
Anomaloglossus dewynteri, Antoine & Vacher & Courtois & Villette & Reizine & Gaucher & Jairam & Ouboter & Kok, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4379.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:31A398C2-9973-4666-B201-F37B98608CA4 |
|
DOI |
https://doi.org/10.5281/zenodo.5987735 |
|
persistent identifier |
https://treatment.plazi.org/id/083B6A4A-FF9D-2700-19B7-5EA3FAEFFB51 |
|
treatment provided by |
Plazi |
|
scientific name |
Anomaloglossus dewynteri |
| status |
sp. nov. |
Anomaloglossus dewynteri sp. nov.
Anomaloglossus sp. “ Itoupé” Vacher et al. 2017
Holotype. MNHN2017.0111 View Materials (field no. AF3644/APA-973-23-2), an adult male, collected by Antoine Fouquet, 11 January 2016, near base camp on Mont Itoupé , French Guiana, 3.0230 N 53.0955 W, elevation ca. ~ 600 m ( Figure 7 View FIGURE 7 ). GoogleMaps
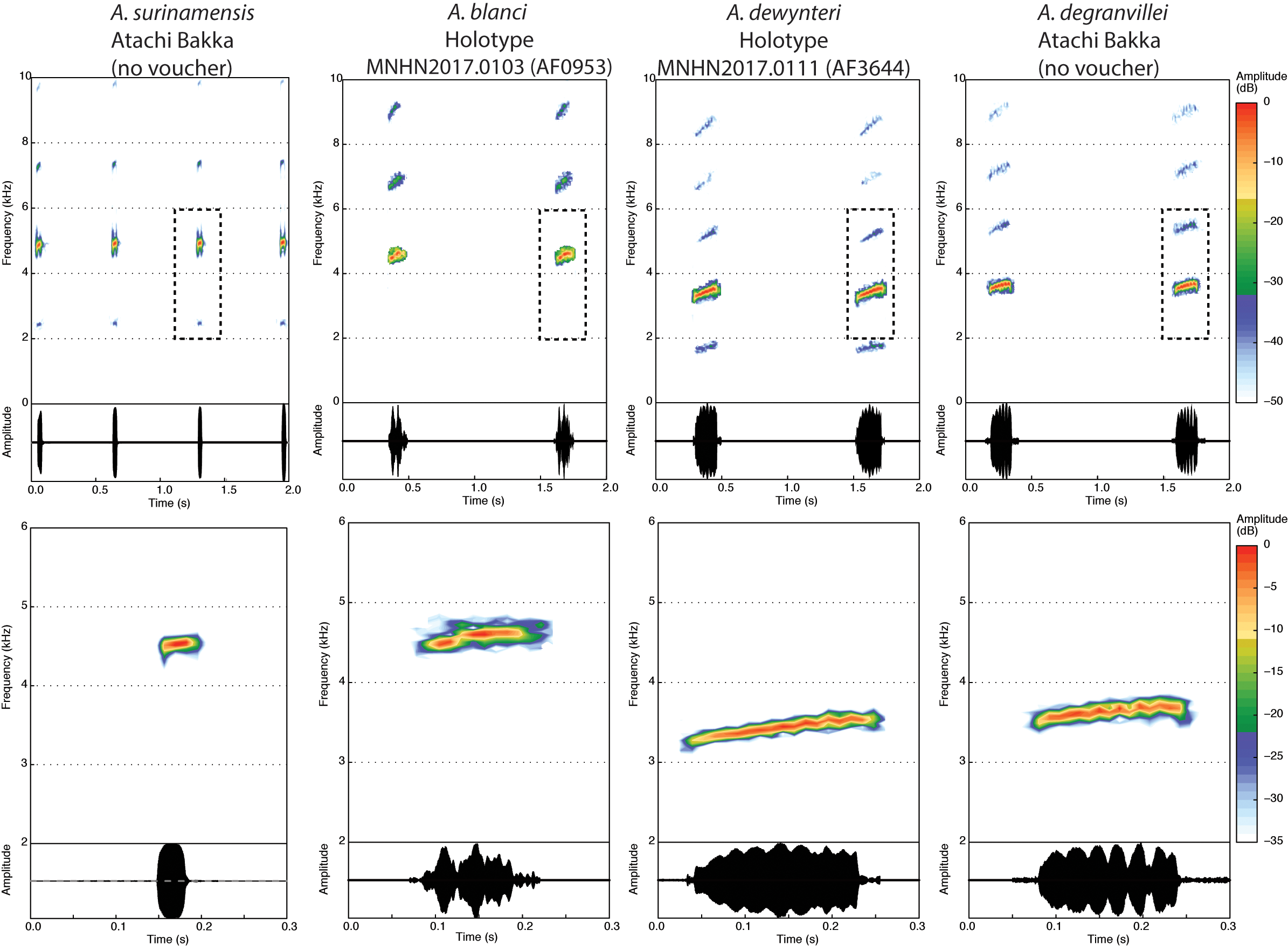
Paratopotypes. Three specimens: MNHN2017.0112 View Materials (field no. AF0574; not measured because badly preserved), an adult male, collected by Maël Dewynter, 0 5 April 2010, near base camp on Mont Itoupé , French Guiana, 3.0230 N 53.0955 W, ~ 600 m elevation GoogleMaps . MNHN2017.0113–4 (field no. AF3686/APA-973-23-1, AF3645/ APA-973-23-3), two adult males, collected by Antoine Fouquet, with the holotype.
Etymology. This species is dedicated to our friend Maël Dewynter for his invaluable contribution to the herpetology of French Guiana.
Adult definition and diagnosis. We assigned the new species to the genus Anomaloglossus based on previous studies (Fouquet et al. 2012; Vacher et al. 2017) and the presence of a median lingual process. The new species belongs to the A. degranvillei clade according to Vacher et al. (2017).
