
Allobates nunciatus, Moraes & Pavan & Lima, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4648.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:E60E50F9-CE24-4CB6-BC7C-2668854564CA |
|
persistent identifier |
https://treatment.plazi.org/id/7ED72E73-39BF-41A5-8946-7F7C101C0C69 |
|
taxon LSID |
lsid:zoobank.org:act:7ED72E73-39BF-41A5-8946-7F7C101C0C69 |
|
treatment provided by |
Plazi |
|
scientific name |
Allobates nunciatus |
| status |
sp. nov. |
Allobates nunciatus sp. nov.
( Figs. 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
urn:lsid:zoobank.org:act:7ED72E73-39BF-41A5-8946-7F7C101C0C69
Allobates cf. crombiei Moraes et al. 2016 View in CoL
Holotype. Adult male ( INPA-H 40486 , field number DT 3955, 21.4 mm SVL, Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ) collected by D. Pavan on 15 December 2012 on the east bank of Middle Tapajós River , municipality of Itaituba (05°09’07.1” S 56°27’35.2” W, 96 m asl), state of Pará, Brazil. GoogleMaps
Paratypes. Ten specimens, three adult males ( INPA-H 40305 , 40307 , 40482 ), six adult females ( INPA-H 40324 , 40475 , 40476 , 40480 , 40484 , 40489 ), and one young female ( INPA-H 40485 ) collected by D. Pavan, L.J.C.L. Moraes, T.F.D. Rodrigues, J. Gomes and L.F. Storti between July 2012 and October 2013 at east bank of Middle Tapajós River , municipalities of Itaituba and Trairão, Pará state, Brazil ( Fig. 5 View FIGURE 5 ). Of these, eight were collected in the west bank of Jamanxim River , inside the municipality of Itaituba ( INPA-H 40305 , 40307 , 40475 , 40480 , 40485 , 40489 at 5°13’49.1” S 56°55’32.7” W, 82 m asl GoogleMaps ; and INPA-H 40324 , 40476 at 4°44’47.6” S 56°36’57.3” W, 92 m asl) GoogleMaps and two in the east bank of this river, inside the municipality of Trairão ( INPA-H 40482 , 40484 at 5°06’12.6” S 56°26’35.5” W, 91 m asl) GoogleMaps . Another eight adult males and one female were collected by A.P. Lima in 16 January 2014 and 18 February 2018 in three additional localities at banks of Jamanxim River : inside the municipality of Itaituba ( INPA-H 40320 , 40474 , 40477 , 40478 , 40481 , 40483 at 06°16’ S 55°39’ W, 218 m asl GoogleMaps ; and INPA-H 40308 , 40479 at 06°17’42.7” S 55°49’19.6” W, 221 m asl) GoogleMaps and Trairão ( INPA 40488 View Materials at 04°45’25.1” S 55°57’56.4” W, 167 m asl) GoogleMaps . We designate the adult female specimen INPA-H 40489 ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ) as the species allotype.
Etymology. The specific epithet ‘ nunciatus ’ (Latin verb: announce, report; derived from ‘nuntiatus’), refers to the great conspicuity of the loud advertisement call of the species, allowing it to be readily recognized by the human ear wherever it occurs.
Generic placement. The new species was allocated to the genus Allobates based on its external morphology similar to other species of the genus (see comparisons), behavior (diurnal activity), and molecular phylogenetic relationships.
Definition. A large species of cryptically-colored Allobates , which can be characterized by the following combination of characters: (1) mean SVL of adult males 20.8 mm (range 19.2–21.7 mm), mean SVL of adult females 20.9 mm (range 19.3–22.0 mm) ( Table 1 View TABLE 1 ); (2) skin texture of dorsum mostly smooth, with some granules posteriorly; (3) finger III not swollen in adult males; (4) absence of lateral fringes and basal webbing on fingers; (5) paired dorsal digital scutes present; (6) fingers and toes discs expanded, larger in toes II, III, and IV; (7) basal webbing between toes II–III and III–IV; (8) finger II longer than or with a similar size as finger IV; (9) toe III length reaches or surpasses the middle of the central sub-articular tubercle of toe IV; (10) metatarsal fold absent; (11) tympanum conspicuous and small; (12) in life, background color of dorsum uniformly ochre; cream or pale brown dorsolateral stripe absent; dark brown lateral stripe from the tip of the snout to the cloacal region; short oblique lateral stripe diffuse, conspicuous pale cream ventrolateral stripe, which may be diffuse at the lower margin; cream paracloacal marks present; (13) in life, dorsal surface of arms orange; dorsal surface of legs ranging from ochre to orange; (14) in life, throat and chest violaceus in non-vocally active males, becoming darker when excited, and yellowish in females; abdomen pale violaceus in males and yellowish in females; (15) in life, iris golden with black vermiculations and pupil ring gold; (16) in preservative, dorsum pale brown; pale brown dorsolateral stripe weakly defined; dark brown lateral stripe from the tip of the snout to the cloacal region; short oblique lateral stripe diffuse, whitish ventrolateral stripe; cream paracloacal marks present (17) in preservative, dorsal surface of arms cream; dorsal surface of legs pale brown; (18) in preservative, throat, chest and abdomen greyish in males and cream in females; (19) advertisement call in general with a continuous emission of 4-pulsed units of repetition, 0.317 s long, on average, followed by silent intervals, and an average dominant frequency of 4.163 kHz; (20) reproductive cycle with the presence of exotrophic tadpoles carried to water bodies, with an unique fin morphology, which begins after the body-tail insertion and is deeper posteriorly to half of the caudal length.
