
Sphenarium rugosum Bruner, 1906
|
publication ID |
https://doi.org/ 10.5281/zenodo.804182 |
|
publication LSID |
lsid:zoobank.org:pub:27748C60-F64A-4E2C-B5CD-8DB413480DF4 |
|
DOI |
https://doi.org/10.5281/zenodo.6029390 |
|
persistent identifier |
https://treatment.plazi.org/id/A12C6E25-AC21-3B40-2CBE-D603FEA0AFDF |
|
treatment provided by |
Plazi |
|
scientific name |
Sphenarium rugosum Bruner, 1906 |
| status |
|
Sphenarium rugosum Bruner, 1906 View in CoL
(http://lsid.speciesfile.org/urn:lsid: Orthoptera .speciesfile.org:TaxonName:37030)
Sphenarium barretti Bruner, 1906 View in CoL .
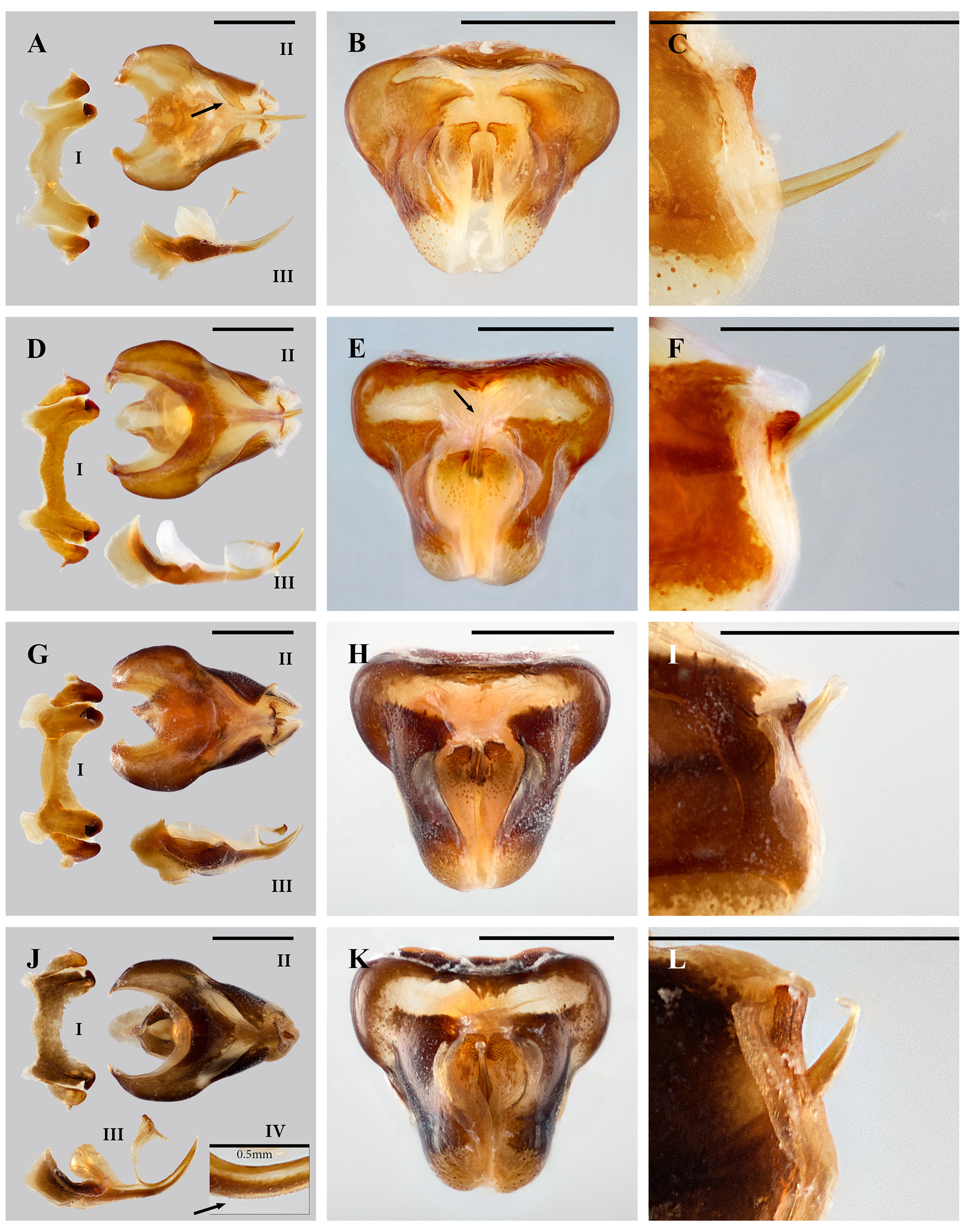
Description. External morphology ( Figs. 14 View FIGURE 14 A, B, C; 9S, T; 15A, B, C, D): total body length ranging from 25.46 to 36.14 mm in females and from 19.8 to 35.07 mm in males; antennae filiform, slightly shorter in females or longer than head and pronotum together in males; head subtriangular-elongated nearly as wide as long in females or moderately longer than wide in males with oval eyes in both sexes; fastigium moderately elongated, nearly half the length of interocular space in both sexes; tegmina spatula-like in both sexes; subgenital plate of males rounded moderately developed posteriorly; dorsal ovipositor valves lanceolate moderately elongated towards the apex. Male genitalia: bridge of epiphallus moderately longer than the length of lateral plates ( Fig. 13 View FIGURE 13 D-I, G-I, J-I). Ectophallus in dorsal view ( Fig. 13 View FIGURE 13 D-II, G-II, J-II) with lateral borders of ramus convergent; basal emargination of cingulum moderately or not developed (morphotypes 1 and 3; Fig. 13 View FIGURE 13 D-II, J-II, respectively) or notably developed (morphotype 2; Fig. 13 View FIGURE 13 G-II); interspace between apodemal plates of cingulum moderately open (morphotypes 1 and 3; Fig. 13 View FIGURE 13 D-II, J-II, respectively) or notably close (morphotype 2; Fig. 13 View FIGURE 13 G-II). Ectophallus in posterior view ( Fig. 13 View FIGURE 13 E, H, K) without a conspicuous sclerotized hollow in the sheath; inflections of supraramus moderately developed with distal margins laterally or dorsally directed; valves of cingulum triangular to slightly quadrangular, relatively small (morphotypes 1 and 2; Fig. 13 View FIGURE 13 E, H, respectively) or large (morphotype 3; Fig. 13 View FIGURE 13 K), slightly developed posteriorly ( Fig. 13 View FIGURE 13 F, I, L). Endophallus in lateral view ( Fig. 13 View FIGURE 13 D-III, G-III, J-III) with elongated pseudoarch, loosely