(1) Medium-sized Anomaloglossus (average male SVL= 19.9 mm [19.4–20.4, n=3], female unknown) ( Table 1); (2) body robust; (3) skin tuberculate on dorsum (particularly the posterior half) and legs, with a larger tubercle on each eyelid, ventral skin smooth; (4) conspicuous dark brown to black glandulous supratympanic fold flanked anteroventrally by a discontinuous stripe (orange in life, cream in preservative) running from the ventroposterior edge of the eye and extending onto the upper arm; (5) tympanum indistinct; (6) snout short and acute in lateral view; (7) nares oriented ventrolaterally, situated near tip of snout; (8) Finger II shorter than Finger I when fingers adpressed; (9) tip of Finger IV not reaching distal subarticular tubercle on Finger III when fingers adpressed; (10) distal subarticular tubercle on Finger III and IV indistinct; (11) Finger III not distinctly swollen in males; (12) fingers with fringes particularly developed on preaxial edges of Fingers II and III; (13) toes moderately webbed, with well-developed fringes (sensu Grant et al. 2006; keel-like lateral folds sensu Myers & Donnelly 2008); (14) tarsal keel well-defined, curved, slightly tuberclelike; (15) no black arm gland in males (sensu Grant & Castro 1998. see also Grant et al. 2006); (16) cloacal tubercles present; (17) paracloacal mark inconspicuous; (18) dorsolateral stripe absent or inconspicuous, dorsal part of flanks darker than dorsum; (19) ventrolateral stripe sometimes present as a broken, poorly defined pale brown blotch, ventral part of flanks with flecks white or bluish (in life); (20) sexual dichromatism in throat colour pattern unknown (although likely because present in other species of the clade), throat solid black in reproductive males; (21) abdomen and ventral side of legs dark grey with small ill-defined white dots; (22) iris with metallic pigmentation and pupil ring interrupted ventrally by transversal black pigmentation; (23) median lingual process longer than wide, tapered, bluntly pointed, smooth (nonpapillate), reclined in pit; (24) single note call of 0.221– 0.237 s length and dominant frequency at 3.45–3.59 kHz (n=3) ( Table 2; Figure 3 View FIGURE 3 ).