Comparisons with other species. We limited our comparisons to Brazilian congeners (see Appendix 1 for specimens examined) as Allobates nunciatus sp. nov. has a geographical distribution range restricted to the Tapajós River Basin , eastern Amazonia, approximately 700 km from the nearest country border. We also detail the differences of the new species with respect to most closely related species, based on results of our phylogenetic analysis.
Allobates nunciatus sp. nov. occurs syntopically with other three nominal Allobates species: A. femoralis ( Boulenger, 1884) , A. magnussoni Lima, Simões & Kaefer, 2014, and A. tapajos Lima, Simões & Kaefer, 2015. From those species A. nunciatus sp. nov. can be distinguished as follows: from A. femoralis , by a cryptic external coloration in life and preservative, with color spectrum ranging between brown, ochre and cream ( Fig. 6 View FIGURE 6 ) (bright coloration, with white dorsolateral stripes on a black body, and bright red, orange, or yellow marks on thighs in A. femoralis ); from A. tapajos , by a larger size (minimum SVL 19.2 mm in A. nunciatus sp. nov., maximum SVL 16.9 mm in A. tapajos ), a conspicuous pale ventrolateral stripe (absent in A. tapajos ), and a uniformly colored dorsum, with no distinct contrasting brown markings (irregular brown blotches in A. tapajos ); from A. magnussoni , by a conspicuous pale cream ventrolateral stripe (absent in A. magnussoni ), a uniformly colored dorsum, with no distinct contrasting brown markings (brown hourglass-shaped marking in A. magnussoni ), and a continuous well-marked brown lateral stripe (discontinuous brown lateral stripe in A. magnussoni ).
Twenty-two cryptically colored Allobates species are known to occur in Brazil: A. bacurau Simões, 2016 , A. brunneus ( Cope, 1887) , A. caeruleodactylus ( Lima & Caldwell, 2001), A. carajas Simões, Rojas & Lima 2019, A. crombiei ( Morales, 2002) , A. conspicuus ( Morales, 2002) , A. flaviventris Melo-Sampaio, Souza & Peloso, 2013 , A. fuscellus ( Morales, 2002) , A. gasconi ( Morales, 2002) , A. goianus ( Bokermann, 1975) , A. grillisimilis Simões, Sturaro, Peloso & Lima, 2013 , A. juami Simões, Gagliardi-Urrutia, Rojas-Runjaic & Castroviejo-Fish- er, 2018, A. masniger , A. marchesianus ( Melin, 1941) , A. nidicola , A. paleovarzensis Lima, Caldwell, Biavati & Montanarin, 2010, A. olfersioides ( Lutz, 1925) , A. subfolionidificans ( Lima, Sanchez & Souza, 2007), A. sumtuosus ( Morales, 2002) , A. trilineatus ( Boulenger, 1884) , A. tinae Melo-Sampaio, Oliveira & Prates, 2018 , and A. vanzolinius ( Morales, 2002) . The body size of A. nunciatus sp. nov., with minimum SVL 19.2 mm is larg- er than 13 of these species (maximum SVL in mm: 14.9 in A. bacurau , 16.0 in A. grillisimilis and A. sumtuosus , 16.8 in A. caeruleodactylus , 17.0 in A. subfolionidificans and A. marchesianus , 17.2 in A. conspicuus and A. tinae , 17.3 in A. gasconi , 17.4 in A. goianus , 18.0 in A. trilineatus , 18.5 in A. juamii , and 19.1 in A. carajas ).
Besides the size, A. nunciatus sp. nov. is also distinguished by the absence of a pale cream to pale brown conspicuous dorsolateral stripe in living individuals (present in A. bacurau , A. conspicuus , A. gasconi , A. goianus , A. grillisimilis , A. tinae , A. juamii , and A. trilineatus ), a dorsum with no distinct contrasting brown markings in life and preservative (brown dorsal hourglass-shaped markings in A. carajas , A. gasconi , and A. goianus ), living males with white abdominal surfaces and violaceous throats ( Fig. 7 View FIGURE 7 ) (white abdomen and yellow throat in A. sumtuosus , grey abdomen and throat in A. bacurau , yellow abdomen and throat in A. juamii and A. tinae , yellow abdomen and pinkish throat in A. carajas , dark grey throat in A. marchesianus , white throat in A. caeruleodactylus and A. subfolionidificans ), finger III not swollen in adult males (swollen in A. gasconi and A. trilineatus ), and absence of blue dots on fingers of living individuals (present in A. caeruleodactylus ).
