View Figure 5 ; red arrowheads). This open sulcus can be identified reliably as the oral sulcus of the lateral line system, because it is too long to represent a mental foramen. This groove runs externally on the dermal surface at least to the posterior three-quarters of the dentary length, where it disappears abruptly. The identical course of the lateral line sulcus can be observed in several other specimens referred to as S. scalaris ( Fig. 5H, I View Figure 5 ) and represents the first evidence of this structure in Urocordylidae . The identity of the small pits in the anterior portion of the dentary is less certain. Although they could represent mental foramina, as described in some early tetrapods ( Wellstead 1991, Carroll 1998, Anderson and Reisz 2003, Anderson et al. 2020), such openings are never accompanied by an elongated sulcus on the mandible, as seen in S. scalaris . Given that the pits in the anterior portion of the dentary in ÚGV PAL00158 seem to be developed in the same horizontal plane as the continuous lateral line sulcus, rather than irregularly scaưered along the bone, we tentatively consider at least some of them to be a part of the lateral line system. This can also be supported by the fine details in the structure on their surface. In some pits, the inner (ventral) surface possesses a net-like bone structure of the same type as that developed on the labial surface of the dentary ( Fig. 5F View Figure 5 ), whereas in others the same surface appears to be smooth ( Fig. 5E, G View Figure 5 ). The former pattern might indicate the presence of shallow depressions of the lateral line pits, whereas the state in the laưer is ambiguous and might represent either the lateral line pits or mental foramina. It is possible that the lateral line pits in the anterior portion of the mandible are combined with the mental foramina, which would make the proper recognition of these structures in urocordylids even more difficult.
Postcranial skeleton
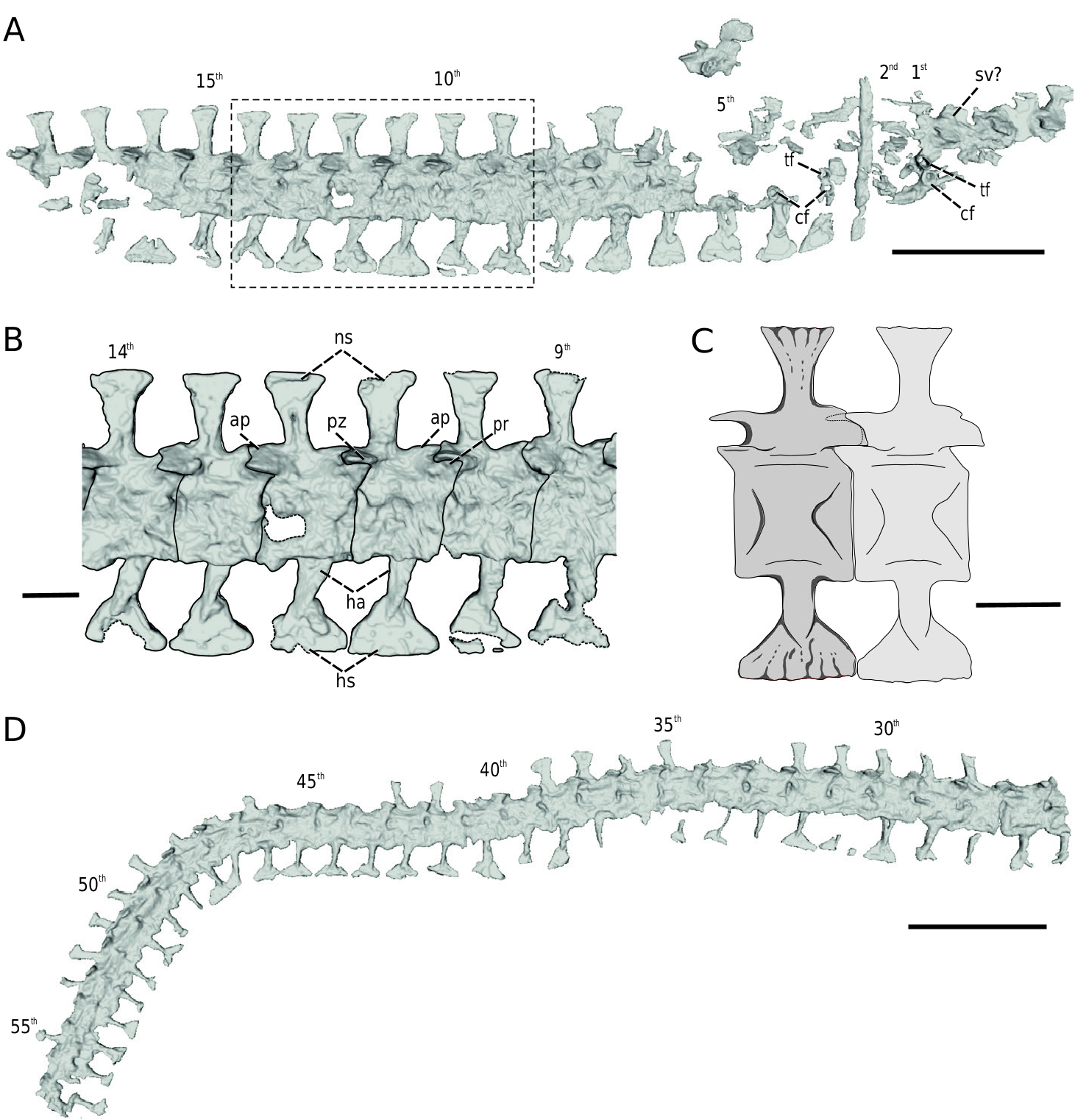
Axial skeleton: The presacral section of the axial skeleton preserves about nine articulated but badly crushed vertebrae, which are anatomically uninformative. Two other isolated presacral vertebrae are displaced approximately to the level of the anterior-most caudals, whereas at least two more very incomplete posterior-most presacrals are articulated to the supposed fragment of the sacral vertebra. The tail section of the axial skeleton is articulated and basically complete ( Figs 1A, C–E View Figure 1 , 6 View Figure 6 ). A total of 70 caudal vertebrae can be observed, but the tail is interrupted in two places, and the posterior-most vertebrae are missing. This means that the total number of caudal vertebrae was higher. The anterior-most caudals are heavily crushed, but still in their original position ( Figs 1C View Figure 1 , 6A, B View Figure 6 ). Likewise, several other more posterior vertebrae are broken, deformed or incomplete to various degrees ( Fig. 6D View Figure 6 ). The external surface of the mid to posterior section of the tail suffered most from pyritic decay, thus those vertebrae are preserved only as indistinct outlines of bituminous residues ( Fig. 1D, E View Figure 1 ; see also Milner 2019). However, their morphology can still be observed through the µCT imaging, because the bone embedded within matrix is essentially intact ( Fig. 6D View Figure 6 ).