Morphological comparisons with other lowland Anomaloglossus . The only other species group cooccurring with the Anomaloglossus degranvillei species group is the A. stepheni species group, represented by A. baeobatrachus in French Guiana, which is readily distinguishable by conspicuous dorsolateral stripes, swollen third fingers in males and absence of webbing and fringes on toes.
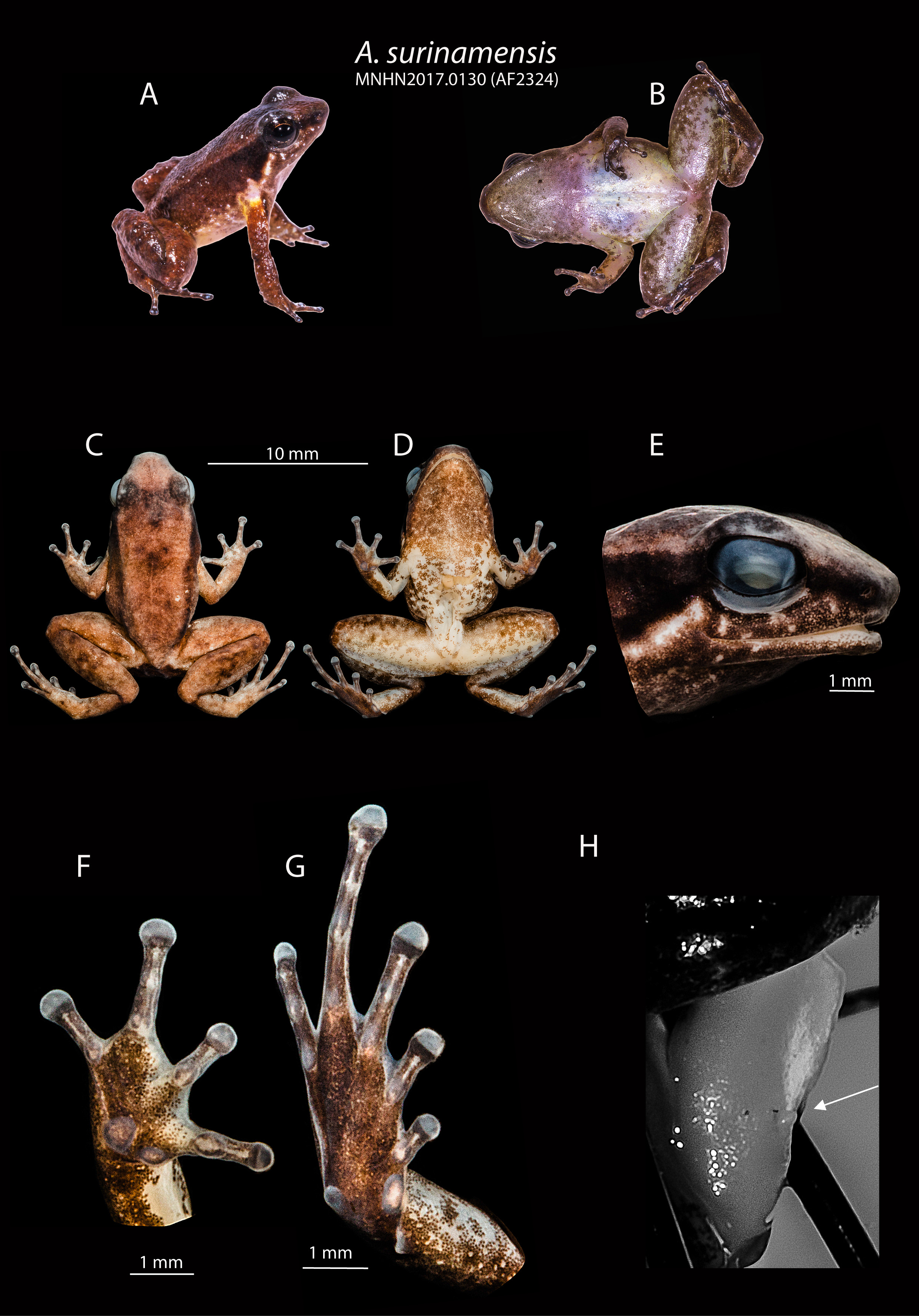
Within the Anomaloglossus degranvillei group, A. dewynteri can be distinguished from A. surinamensis ( Figure 4 View FIGURE 4 ) by (1) a shorter snout, acute in lateral view (vs. rounded in A. surinamensis ); (2) a larger body size in males (X¯ = 19.9 mm, range 19.4–20.4 mm in A. dewynteri [n=3] vs. X¯ =14.75, range 14.0– 15.3 mm in A. surinamensis [n=8]); (3) more developed fringes on fingers and toes; (4) presence of a large tubercle on the top of the eyelid (inconspicuous in A. surinamensis ); (5) belly dark grey with small white spots (vs. cream with irregular grey blotches less abundant posteriorly in A. surinamensis ); (6) ventral surface of thigh dark grey (vs. cream with dark spots on the edges in A. surinamensis ); (7) larger MLP (> 0.5 mm vs <0.5 mm in A. surinamensis ; (8) call characterized by much longer pulsed notes (pulsed note