Allobates nunciatus sp. nov. is distinguished from similarly sized species ( A. brunneus , A. crombiei , A. flaviventris , A. fuscellus , A. olfersioides , A. paleovarzensis , and A. vanzolinius ) by a uniformly ochre dorsal coloration (uniformly brown in A. fuscellus and A. vanzolinius ; brown hourglass-shaped markings in A. brunneus , A. crombiei , and A. flaviventris ; brown with a darker brown marking on the head in A. paleovarzensis ; and brown X-shaped markings in A. olfersioides ), absence of a distinct pale cream to pale brown continuous dorsolateral line (distinct pale dorsolateral line present in A. fuscellus , A. paleovarzensis , and A. vanzolinius ), white abdominal surfaces in living individuals (greenish-yellow abdomen in A. brunneus ; yellow in A. flaviventris , males of A. crombiei , and females of A. paleovarzensis ), violaceus throat in living males (yellowish throat in A. crombiei and A. flaviventris and white throat in A. olfersioides ) and dorsal surface of thighs ranging from beige to orange in living individuals, similar to dorsum color (greyish dorsal surface of thighs in A. crombiei , different from its brown dorsum).
Two other similarly sized species are closely related and morphologically similar to A. nunciatus sp. nov.: A. masniger and A. nidicola . The new species differs from these by a combination of morphological, acoustic, molecular, reproductive, and distributional data. The external morphology of A. nunciatus sp. nov. differs from theirs by a paler coloration on dorsum of living adult males, ranging from ochre to beige, and ochre-orange limbs (dark brown dorsum and pale brown limbs in A. masniger and A. nidicola ), a conspicuous pale cream ventrolateral stripe (inconspicuous or absent in A. masniger and A. nidicola ), a violaceus throat in living males, which only becomes darker in vocally active individuals (dark grey throat in A. masniger and A. nidicola which does not change in color during non-reproductive period), a weaker sexual dimorphism in dorsal coloration of living specimens (adult males of A. masniger and A. nidicola are darker brown than females), finger II longer than or with a similar size to finger IV (finger II longer than finger IV in A. nidicola ), and toe III length reaches or surpasses the middle of the central sub-articular tubercle of toe IV (toe III length only reaches the posterior margin of the same tubercle in A. nidicola ). In addition to these subtle morphological differences, A. nunciatus sp. nov differs from A. masniger and A. nidicola by a distinct advertisement call, which has a continuous emission of bouts of four closely spaced notes, which we term 4-pulsed units of repetition (UR) (single-pulsed UR in those species; see call section below), approximately 6–10% of genetic distance based on mtDNA genes 16S and COI (see phylogenetic relationships section below), and exotrophic tadpoles carried to water bodies (endotrophic tadpoles developing in terrestrial nests, see tadpole section below). Furthermore, these three species are allopatrically distributed along banks of the Madeira and Tapajós rivers, some of the largest southern tributaries of the Amazon River ( A. nidicola on the west bank of Madeira River, A. masniger in the Madeira-Tapajós interfluve and A. nunciatus sp. nov. on the east bank of Tapajós River ; see geographical distribution below).
