
Telephryxus clypeus, Williams & Boyko, 2021
|
publication ID |
https://doi.org/ 10.5252/zoosystema2021v43a4 |
|
publication LSID |
urn:lsid:zoobank.org:pub:4F2A16F1-B100-4236-AD31-945896D6F910 |
|
DOI |
https://doi.org/10.5281/zenodo.4555526 |
|
persistent identifier |
https://treatment.plazi.org/id/B21DCDDB-9E3D-4380-A7E8-C0968B50C99C |
|
taxon LSID |
lsid:zoobank.org:act:B21DCDDB-9E3D-4380-A7E8-C0968B50C99C |
|
treatment provided by |
Felipe |
|
scientific name |
Telephryxus clypeus |
| status |
sp. nov. |
Telephryxus clypeus View in CoL n. sp.
( Figs 6-10 View FIG View FIG View FIG View FIG View FIG ; 11N View FIG )
urn:lsid:zoobank.org:act:B21DCDDB-9E3D-4380-A7E8-C0968B50C99C
“remarkable isopod parasite ( Dajidae View in CoL ?)” – Gore 1983: 207.
TYPE MATERIAL. — Holotype. Caribbean Sea • USNM 1163461 View Materials ; female (8.0 mm diameter) with pre-molt epicaridean larvae, attached to right antennule of female Munidopsis crassa Smith, 1885 (40.9 mm CL, incl. rostrum; USNM 204595 View Materials ); NORDA Sta. 99; 14°51.70’N, 67°27.8’W; Venezuela Basin, north of Islas de Aves ; 4956-4997 m; coll. United States Navy research vessel USNS Bartlett, taken by 45 ft balloon net trawl ( Gore 1983); 3.XII.1981. GoogleMaps
Allotype. Caribbean Sea • USNM 11616635 View Materials ; mature male (2.4 mm L); same data as for holotype. GoogleMaps