X¯ =0.228, range 0.221– 0.237 s [n=3] in A. dewynteri vs. tonal note X¯ =0.032, range 0.028– 0.037 s in A. surinamensis [n=20]) emitted between longer intervals (X¯ =0.76, range 0.60– 0.88 s in A. dewynteri [n=3] vs. X¯ =0.573, range 0.372– 0.825 s in A. surinamensis [n=20]) and with lower dominant frequency (X¯ =3.52, range 3.45–3.59 kHz in A. dewynteri [n=4] vs. X¯ =4.89 kHZ, range 4.55–5.35 kHz in A. surinamensis [n=20]).
Anomaloglossus dewynteri can be distinguished from A. blanci ( Figure 2 View FIGURE 2 ) by (1) its larger size in males (X= 19.9 mm, range 19.4–20.4 mm in A. dewynteri [n=3] vs. X¯ =16.9, range 15.9–18.8 mm in A. blanci [n=8]); (2) belly from solid black to dark grey with abundant small conspicuous white spots (pale grey with small white illdefined spots in A. blanci ); (3) ventral surface of thighs dark grey (yellowish in life and cream in preservative in A. blanci ); (4) strong metacarpal ridge (inconspicuous in A. blanci , Figures 2F View FIGURE 2 , 7F View FIGURE 7 ); (5) call with longer notes (X=0.228, range 0.221– 0.237 s [n=3] in A. dewynteri vs. X¯ =0.094, range 0.090– 0.103 s in A. blanci [n=6]) and higher dominant frequency (X¯ =3.52, range 3.45–3.59 kHz in A. dewynteri [n=4] vs. X¯ =4.75, range 4.48–5.41 kHz in A. blanci [n=6]).