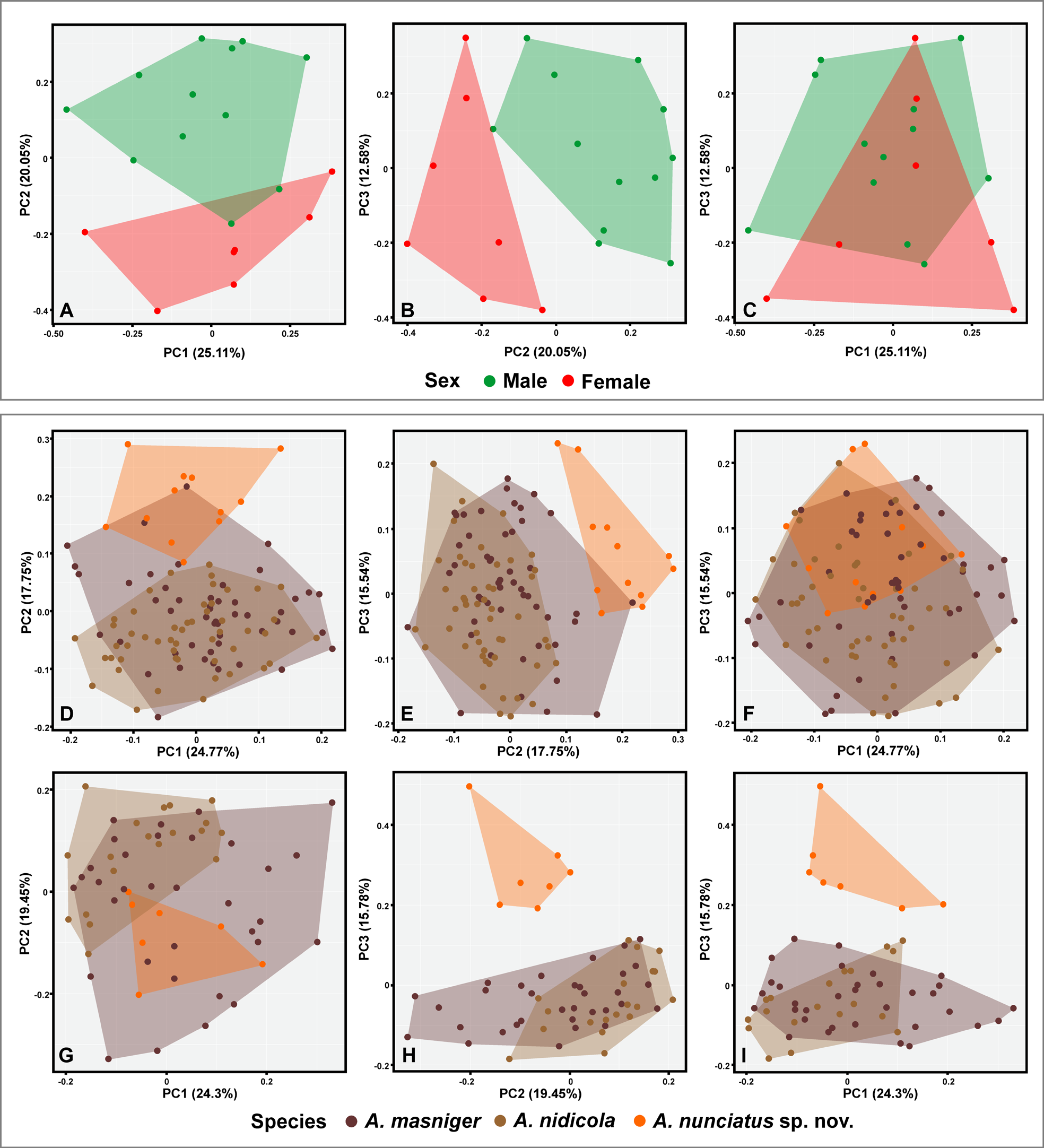
A segregation pattern between clusters and significant differences were detected when we compared body proportions of adult males (Pillai trace = 0.666, df = 216, p <0.001) and females (Pillai trace = 0.924, df = 110, p <0.001) of the new species with proportions of its sister taxa A. masniger and A. nidicola ( Fig. 8 View FIGURE 8 , D–I). In the morphometric multivariate space, A. nunciatus sp. nov. males differed from A. nidicola males when we compared PC1 to PC2 and PC2 to PC3 ( Fig. 8 View FIGURE 8 , D–F). Likewise, A. nunciatus sp. nov. females differed from A. nidicola (comparison between PC1 and PC2) and both sister taxa (when comparing PC1 and PC2 to PC3) ( Fig. 8 View FIGURE 8 , G–I). The first three axis summarized 24.77%, 17.75%, and 15.54% of the variation in sister taxa for males and 24.3%, 19.45%, and 15.78% for females. The highest loading values of sister taxa variation regarding males came from snout-vent length (PC1) and the morphometric ratios of hand length (PC2), tibia length (PC1, PC2), head length (PC3), foot length (PC1), tympanum diameter (PC2), head width (PC3) and toe disc width (PC3). The highest loading values of sister taxa variation for females came from snout-vent length (PC1) and the morphometric ratios of hand length (PC2), head length (PC1), foot length (PC3), head width (PC1), finger disc width (PC2, PC3), and toe disc width (PC2, PC3). Despite these significant differences, highest influenced morphometric characters in loading values showed some degree of measurement overlap or very subtle dissimilarities, indicating that morphometric measurements and ratios used as diagnostic characters between sexes and sister species should be treated with caution. See Appendix 2 View APPENDIX 2 for complete loading values.
Furthermore, the advertisement call of A. nunciatus sp. nov. (with a continuous emission of mostly 4-pulsed units of repetition (UR), followed by silent intervals ( Fig. 9 View FIGURE 9 , 10 View FIGURE 10 )) distinguishes this species from most Brazilian cryptically colored Allobates ( Table 3 View TABLE 3 ), which have advertisement calls in series of trills ( A. bacurau , A. crombiei , A. gasconi , A. grillisimilis , A. marchesianus , A. paleovarzensis , A. juami , A. sumtuosus , A. olfersioides from the state of Alagoas), double-pulsed UR or trills ( A. flaviventris , A. tapajos ), or single-pulsed UR ( A. brunneus , A. caeruleodactylus , A. goianus , A. marchesianus , A. magnussoni , A. subfolionidificans , A. trilineatus , A. tinae , A. olfersioides from the states of Bahia and Rio de Janeiro; Caldwell et al. 2002; Carvalho et al. 2016; Forti et al. 2017; Grant & Rodriguez 2001; Kaefer & Lima 2012; Lima & Caldwell 2001; Lima et al. 2007, 2009, 2010, 2012, 2014, 2015; Melo-Sampaio et al. 2018; Simões 2016; Simões et al. 2013a, b, 2018; Tsuji-Nishikido et al. 2012). Despite the differences in call structure between the related species A. masniger and A. nidicola , A. nunciatus sp. nov. calls at a similar range and dominant frequency ( Table 3 View TABLE 3 ), and they share an ascending frequency modulation of notes. In relation to Brazilian brightly colored Allobates , the call structure (emission of 4-pulsed UR) distinguishes A. nunciatus sp. nov. from the allopatrically distributed A. hodli , which emits 2-pulsed UR ( Simões et al. 2010) and A. myersi ( Pyburn, 1981) , which emits 6- or 8-pulsed UR ( Simões & Lima 2011), but not from the sympatric A. femoralis . However, the call of A. nunciatus sp. nov. is much faster (mean of 5.16 notes/s) and emitted at a higher dominant frequency of 4.16 ± 0.18 (3.84–4.44) kHz compared to syntopic A. femoralis calls, which has a mean rate of 4.19 notes/s and frequency of 3.21 ± 0.08 (3.10–3.33) kHz ( Table 3 View TABLE 3 ). The three remaining Brazilian Allobates species ( A. conspicuus , A. fuscellus , and A. vanzolinius ) have no advertisement call description ( Guerra et al. 2018).
Description of the holotype. Adult male ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ), 21.4 mm SVL. Dorsum mostly smooth, with two rows of small granules from the distal margin of eye to urostyle, granules more abundant in the distal region of dorsum; throat and abdomen smooth ( Fig. 2 View FIGURE 2 ). Head slightly longer than wide (HW: 98% of HL; HL: 33% of SVL); snout truncated in dorsal view and rounded in lateral view, slightly shorter than eye diameter; canthus rostralis indistinct; loreal region obtuse; tympanum distinct, tympanum diameter <half of eye diameter; eye diameter: 36% of HL; tongue longer than wide, not adhered to floor of mouth posteriorly; choanae relatively small, round; vomerine odontophores absent. Upper arm and forearm with approximately the same size; hand slightly shorter than upper arm; relative length of fingers IV = II <I <III; fingers with moderately expanded, rounded tips with distinct dorsal scutes; metacarpal tubercles present, inner metacarpal tubercles small and oval and outer metacarpal tubercles large and rounded; sub-articular tubercles present and rounded, one in fingers I, II, and IV and two in finger III; finger III not swollen; basal webbing and lateral fringes absent on fingers ( Fig. 3 View FIGURE 3 ). Thigh slightly longer than tibia, foot shorter than thigh and tibia; relative length of toes: I <II <V<III <IV; toe III surpassing the mid-level of the central subarticular tubercle of toe IV; basal webbing present between toes III and IV, and rudimentary webbing between toes II and III; toes II, III, and IV with expanded rounded tips; metatarsal tubercles present, oval, inner metatarsal tubercle slightly larger than outer ones; metatarsal fold absent; tarsal keel in form of a curved tubercle, not connected with metatarsal fold; sub-articular tubercles present, rounded; one sub-articular tubercle on toes I and II, two on toes III and V, and three on toe IV, with proximal and distal tubercles less evident ( Fig. 4 View FIGURE 4 ). For holotype measurements see Table 1 View TABLE 1 .
Coloration of the holotype in life. Ochre on the dorsal surface of the snout, anteriorly to the interorbital region and close to the dark brown lateral stripes, and a darker ochre, rectangular area at the center of dorsum, with some darker small dots scattered, following granule rows. Pair of cream paracloacal marks present. A continuous dark brown stripe extends from the tip of snout to flanks and reaches the cloacal region. A cream ventrolateral stripe runs below this dark line, with diffuse limits ventrally. Dorsal surfaces of arms orange, paler than the dorsum color; dorsal surfaces of legs ochre, with a similar color to the dorsum. Weakly marked single transversal brown bar on the central dorsal surface of tibia and thigh. Dorsal surfaces of hands and feet with punctuate white dots. Throat and chest violaceus, and abdomen pale violaceus to slightly transparent, revealing a white peritoneum. Ventral surfaces of forearms, palms of hands, and soles of feet brown.Iris golden with black vermiculations and pupil black, circled by a golden ring.
Coloration of holotype in preservative. Dorsal background surface of body and legs pale brown, with a darker brown rectangular region at the center of the dorsum ( Fig. 2 View FIGURE 2 ). Paler cream markings at the dorsal paracloacal region. Dorsal surfaces of arms cream, paler than the dorsum. A dark brown stripe from the tip of snout to cloacal region. White dots at dorsal surfaces of hands and feet. Weakly marked transversal brown bars above brown dorsal surfaces of thighs and tibia. On ventral surface, brown pigmentation on the throat, chest, and anterior abdomen; posterior abdomen, thighs, and tibia cream. Palmar surface of hands and plantar surface of feet uniformly brown ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ).
Variation. Adults show a weak sexual dimorphism in dorsal coloration ( Fig. 6 View FIGURE 6 ), but on the ventral surface, the differences can be prominent, with males presenting violaceus throats and females white-yellowish ( Figs. 5 View FIGURE 5 , 7 View FIGURE 7 ). Some individuals have a narrow golden stripe on the snout region, above the lateral dark brown stripe, anteriorly to the eye-level. The definition of the pale ventrolateral stripe towards the ventral area varies between individuals but is always present. A weak ontogenetic variation occurs, with some color details more pronounced in juveniles, such as dark transversal bars in legs and a darker region at the center of dorsum, contrasting with adjacent colors ( Fig. 6 View FIGURE 6 ).
The sexual dimorphism based on body proportions (morphometric ratios) showed a degree of segregation of both sexes in the morphometric multivariate space ( Fig. 8 View FIGURE 8 , A–C) when PC1 was compared to PC2 and PC2 to PC3, showing significant differences between males and females (Pillai trace = 0.723, df = 15, p <0.001). The first three PCA axis summarized 25.11%, 20.05%, and 12.58% of the variation. The highest loading values represent snoutvent length (PC3) and the morphometric ratios (measurement/SVL) of hand length (PC1), tibia length (PC1), first finger length (PC1), thigh length (PC2); eye diameter (PC3); interorbital distance (IO); finger disc width (PC2) and toe disc width (PC2).
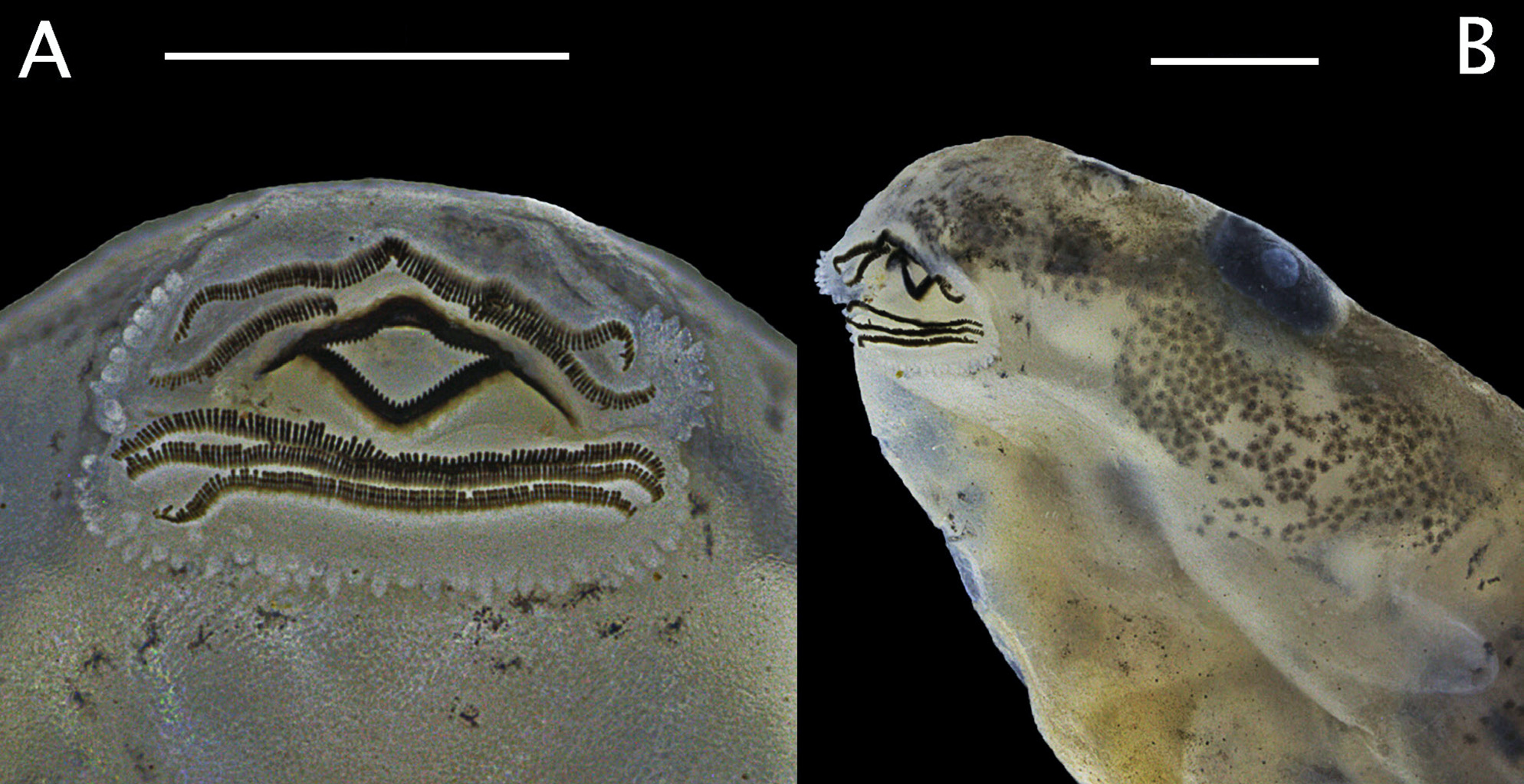