joined to the valves of cingulum; aedeagal valves moderately (morphotype 1; Fig. 13 View FIGURE 13 D-III) or notably (morphotype 2 and 3; Fig. 13 View FIGURE 13 G-III, J-III, respectively) broad, medium-sized (morphotype 1 and 3) or short (morphotype 2), with ventral margins smooth (morphotypes 1 and 2) or somewhat serrated (morphotype 3; Fig. 13 View FIGURE 13 J-IV, arrow), broadly rounded apically ( Fig. 13 View FIGURE 13 E, I, K, arows); aedeagal valves and sclerites together varying from ¾ (morphotype 2; Fig. 13 View FIGURE 13 G-III) to 1 ½ (morphotypes 1 and 3; Fig. 13 View FIGURE 13 D-III, J-III) fold the length of dorsal inflections of endophallic apodemes.
Colouration. Ground colours vary from olive green to brown. Body uniformly coloured ( Fig. 9 View FIGURE 9 T; 15B, D) or with the following colour traits: antennae frequently black or dark brown; fastigium generally reddish; lateral postocular bands frequently present, wide and yellow; dorsomedial line frequently present, narrow and yellowish; dorsal shades often present, covering partially ( Fig. 9 View FIGURE 9 S) or entirely ( Fig. 15 View FIGURE 15 C) the dorsal part of the body, black to brown; lateral shades often present, black or dark brown, mostly evident in the abdomen ( Fig. 9 View FIGURE 9 S); lateral bands of blotches often present with distinct yellow colouration; ventral bands of pronotum often present and yellow; sometimes lateral carinas of pronotum highlighted with yellow ( Fig. 9 View FIGURE 9 S); mesonotum partially or entirely black; lateral blotches of 1st abdominal segment frequently present, whitish or yellowish; hind femora uniformly coloured or with upper medial area black and lower medial area yellowish: knees of hind femora laterally black, dorsally reddish; hind tibia frequently black.
Diagnosis. Externally this species closely resembles S. macrophallicum , S. planum , S. tarascum sp.n., S. crypticum sp.n., and S. infernalis sp.n. Sometimes S. rugosum differs from all these species by its more conspicuous yellow colourations in the dorsomedial line, lateral carinas and lateral bands of blotches, in combination with darker dorsal colourations in the body ( Fig. 15 View FIGURE 15 C). Nevertheless, S. rugosum clearly differs from all these species by the following combination of male genitalia characters: lateral borders of ramus convergent, inflections of supraramus moderately to notably developed, and aedeagal valves moderately or notably broad, medium sized or small, broadly rounded apically.
Distribution. This species is distributed in elevations ranging from 457 to 2344 m a.s.l. and is apparently confined to the eastern Balsas River Basin and the Sierra Madre del Sur in the Mexican states of Guerrero, Mexico, Michoacan, Morelos, Oaxaca, and Puebla ( Fig. 7 View FIGURE 7 A). The morphotype 1 of this species is widely distributed in the western portion of the Balsas River Basin; whereas the morphotype 2 is apparently restricted to inner slope of middle portion of the Sierra Madre del Sur ( Fig. 7 View FIGURE 7 A). The morphotype 3 is only known from three localities east of the middle portion of the Balsas River Basin ( Fig. 7 View FIGURE 7 A).
Material examined. S. rugosum : lectotype m ( Fig. 14 View FIGURE 14 A) and paralectotype f ( Fig. 14 View FIGURE 14 B) from Mexico: Morelos, Cuernavaca, I-4-1899 (C. C. Deam); designation: Rehn and Hebard (1912); location: ANSP . S. barretti : lectotype m ( Fig. 14 View FIGURE 14 C), from Mexico: Guerrero, Rio Cocula , XII (O. W. Barrett); designation: Rehn and Hebard (1912); location: ANSP. We examined only external morphology of this type material. Additional material: 355 m, 201 f, from 80 localities (see Appendix Table 5).
Taxonomic discussion. Bruner (1906) originally described this species based on one male and female syntypes from Cuernavaca, Morelos. He considered the dull granulose and pubescent surface of the type specimen of S. rugosum as the main evidence for separating this species from its congeners, though these traits are variable across the species and the genus. Bolivar (1909) and Hebard (1932) recognized S. rugosum as a valid species. Márquez (1962) synonymized this species within S. purpurascens based on variable and ambiguous morphologic traits of the epiphallus and endophallus of both species. Later, Boyle (1974) and Kevan (1977) re-established S. rugosum as valid species based on more detailed analysis of its male genital morphology. Recent genetic studies (Pedraza-lara et al. 2015; Sanabria-Urbán et al. 2015) have also supported the validity of S. rugosum . In this study we analysed several specimens of S. rugosum , some of them collected at or near the type locality of this species (L414, L435, L440, and L449; Appendix Table 5). Our phylogenetic analysis recovered S. rugosum as a paraphyletic species and we observed relatively low levels of genetic differentiation between S. rugosum and other morphologically different species including adjacent ( S. crypticum sp.n., S. macrophallicum , S. purpurascens , and S. tarascum sp.n.) and distant taxa ( S. minimum , S. variabile , and S. zapotecum sp.n.) (see Table 3). Nevertheless, the unique combination of morphologic traits, and restricted geographic distribution of this species supports its recognition as an independent species within the genus. Moreover, we observed considerable morphological similarity (externaly and in male genitalia), low genetic differentiation (CO1 P-distances <3%; Table 3) and close phylogenetic relationships ( Fig. 11 View FIGURE 11 ) between S. rugosum and the putative new taxa Sphenarium sp. Gro1+Gro8 and Sphenarium sp.n. 6 recognized in Pedraza-Lara et al. (2015) and Sanabria-Urbán et al. (2015) respectively. Therefore, we consider that these taxa represent part of the same species, S. rugosum (morphotype 2 and 3, respectively).
Sphenarium barretti View in CoL was originally described by Bruner (1906) based on an unstated number of specimens, from Rio Cocula, Guerrero. Bruner (1906) originally considered a closer relationship between S. barretti View in CoL and S. purpurascens View in CoL . Posteriorly, S. barretti View in CoL was initially synonymised as S. purpurascens ( Márquez 1962) View in CoL and later as S. rugosum View in CoL ( Boyle 1974; Kevan 1977). In this revision we examined several specimens collected near the type locality of this species (L428, L427 and L423; Appendix Table 5); which were similar in their male genitalia to other S. rugosum View in CoL specimens (morphotype 1). Therefore, we agree in considering S. barretti View in CoL as a synonym of S. rugosum View in CoL .





| ANSP |
Academy of Natural Sciences of Philadelphia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
Sphenarium rugosum Bruner, 1906
| Sanabria-Urbán, Salomón, Song, Hojun, Oyama, Ken, González-Rodríguez, Antonio & Castillo, Raúl Cueva Del 2017 |
S. purpurascens ( Márquez 1962 )
| Marquez 1962 |
Sphenarium barretti
| Bruner 1906 |