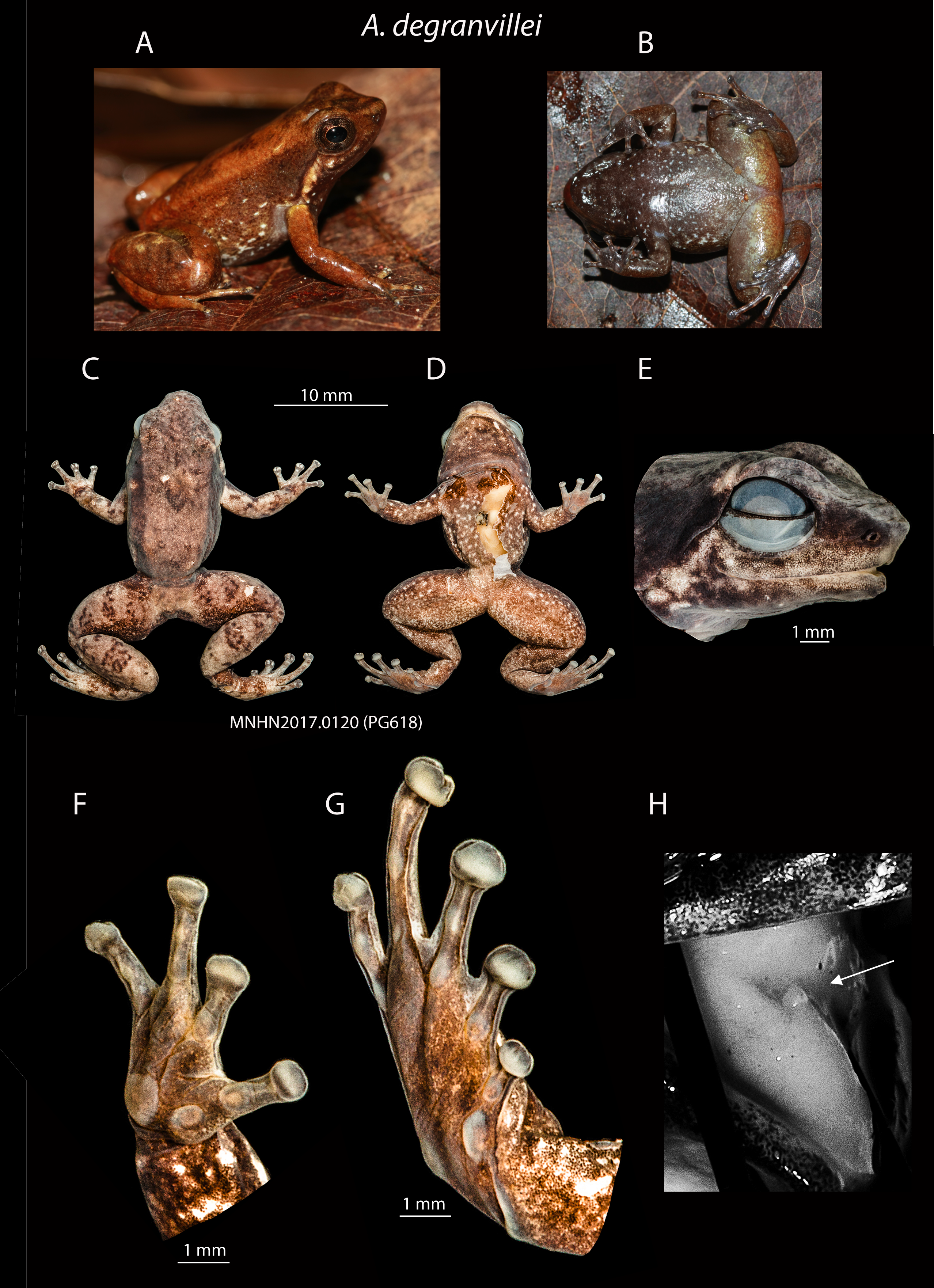
Anomaloglossus dewynteri can only be distinguished from the most similar A. degranvillei ( Figure 5 View FIGURE5 ) by its call consisting of longer notes (X¯ =0.228, range 0.221– 0.237 s [n=3] in A. dewynteri [n=4] vs. X¯ =0.158, range 0.157– 0.160 s in A. degranvillei [n=2]) ( Figure 3 View FIGURE 3 ; Table 2). Molecular data provided by Vacher et al. (2017) demonstrated that these two species harbour well-differentiated mtDNA lineages (2.3% on 16S) and our phylogenetic analysis indicates that A. dewynteri is not the sister species of A. degranvillei (see below). Anomaloglossus dewynteri could possibly be distinguished from A. degranvillei by its more tuberculate skin and by its darker dorsal and lateral colouration. However, these two features should be taken with caution given the small number of specimens available and the paucity of available data from living specimens.