Tadpole description. Qualitative characters based on 12 tadpoles raised to developmental stages 36, 37, and 39 (lot INPA-H 40487, field number APL 22371). Quantitative characters based exclusively on larvae at stage 37 (n = 6; Table 2 View TABLE 2 ). Body ellipsoid, longer than wide, acuminate anteriorly and rounded posteriorly in dorsal view, flattened in lateral view ( Fig. 11 View FIGURE 11 ). Body length 32% of TL and tail length 68% of TL; HWLE 80% of BW; BH 52% of BW; snout rounded in dorsal view and truncate in lateral view; eyes dorsal and directed laterally; END 97% of ED; mean ED 0.96 ± 0.06 mm; IOD 62% of HWLE. Small naris located dorsolaterally and directed laterally; mean internarial distance 1.54 ± 0.09 mm. Fleshy ring present on the inner margin of nostrils, round and straight, not ornamented. Sinistral spiracle as a free tube at mid-body, measuring 0.21 ± 0.07 mm. Average length of dextral vent tube 0.95 ± 0.10 mm. Dorsal fin begins after the body-tail insertion, with dorsal edge shallow and straight along approximately half of the caudal length, deeper posteriorly. Lower fin similar to the dorsal fin in structure and depth. Tail tip acuminate. Caudal musculature bifurcated dorsally, reaching half of the body. Anteroventral oral apparatus ( Fig. 12 View FIGURE 12 ), slightly emarginated laterally, transversely elliptical, 1.53 ± 0.16 mm wide. Anterior labium with groups of eight to nine short and round papillae distributed in a single row on each lateral margin. Posterior labium with a single row of short marginal papillae with similar length. Upper jaw sheath arch-shaped, slightly curved anteriorly in the middle, longer than the lower jaw sheath. Lower jaw sheath V-shaped, as deep as the upper jaw sheath. Cutting edge of upper and lower jaw sheaths serrated along the entire length. Labial tooth row formula 2(2)/3(1); tooth row A-1 complete, measuring 1.54 ± 0.17 mm in length; tooth row A-2 interrupted medially, consisting of two widely sepa- rated rows at the level of upper jaw sheath, segments measuring 0.59 ± 0.07 mm in length and a gap measuring 0.33 ± 0.06 mm. Posterior tooth rows P-1 and P-2 with approximately the same length as row A-1 (means: 1.44 ± 0.20 and 1.52 ± 0.15 mm, respectively), both slightly longer than the complete P-3 (1.32 ± 0.10 mm). Such a structure of oral disc ( Fig. 12 View FIGURE 12 ) was constant throughout all developmental stages examined.
Coloration of preserved tadpoles. Dorsum, anterior part of the ventral area, and lateral surfaces with a pale cream background color, densely covered with dark brownish-grey melanophores ( Fig. 11 View FIGURE 11 ). Background color appears as irregular pale cream blotches or spots, more frequent on tail surface. Posterior part of the ventral area translucent, with intestines visible through the skin. Tail musculature cream. Tail fins translucent, with few irregular brown blotches.