Paratype. Caribbean Sea • USNM 11616636 View Materials ; cryptoniscus larva (0.9 mm L); same data as for holotype GoogleMaps .
ADDITIONAL MATERIAL. — USNM 11616637, c. 30 pre-molt epicaridium larvae, same data as for holotype (on SEM stubs).
TYPE LOCALITY. — 14°51.70’N, 67°27.8’W, Venezuela Basin, Caribbean Sea, 4956-4997 m (fide Gore, 1983).
TYPE HOST. — Munidopsis crassa Smith, 1885 [Crustacea: Anomura: Munidopsidae ].
ETYMOLOGY. — The species name is derived from the Latin for “shield” in reference to the broad subquadrate plate that partially surrounds the host antennule and is reminiscent of the shape of a shield. The gender is masculine.
DISTRIBUTION. — Known only from the type locality and type host.
DESCRIPTION
Female
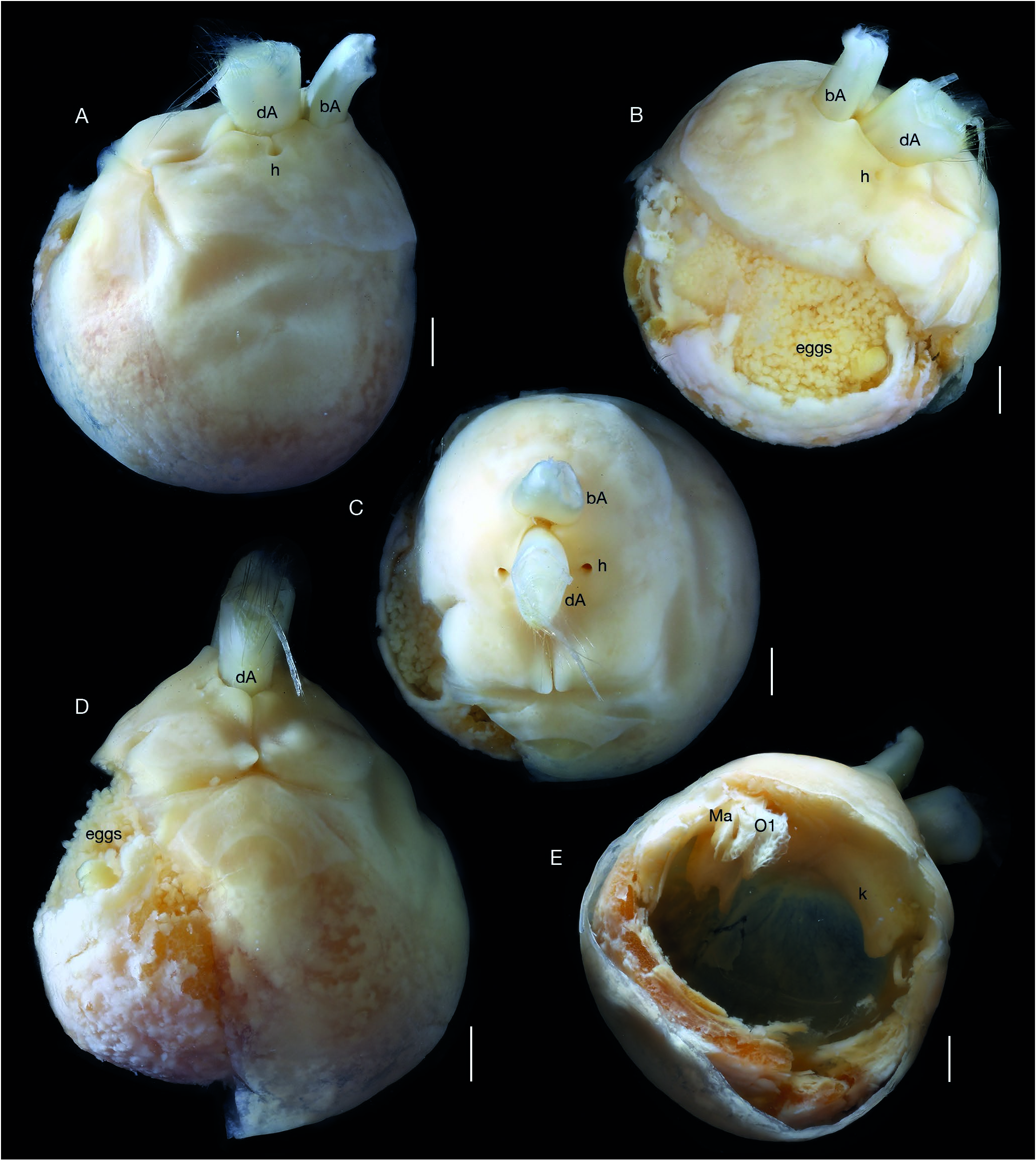
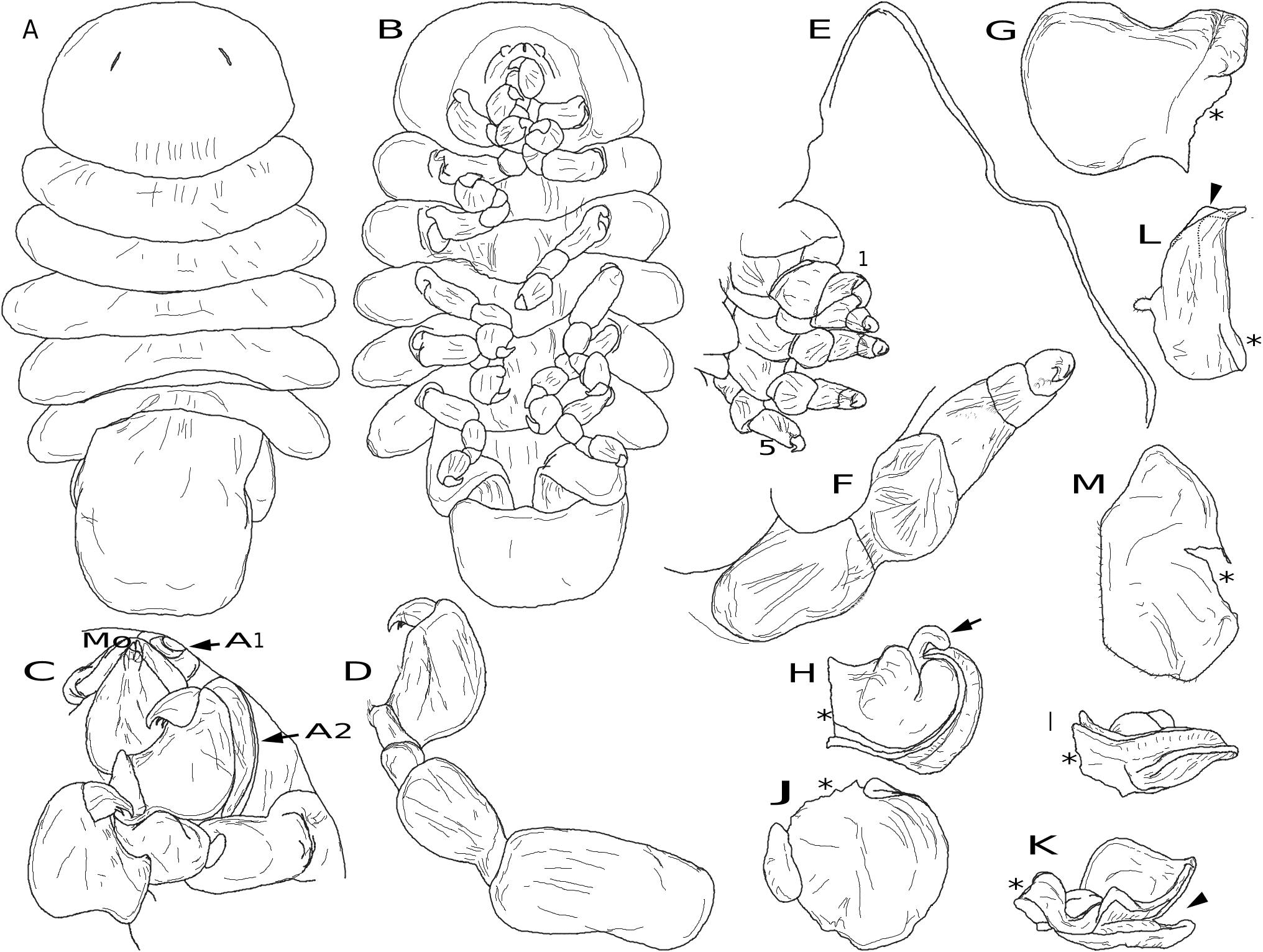
Body spheroid ( Figs 6 View FIG ; 7A, B View FIG ), length and width nearly equal, filled with numerous pre-molt epicaridium larvae (see description below). Cephalon externally indistinguishable from pereon, without eyes. Antennules ( Fig. 7F View FIG ) each as flat triangular plate covered with minute scales (not shown); antennae each as rectangular flat plate lateral to oral cone ( Fig. 7E, G View FIG ), covered with minute scales ( Fig. 7G View FIG inset). Oral