Description of the holotype ( Figure 7 View FIGURE 7 ). An adult male 19.9 mm SVL; body robust; head wider than long, HL 85% of HW; HL 32% of SVL; dorsal skin tuberculate, large tubercle on eyelids, snout large, rounded to nearly truncate in dorsal view, acute in lateral view, extending past lower jaw, SL 56% of HL. Nares located anterolaterally; canthus rostralis rounded but well defined, loreal region concave; IN 40% of HW; EN 31% of HL, 77% of ED. Tympanum indistinct; supratympanic fold present, extending from ventrodorsal corner of the eye onto the upper arm; choanae round, small, located anterolaterally.
Forelimb slender, skin tuberculate; metacarpal ridge present; HAND 25% of SVL; Finger I longer than Finger II when fingers adpressed; fingers large and flattened without webbing, lateral fringes present on preaxial edges of Fingers II and III; Finger III not distinctly swollen; tip of Finger IV not reaching distal subarticular tubercle on Finger III when fingers appressed; finger discs expanded, wider than long, about 1.5X width of digit; width of disc on Finger III 0.8 mm. Relative lengths of adpressed fingers III> IV> I> II; palmar tubercle large, quadrangular. 1.0 mm in diameter (larger than Finger III disc), thenar tubercle small (equal to Finger III disc), elliptic, half the size of palmar tubercle, well separated from palmar tubercle. Only basal subarticular tubercles of Fingers III and IV are conspicuous; Finger I subarticular tubercle largest followed by Finger II subarticular tubercle, basal subarticular tubercle on Finger III and IV smaller, subequal. The holotype has a missing left hand (all the individuals collected in 2016 had some atrophied or missing fingers/toes).
Hind limb robust, skin tuberculate; TL 47% of SVL; heels not in contact when hind limbs are flexed at right angle to sagittal plane of body; FL 42% of SVL; relative length of adpressed toes IV> III> V> II> I; Toe I very short, its tip reaching the base of subarticular tubercle on Toe II when toes adpressed; discs on toes larger than width of toes; disc on Toe I only slightly larger than width of digit. Size of disc on Toe IV 1.0 mm. Feet moderately webbed, webbing present between Toes I–IV, webbing without melanophores; lateral fringes present on all toes. Toe webbing formula I 0- -1- II 1 ½-2- III 1 - -3 IV 3 - -2+ V. One to three subarticular tubercles on toes as follows: one on Toes I and II, two on Toes III and V, three on Toe IV. Inner metatarsal tubercle protuberant elliptical, 0.7 mm in length, outer metatarsal tubercle round, protuberant, 0.5 mm in diameter. Tarsal keel well defined, tubercle-like and strongly curved at proximal end, extending distally to the fringe on preaxial edge of Toe I. Metatarsal fold strong.
Colour of holotype in life ( Figure 7 View FIGURE 7 ). Dorsal colour dark brown, faint dorsolateral stripe, upper flanks black becoming slightly lighter ventrally with a few white-blue freckles. Supratympanic fold black bordered by a discontinuous orange postocular stripe. Upper lip brown, loreal and internarial region black. Throat solid black becoming slightly paler posteriorly, belly and ventral parts of legs dark grey with light blue freckles. Iris with reddish metallic pigmentation and pupil ring interrupted ventrally by transversal pigmentation.
Upper and posterior surfaces of arm pale orange, black anteriorly (same colour as throat). Dorsal surfaces of thigh, shank and tarsus with diffuse combination of orange, pale brown and two wide poorly defined dark brown cross bands; more cross bands on tarsus. Paracloacal marks inconspicuous. Toes and digits with small light blue dots. Palms and soles black.