Comparison with tadpoles of other species. The presence of an oral disc and a spiracle ( Figs. 11 View FIGURE 11 , 12 View FIGURE 12 ) clearly distinguishes the exotrophic tadpoles of A. nunciatus sp. nov. from endotrophic tadpoles of closely related species, such as A. nidicola and A. masniger , which lack those structures and develop entirely in terrestrial nests. Allobates nunciatus sp. nov. differs from all described Allobates tadpoles: from A. caeruleodactylus , A. marchesianus , A. grillisimilis , and A. tapajos by a posterior labium with short papillae of similar size (those species have a posterior labium partially surrounded by long marginal papillae, variable in length; Caldwell et al. 2002; Simões et al. 2013a; Lima et al. 2015); from A. granti (Kok, MacCulloch, Gaucher, Poelman, Bourne, Lathrop, & Lenglet, 2006), A. hodli , A. subfolionidificans , A. brunneus , A. paleovarzensis , A. sumtuosus , and A. magnussoni ( Lima et al. 2007, 2009, 2010, 2014; Simões & Lima, 2012) by the presence of labial tooth row P-3 (absent in A. granti ), the absence of a distinct brown longitudinal bar from the snout to the mid-body (present in A. sumtuosus ), the presence of similar-sized papillae on anterior and posterior labium (posterior papillae slightly larger in A. subfolionidificans ), almost subequal tooth rows P-1, P-2, and P-3 (P-3 distinctly shorter than others in A. brunneus , A. magnussoni , and A. paleovarzensis ), and the presence of pale cream blotches or spots along the body and tail (uniform brownish-yellow coloration in A. hodli ). Furthermore, the fin morphology, beginning after the body-tail insertion, but deeper poste- riorly to half of the caudal length is not shared by any other known Allobates larvae (fins increase in size gradually after the body-tail insertion).
Advertisement call and variation. Allobates nunciatus sp. nov. has a complex advertisement call, with a continuous emission of short tonal notes, usually arranged in bouts of four closely spaced notes, which we term 4-pulsed units of repetition (UR), followed by silent intervals ( Figs. 9 View FIGURE 9 , 10 View FIGURE 10 ). Such an arrangement in four notes is the most frequently emitted by excited males, corresponding to 356 of 527 (67.5%) arrangements analyzed in nine distinct recordings. The second most frequent arrangement was the emission of 3-pulsed UR (14.2%). The species also emits arrangements in 2- (6.6%), 5- (3.4%), 6- (6%), 8- (1.5%), and 10-pulsed UR (the rarest arrangement, 0.6%) ( Fig. 10 View FIGURE 10 ). The 2-pulsed UR is the most frequent arrangement when males start to call and can be considered a warming call. Disturbed males interrupt the main series of 3- to 10-pulsed UR and restart emitting 2-pulsed UR. Despite the note arrangement variation, the spectral and temporal characteristics of notes are constant in all analyzed advertisement calls. Note duration lasts 0.031 ± 0.006 (0.025 –0.045) s, with a low- er frequency of 3.60 ± 0.34 (3.21–4.06) kHz and higher frequency of 4.60 ± 0.13 (4.30–4.79) kHz. Four-pulsed UR are 0.317 ± 0.034 (0.256 –0.357) s long, and with two types of inter-note intervals, two short with 0.040 ± 0.005 (0.029 –0.047) s, interpolated with a long one lasting 0.076 ± 0.007 (0.064 –0.085) s. Silent intervals between UR lasts 0.360 ± 0.078 (0.257 –0.581) s. This pattern is observable in all paired arrangements ( Figs. 9 View FIGURE 9 , 10 View FIGURE 10 ).
Geographical distribution and natural history. Allobates nunciatus sp. nov. is the most abundant Allobates species in non-flooded (terra firme) forests on the east bank of Middle Tapajós River ( Figs. 13 View FIGURE 13 , 14 View FIGURE 14 ), and might be geographically restricted to this bank and river basin. It also inhabits riparian habitats of small streams but is rarely recorded in igapó forests, periodically flooded by large rivers. The species occurs on both banks of the large tributary Jamanxim River ( Figs. 13 View FIGURE 13 , 14 View FIGURE 14 ), with no evident morphological, acoustic or molecular distinction between these two separate populations.
Alobates nunciatus sp. nov. is diurnal and breeds ( Fig. 15 View FIGURE 15 ) during the rainy season (from November to March), mostly in the beginning of rains (November and December). However, a higher number of individuals, comprising adults and juveniles, were spotted during the dry season (September and October), which indicates a high spatial movement in this period. Males call during the day, mostly in the morning and late afternoon. Most of the visually recorded individuals were in the leaf litter ( Fig. 15 View FIGURE 15 ). Eggs were deposited in a gelatinous mass inside nests made of dry fallen leaves on the leaf litter ( Fig. 15 View FIGURE 15 ). The eggs developed into exotrophic tadpoles, which were then carried by the males to nearby water bodies. This reproductive strategy differs from that of the closely related A. nidicola and A. masniger , whose entire development of endotrophic tadpoles takes place in forest floor nests ( Caldwell & Lima 2003; Tsuji-Nishikido et al. 2012).












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.