Presacral vertebrae: The preserved presacral portion of the column comprises approximately nine vertebrae in articulation ( Fig. 1A View Figure 1 ). These are, however, heavily crushed and do not provide any useful anatomical information. One broken and poorly preserved isolated presacral vertebra possesses an amphicoelous centrum, massive postzygapophyses with posterolaterally directed facets and a suboval zygantrum.
Sacral vertebra: A fragment of the neural arch with a preserved base of the neural spine is identified tentatively as part of the sacral vertebra ( Fig. 6A View Figure 6 ). This identification is based strictly on its anatomical position, and the poor preservation of the vertebra provides no further anatomical details.
Caudal vertebrae: Although the anterior-most caudal vertebrae are heavily crushed, they can be identified reliably on the basis of several distinctive features lacking in the more posterior elements (Bossy and Milner 1998). In agreement with other sauropleurines, the centra of the first two caudals bear well-defined articular facets for caudal ribs and lack any sign of developed haemal arches ( Fig. 6A View Figure 6 ). The articular facets for the capitulum and tuberculum are cylindrical in shape and almost equal in size. They are aligned vertically, with the tubercular facet located immediately above that for capitulum. The third and fourth caudal vertebrae also possess the articular facets for caudal ribs but are already accompanied by haemal arches. As noted by Bossy and Milner (1998), the first haemal spine is distinctive from those of the following caudals in its slightly shorter height and an asymmetrical shape in lateral view, with an oblique ventral border strongly directed anteroventrally.
The more posterior caudal vertebrae have amphicoelous, box-like centra with a slightly constricted ventral surface and lack the articular facets for caudal ribs ( Fig. 6 View Figure 6 ). The haemal arches project from the mid-length of the vertebral centra as two slender, laterally flaưened struts, which meet each other more distally to enclose a roughly suboval intervening haemal canal, as figured in other urocordylids (Thayer 1985, Bossy and Milner 1998, Carlson 1999). The slender, high bases of the haemal arches reach in lateral aspect only about one-third of the length of the corresponding caudal centra, resembling other sauropleurines, but distinguished from the short and broad bases of urocordylines (Thayer 1985, Bossy and Milner 1998). The distal terminations of the merged struts of haemal arches form the anteroposteriorly sharply expanded subtriangular haemal spines. Their distal extensions have about the same length or are slightly shorter than the corresponding centra and closely approach or nearly touch the distal ends of the haemal spines of the adjacent vertebrae.
The neural arches are firmly fused to the centra, as revealed in lateral view, with no clearly defined borders. Some of them are slightly swollen and might indicate the presence of moderately developed lateral ridges extending from the bases of the postzygapophyses to the prezygapophyses ( Fig. 6B, C View Figure 6 ), a genuine feature of Sauropleura (Bossy and Milner 1998) . The subtriangular prezygapophyses of the caudal vertebrae project from the anterior borders of the neural arches and have subhorizontal facets. The anterior accessory apophyses form interlocking projections on the posterodorsal rim of the neural arches. The postzygapophyses are well developed, subhorizontal and bear a prominent ventrolateral rim. The smaller posterior accessory apophyses are exposed in several vertebrae but seem to be less developed than previously depicted (Bossy and Milner 1998).
Fan-shaped neural spines project approximately from the centre of the neural arches. Their proximal bases are noticeably slender and high but broaden more distally to form subtriangular, craniocaudal extensions. However, these extensions are less developed compared with those of haemal spines, well spaced and not contacting each other. The distal margins seem to be moderately crenulated ( Figs 1C View Figure 1 , 6C View Figure 6 ), but this cannot be observed from the µCT. The height of the neural spines is shorter than that of the haemal spines and roughly equals that of the centrum.
Ribs: Apart from a few ribs articulated with heavily crushed presacral vertebrae, three more disarticulated but well-preserved ribs are associated with the specimen; one elongated rib being situated anteriorly close to the caudal margin of the skull, while the two remaining ones lie posteriorly at the level of the anterior-most caudals ( Fig. 7A–F View Figure 7 ). In general, the ribs are most similar to those of stem amniotes ( Smithson 1985, Clack 1987, Holmes 1989, Klembara and Bartík 2000) in having elongated shasss and well-developed proximal articulating heads, with their relative length and form varying according to the position within the vertebral column. An anteriorly displaced rib is identified here as derived from the posterior ‘cervical’ or anterior ‘thoracic’ presacral region ( Fig. 7A, B View Figure 7 ). Its proximal portion has a short, almost triangularly shaped tuberculum that is poorly detached from the well-developed capitular head. It reaches only ~1.5 times the capitulum length and, proximally, bears a suboval articulating facet for connection with the transverse process. The larger capitulum is quadrangular in shape. It bears an anteroventrally elevated rim stretching from its mid-length distally and continuing on the anterodorsal margin of the tuberculum. The shass is long and curved ventrally; it lacks the distal widening, flanges or uncinate processes of many stem tetrapods and temnospondyls ( Moulton 1974, Godfrey 1989a, Clack and Finney 2005). Its proximal portion is suboval in cross-section, whereas more distally it appears to be flaưened.
A long, isolated rib located in the posterior part of the presacral region is similar to the anterior one in most aspects but differs in the morphology and dimensions of its proximal articulating heads ( Fig. 7C, D View Figure 7 ). The tuberculum is comparatively beưer developed, quadrangular in shape and well separated from the capitular head by a deep notch, as present in mid-trunk vertebrae of discosauriscids ( Klembara and Bartík 2000). The capitulum is more elongated; its length is more than twice as long as the tuberculum, and it is slightly constricted at the base. The shass is incomplete distally, yet long and curved ventrally. Here, this rib is interpreted as probably belonging to the mid-trunk vertebra.
A significantly shorter rib associated with an isolated presacral vertebra corresponds best to the posterior trunk region ( Fig. 7E, F View Figure 7 ), as documented by its morphology and position close to the posterior section of the presacral vertebral column. Its proximal articulating heads are clearly separated and almost equally developed, thus close together to articulate with closely spaced capitular and tubercular facets of posterior trunk vertebrae. In accordance with the posterior trunk ribs of other tetrapods, the shass is straight, short and with a pointed distal termination ( Moulton 1974, Klembara and Bartík 2000, Witzmann and Schoch 2018).
Clavicle: The right clavicle is the only appendicular element that can be determined unequivocally in the specimen ( Figs 1F View Figure 1 , 7G, H View Figure 7 ). Most of its dorsal portion, including the ascending process, could be displayed by µCT and is exposed in medial view. The anterior and ventral portions of the clavicular blade are, however, preserved only as imprints of the outer (ventral) surface. The slightly posteriorly tilted ascending process projects from the posterodorsal portion of the clavicle at a relatively acute angle from the ventral blade. It is smooth, probably suboval in cross-section and tapers to the slightly posteriorly curved termination. The ventral blade is broad and smoothly narrows anteriorly, where it forms a slightly jagged edge. Its posteroventral portion bears a patch of imprinted ventral ornamentation consisting of many small, subcircular pits separated by ridges. A short, subtriangular process protrudes from the posteroventral margin of the blade.
Forelimb elements: Stehlík (1924) reported in ÚGV PAL00158 the presence of a poorly impressed right humerus and two radii. These elements could not be recognized clearly, and it is not possible to describe their morphology.



| NMP |
National Museum (Prague) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sauropleura scalaris ( Frič, 1875 )
| Ivanov, Pavel Barták and Martin 2023 |
Sauropleura scalaris (Frič)
| Rocek Z 1988: 525 |
Urocordylus angustatus (Fritsch)
| Rocek Z 1988: 525 |
Sauropleura scalaris (Frič)
| Baird D 1964: 14 |
Urocordylus scalaris Frič
| Steen MC 1938: 206 |
Urocordylus angustatus (Fritsch)
| Steen MC 1938: 213 |
Urocordylus scalaris Frič
| Stehlik A 1924: 242 |
Ptyonius distinctus
| Fritsch A 1901: 90 |
Ptyonius cf. pectinatus Fritsch – Fritsch 1895: 3
| Fritsch A 1895: 3 |
Urocordylus scalaris Frič
| Fritsch A 1881: 129 |
Dolichosoma angustatum Fritsch
| Fritsch A 1880: 117 |
Urocordylus scalaris Frič
| Fric A 1879: 187 |
Urocordylus scalaris Frič
| Fric A 1875: 73 |