Colour of holotype in preservative. After one year in preservative (70% ethanol), the specimen faded and he dorsal colouration now varies from brown to grey. The bluish freckles turned cream as well as the orange and reddish marks.
Variation among type specimens. Measurements (range, mean, and standard deviation) of the type series are provided in Table 1. There is only slight variation among specimens of the type series. However, specimens photographed in 2010 displayed a dorsal colouration going from brown with black blotches to uniform brick; flank colouration black with various pattern of brown ventroposterior marks as well as white and bluish dots ( Figure 6 View FIGURE 6 ).
Advertisement call ( Figure 3 View FIGURE 3 ). Three specimens (one uncollected) calling from the leaf litter near a stream were recorded from a distance of about 1 m and at temperatures ranging from 21 to 24°C. They emitted single pulsed notes (note length X¯ =0.228, range 0.221– 0.237 s [n=3]) at a regular pace (inter-note interval X¯ =0.761, range 0.600– 0.883 s). The spectral structure of the note has a developed harmonic structure and the dominant frequency is 3.52 kHz on average (range 3.45–3.59 kHz) with a slight upward modulation (ca. 0.2 kHz).
Distribution and ecology. Anomaloglossus dewynteri is only known from two rocky streams close to each other at 600 m elevation, on the slope of the Itoupé massif, an isolated mountain reaching 800 m elevation in the south-eastern part of French Guiana. Males call from the banks of these streams during the day and their activity seems fostered by rainfall. Many individuals were observed during the first exploration of the massif in 2010. However, only four individuals were detected in 2016 despite eight days of searching by four herpetologists, and despite any obvious perturbation of the habitat. This species co-occurs with A. surinamensis on the massif, but the two species were not found together along the same streams.
Anomaloglossus dewynteri has the same reproductive mode as A. blanci and A. degranvillei , with endotrophic tadpoles carried by the male until metamorphosis. In 2010, one individual carrying six metamorphs at similar stages (Gosner stages 42–43) was observed. We cannot exclude that these metamorphs are not from two different clutches of smaller size.
Comparison with the type series of Anomaloglossus degranvillei . Our examination of the original type series used by Lescure (1975) to describe Anomaloglossus degranvillei shows that it included three distinct species of the A. degranvillei clade occurring in French Guiana. Based on the original description and according to the characters observed in the holotype, we restrict the type series to the holotype and three of the original paratypes (MNHNP1973- 1656–58) from Atachi Bakka (3.5455 N 53.9068 W).
Paratypes MNHNP 1659–63 and MNHNP 1667–70 from Eaux Clément (4.6556 N 52.2480 W) and MNHNP 1665-66 from Cacao (4.5610 N 52.4629 W) correspond to A. blanci based on the following combination of characters: (1) body size comprised between 15.9–18.8 mm in males; (2) belly pale grey with small white freckles; and (3) protruding snout in lateral view.
Paratype MNHNP1973-1672 from Galbao (3.600 N 53.2833 W), MNHNP1664 from Saül (3.62556 N 53.2072 W) and MNHNP1676 from Crique Grégoire (5.0975 N 53.0506 W) correspond to A. surinamensis as inferred from the following combination of characters: (1) small body size (between 14.0– 15.3 mm in males 16.0– 17.1 mm in females); (2) snout short and rounded in lateral view; (3) toes moderately webbed, with fringes (4) abdomen and ventral side of legs cream with brown freckles.
Unfortunately, we could not examine LACM44211-12 from Crique Ipoussing (4.1307 N 52.5760 W), LG870–873 from Eaux Clément (4.6556 N 52.2480 W) and LACM44213–23 from Montagne Tortue (4.300 N 52.3667 W), but these localities lie within the range of Anomaloglossus blanci and, to our knowledge, no other species of the group occur there. These paratypes are thus excluded from the type series of A. degranvillei .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |