
Anomaloglossus apiau, Fouquet, Antoine, Souza, Sergio Marques, Sales Nunes, Pedro M., Kok, Philippe J. R., Curcio, Felipe Franco, Carvalho, Celso Morato De, Grant, Taran & Rodrigues, Miguel Trefaut, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3926.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:BCA3901A-DF07-4FAF-8386-C24649557313 |
|
DOI |
https://doi.org/10.5281/zenodo.5691249 |
|
persistent identifier |
https://treatment.plazi.org/id/03E9294C-FF81-FFAC-FF29-FF21FE3EFEEB |
|
treatment provided by |
Plazi |
|
scientific name |
Anomaloglossus apiau |
| status |
sp. nov. |
Anomaloglossus apiau View in CoL sp. nov.
( Figs. 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 ; Table 1)
Holotype. MZUSP 155833 (SMS962), an adult male collected by Antoine Fouquet, Sergio Marques de Sousa and Pedro Murilo Sales Nunes in October 2011 at ~ 580 m at Serra do Apiaú , Roraima State, Brazil (2.430168 N, 61.411715 W).
Paratopotypes. (n = 35): MZUSP 155814–15, 155818, 155826, 155828–29 155831–32, eight males, and MZUSP 155812–13, 155816–17, 155819–25, 11 females, collected in October 2011 by AF, SMS and PMSN, and MZUSP 155834, 155836, 155838, 155840, 155842–43, 155851–52, eight males, and MZUSP 155837, 155839, 155841, 155844–46, 155850, 155853, eight females, collected in June 2012 by AF and PMSN.
Etymology. The specific epithet is a noun in apposition and refers to the name of the region where the type locality lies (Serra do Apiaú , Roraima State, Brazil).
Adult definition and diagnosis. (1) a small Anomaloglossus (average male SVL = 17.1 mm [15.5–20.0, n = 14], average female SVL = 20.2 mm [18.4–21.8, n = 15]) ( Table 1); (2) body robust; (3) skin on dorsum granular, especially posteriorly, venter smooth; (4) Finger II shorter than I when fingers are appressed, but Finger I shorter than II (3.3 vs. 3.5 mm in holotype) using the method of Kaplan (1997); (5) tip of Finger IV surpassing the base of the distal subarticular tubercle on Finger III when fingers appressed; (6) distal subarticular tubercle on Finger III indistinct; (7) Finger III distinctly swollen in males; (8) fingers with keel-like lateral folds particularly developed preaxially on Finger III; (9) toe webbing basal, with well developed fringes (sensu Grant et al. 2006; keel-like lateral folds sensu Myers & Donnelly 2008); (10) tarsal keel well defined, short, strongly curved, strongly elevated and tubercle-like proximally; (11) black arm gland in males (sensu Grant & Castro-Herrera 1998, see also Grant et al. 2006) and cloacal tubercles absent; (12) pale paracloacal mark present; (13) dorsolateral stripe absent; (15) ventrolateral stripe present in males as a series of more or less conspicuous white dots; (16) oblique lateral stripe present as conspicuous white dots extending from groin midway along flank and inconspicuous to the upper posterior edge of the orbit; (17) sexual dichromatism in throat and venter colour pattern present, with only a few scattered melanophores in females while males possess marbled ventral skin; (18) iris with metallic pigmentation and pupil ring; (20) median lingual process short, slightly longer than wide, tapered, bluntly pointed, smooth (non-papillate), reclined in pit ( Fig. 5 View FIGURE 5 ); (21) maxillary teeth present, small; (22) testes immaculate white, mature ova very large (3 mm in diameter), yellowish brown.
Morphological comparisons with congeneric species. Anomaloglossus apiau can easily be distinguished from A. ayarzaguenai , A. breweri , A. guanayensis , A. megacephalus , A. moffetti , A. murisipanensis , A. parimae , A. parkerae , A. shrevei , A. tamacuarensis , A. tepequem , A. tepuyensis , A. triunfo , A. verbeeksnyderorum , and A. wothuja in having basal toe webbing (moderate to extensive in the aforementioned species).
Anomaloglossus apiau can be readily distinguished from A. beebei , A. kaiei , A. rufulus and A. roraima in having folded fringes on fingers, best developed preaxially on Finger III (fringes barely distinct, not distinctly folded in the aforementioned species).
Anomaloglossus apiau can be distinguished from A. baeobatrachus , A. leopardus , and A. stepheni by its faint, discontinuous oblique ventrolateral stripe (conspicuous and continuous in A. baeobatrachus , A. leopardus , and A.
stepheni ). Anomaloglossus apiau can be distinguished from A. praderioi by lacking a conspicuous light dorsolateral stripe (present in A. praderioi ). Anomaloglossus apiau can be distinguished from Anomaloglossus degranvillei , A. surinamensis and A. apiau by its Finger II longer than Finger I (equal in size in A. degranvillei and A. surinamensis ), and by a third finger swollen in male (not swollen in A. degranvillei and A. surinamensis ). From all the other nominal species of Anomaloglossus it can be distinguished by its long advertisement call composed of pairs of short pulses (see below).
Description of the holotype. An adult male of 17.7 mm SVL. Dorsal skin granular on posterior dorsum, thighs, and shanks; ventral skin smooth ( Fig. 6 View FIGURE 6 ). Cloacal tubercles absent. Head wider than long. Snout bluntly pointed in lateral view, extending past the lower jaw, bluntly pointed in ventral and dorsal views. Nares located close to tip of snout, directed posterolaterally; nares visible in frontal view, barely visible in dorsal or ventral view; IN 39% of HW. Canthus rostralis well defined; loreal region straight, sloping outward to lip. IO 69% of ED. SL 96% of ED, 45% of HL; EN 62% of ED. Tympanic membrane inconspicuous, round, concealed posterodorsally by a diffuse supratympanic swelling; tympanic annulus not visible posterodorsally and partly unpigmented; TYM 35% of eye length. Maxillary teeth present, small. Median lingual process short, about as wide as long, tapered, bluntly pointed, smooth (non-papillate), reclined in lingual pit. Vocal sac and vocal slits visible.
Hand moderate in size, 27% of SVL, 79% of HW. Relative length of fingers III> IV> I> II. Fingers unwebbed. Discs of fingers expanded, disc on Finger III and IV widest (state 2 sensu Grant et al. 2006), discs on other fingers sub-equal (state 1 sensu Grant et al. 2006). Fingers with fringes, best developed preaxially on Finger II and III ( Fig. 6 View FIGURE 6 ). Palmar tubercle large, rounded, thenar tubercle smaller, elliptical; one or two round to ovoid subarticular tubercles (one each on Fingers I and II, two each on Fingers III and IV). Tip of Finger IV distinctly surpassing the base of distal subarticular tubercle on Finger III when fingers appressed. No fleshy supracarpal fold atop wrist ( Fig. 6 View FIGURE 6 b). Finger III strongly swollen dorsally and preaxially, swelling extending from the wrist to the third subarticular tubercle [state 3 sensu Grant et al. (2006)].
Hind limbs robust, moderately long, TIL 49% of SVL. Relative lengths of appressed toes IV> III> V> II> I; Toe I short, not reaching the base of subarticular tubercle of Toe II. Toe discs expanded, distinctly larger than finger discs, largest on Toes II, III and IV. Feet weakly webbed; all toes with well-developed fringes. Webbing formula I 1 ½–2- II 2 +–3- III 2 ½–4 IV 4 -– 2-V ( Fig. 6 View FIGURE 6 b). Inner metatarsal tubercle small, elliptical; outer metatarsal tubercle small, round, about half the size of the inner metatarsal tubercle. One to three round to ovoid subarticular tubercles (one each on Toes I and II, two each on Toes III and V, and three on Toe IV with proximal tubercle on Toe IV the smallest and least conspicuous). A strong outer metatarsal fold is present, coextensive with the fringe on Toe V and almost reaching the outer metatarsal tubercle. Tarsal keel weakly curved proximally, discontinuous with the fringe along the outer edge of the first toe, strongly elevated and tubercle-like proximally ( Fig. 6 View FIGURE 6 b).
Colour of holotype in life. Dorsal ground colour brown with a light brown interorbital triangle ( Fig. 6 View FIGURE 6 c). Upper surface of arm with a bright yellow mark with ill defined dark brown and white marks on forearm and wrist; upper surface of thigh yellow with well defined dark brown transverse bands, shank and foot light brown also with well defined dark brown transverse bands. Flanks dark brown, with some blue/whitish irregular iridescent blotches on the lower part, and a lateral stripe of yellowish spots from the inguinal region to the middle of the flank becoming conspicuous again with a short orange stripe on the upper posterior corner of the eye. Dorsolateral stripe absent.
Upper lip brown with iridescent white marks below the eye; loreal region and side of head dark; a small whitish stripe is present from arm insertion to tympanum partly depigmented. A dark brown stripe is present on the anterior edge of upper arm, tapering from arm insertion to forearm. Throat, belly and undersurface of arm, thigh and shank marbled with patches of dark brown melanophores. Rear of thigh and cloacal region brown. Paracloacal marks orange. Palms and soles brown ( Fig. 6 View FIGURE 6 c).
Colour of holotype in preservative. Dorsal ground colour dark brown with a thin light gray to light brown interorbital stripe and a similarly coloured small spot behind the eyes ( Fig. 6 View FIGURE 6 a). Dorsal surface of arm brown with dark brown transverse bands on forearm and wrist, upper arm cream; dorsal surface of leg brown with well-defined dark brown transverse bands on thigh, shank, and foot. Flanks dark brown with some irregular spots and a few whitish irregular blotches ventrolaterally. A few white spots (covering small tubercles) form a dotted oblique lateral stripe that only extends from the inguinal region to the middle of the flanks. Throat, belly, and under-surface of arms and legs marbled with patches of melanophores; rear of thigh and cloacal region brown. Paracloacal marks whitish. Palms and soles dark brown ( Fig. 6 View FIGURE 6 b).
Variation. Males are smaller than females (average male SVL = 17.1 mm 15.5–20.0 mm, n = 14; average female SVL = 20.2 mm, 17.7–21.8 mm, n = 15), usually with third Finger III distinctly swollen. There is marked sexual dichromatism ( Fig. 7 View FIGURE 7 ). Dorsal colouration of females is light to dark brown with small diffuse black marks and no lateral dotted stripe. Males, however, are generally darker with a highly variable and conspicuous pattern as well as a lateral stripe consisting of white, bluish and even sometimes orange spots. Whitish-blue marks also occur ventrally on flank, shoulder and lip in males, but found in one female only. Hind limbs are also generally markedly sexually dichromatic, being of similar colouration to the dorsum in females, but with orange markings of various extents in males (from inexistent to small patches, or even sometimes becoming the main background colour). Ventral colouration of females is immaculate white, sometimes with diffuse melanophores and a yellowish throat. Males, however, generally have extensively pigmented ventral surfaces (with one exception; Fig. 7 View FIGURE 7 , MZUSP 155836, SVL 16.8 mm, despite conspicuous dorsal colouration and swollen third finger) giving a marbled aspect, especially on throat.
In addition to sexual dimorphism in size and colour pattern, variation is extensive within sexes. Among females, dorsal colouration varies from light to dark brown and from immaculate to vermiculate with dark patches. The dark bands on thigh and shank vary from conspicuous to very faint. Lip pattern varies from almost immaculate cream to covered with dark brown marks. Shoulder is orange to light brown. In males, variation in colouration is even more striking and might depend partly on reproductive activity given that more conspicuously coloured males also have swollen third fingers. Dorsal colouration in males varies from immaculate dark brown to light brown with various different patterns of dark brown markings. The distribution of bluish white iridescent dots ranges from a few large ones on the ventral part of the flanks and lips to completely peppering flanks and lips. Limbs and ventral surface are also highly variable in colouration (see above).
Advertisement calls. The call is a long trill of 19.6 s on average (sd = 11.0, range = 7.0–39.4, n = 10) consisting of pairs of very short pulses (average duration of pulses = 0.03 s, sd = 0.005, 0.02–0.04, n = 50; interpulse interval within pair 0.03 s and between pairs 0.073 s on average) (Fig. 8). Pulse rate per second 11.3 on average (sd = 0.55). Four harmonics are visible, with the dominant frequency located in the lowest harmonic (= fundamental frequency). The dominant frequency increases during the first seconds of the call (~4 s) but reaches a plateau of 4.3 kHz on average (sd = 0.129, 4.147–4.526, n = 5; Fig. 8; Table 2 View TABLE 2 ).
FIGURE 8. Advertisement call of Anomalogossus apiau ( IND 5) showing call length (>17 s) and spectral structure. Note the paired pulses (inset). The specimen was recorded in October 2011 around 11 am at the type locality (21°C).
The temporal structure of the call differs markedly from that of congeneric species in the duration of the call. Anomaloglossus degranvillei ( Marty & Gaucher 1999) , A. surinamensis ( Ouboter & Jairam 2012) and A. praderioi (Kok 2010) emit a single note at rates between 50 and 100 calls/min; A. beebei ( Kok et al. 2006a), A. tamacuarensis ( Myers & Donnelly 1997) and A. kaiei ( Kok et al. 2006b) emit short series of pulses (4–5, 2, and 1–2 pulses/call, respectively); A. baeobatrachus ( Marty & Gaucher 1999) and A. stepheni ( Lima et al. 2006; Martins 1989) emit short trills of 7–19 pulses and less than 1 s of duration. Anomaloglossus rufulus emits shorter (<3 s) and regular trills with lower pulse rate (7 pulses/s) ( Barrio-Amorós & Santos 2011). The only known Anomaloglossus call that has a similar temporal structure to the call of A. apiau is the one of A. verbeeksnyderorum ( Barrio-Amorós et al. 2010) . However pulse rate of A. apiau is higher (11.27 pulses/s vs. 8.87 in A. verbeeksnyderorum ) and the pulses are paired (vs. regularly spaced in A. verbeeksnyderorum ).
MEAN SD IND 1 REC1 IND 2 REC1 IND 3 REC1
Distribution and ecology. Anomaloglossus apiau sp. nov. was observed exclusively along rocky streams (only a few meters away from water maximal level), on the slopes of Serra de Apiaú above 500 m elevation and up to the summit at ~ 1400 m. These streams are usually shallow, but they can be fast flowing and up to 15 m wide (but unlikely torrential) or reduced to rivulets with only a film of water running on the rocks ( Fig. 6 View FIGURE 6 d). Males call from the stream banks or on rocks; they only call from the banks alongside larger streams. The species is locally abundant with one calling male found every ~ 5 m along the stream when calling activity is at its peak, i.e. during late afternoon or after rainfall.
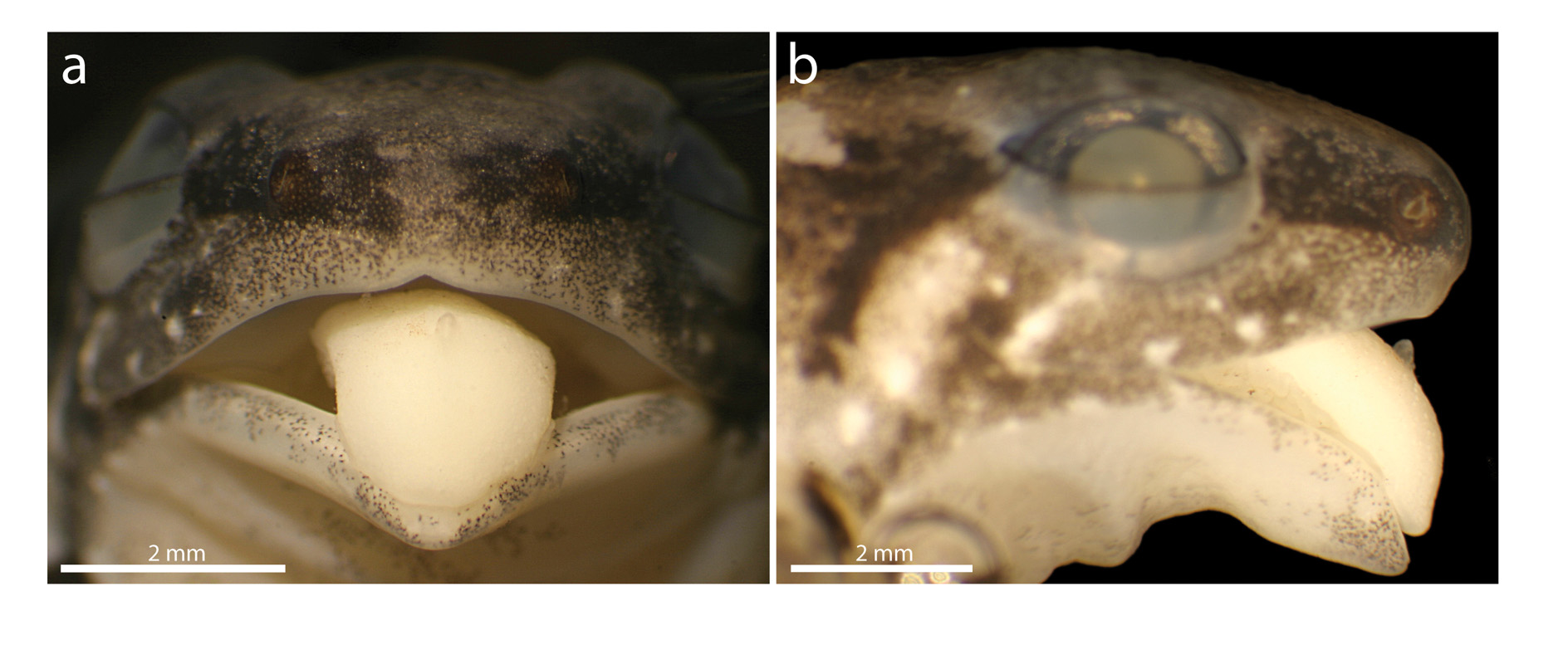


TABLE 2. Acoustic parametres of six recordings of five uncollected males of Anomaloglossus apiau sp. nov. recorded in October 2011 from the same location as the holotype, except IND 5 recorded at the top of the Apiaú mountain.
| Recording duration (s), temperature (°C) and elevation (m) Call duration (s) 19.56 | 97s (23°C) 11.05 9.08 | 580m asl n=4 | 20s 18.4 | 580m asl n=1 | 87s 39.4 | 580m asl n=1 |
|---|---|---|---|---|---|---|
| Inter-call interval (s) 11.17 | 7.24 13.7 | n=3 | NA | NA | ||
| Pulse duration (s) 0.033 | 0.005 0.03 | n=10 | 0.03 | n=10 | 0.03 | n=10 |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.