cone rounded ( Fig. 7E View FIG ); mouthparts indistinct. Maxillipeds inflated, rectangular, each with recurved maxilliped digitiform extension (“appendix”) at posterolateral corner ( Figs 6E View FIG , 7E, H, K View FIG ), “appendix” extending into groove of oostegite 1 ( Fig. 7K View FIG ). Pereopods 1-5 subequal in size and shape, without setae ( Fig. 7E View FIG ); dactylus short, recurved, propodus carpus and merus fused (indistinct ventral indication of segmentation on some pereopods), ischia and bases stout. Oostegite 1 largest, broadly ovate with large triangular posterior accessory lobe ( Figs 6E View FIG ; 7I, J, K View FIG ), broad rounded lobe medially divided in lateral view, forming groove ( Fig. 7J, K View FIG ); oostegites 2-4 flat, ovate, posterior pairs progressively slightly larger ( Fig. 7K View FIG ); oostegite 5 flat, elongate, tapering, lacking marginal setae ( Fig. 5L View FIG ). Pleon presumably modified (see Discussion) as subquadrate, broad, thickened plate ( Figs 6 View FIG A-D; 7A, B, D) partially surrounding host antennule with two circular medial holes, subequal in size; hole surrounding basal antennular peduncle of host closest to mouthparts of parasite, hole surrounding distal portion of host antennular peduncle with one small additional hole on each lateral margin.
Male
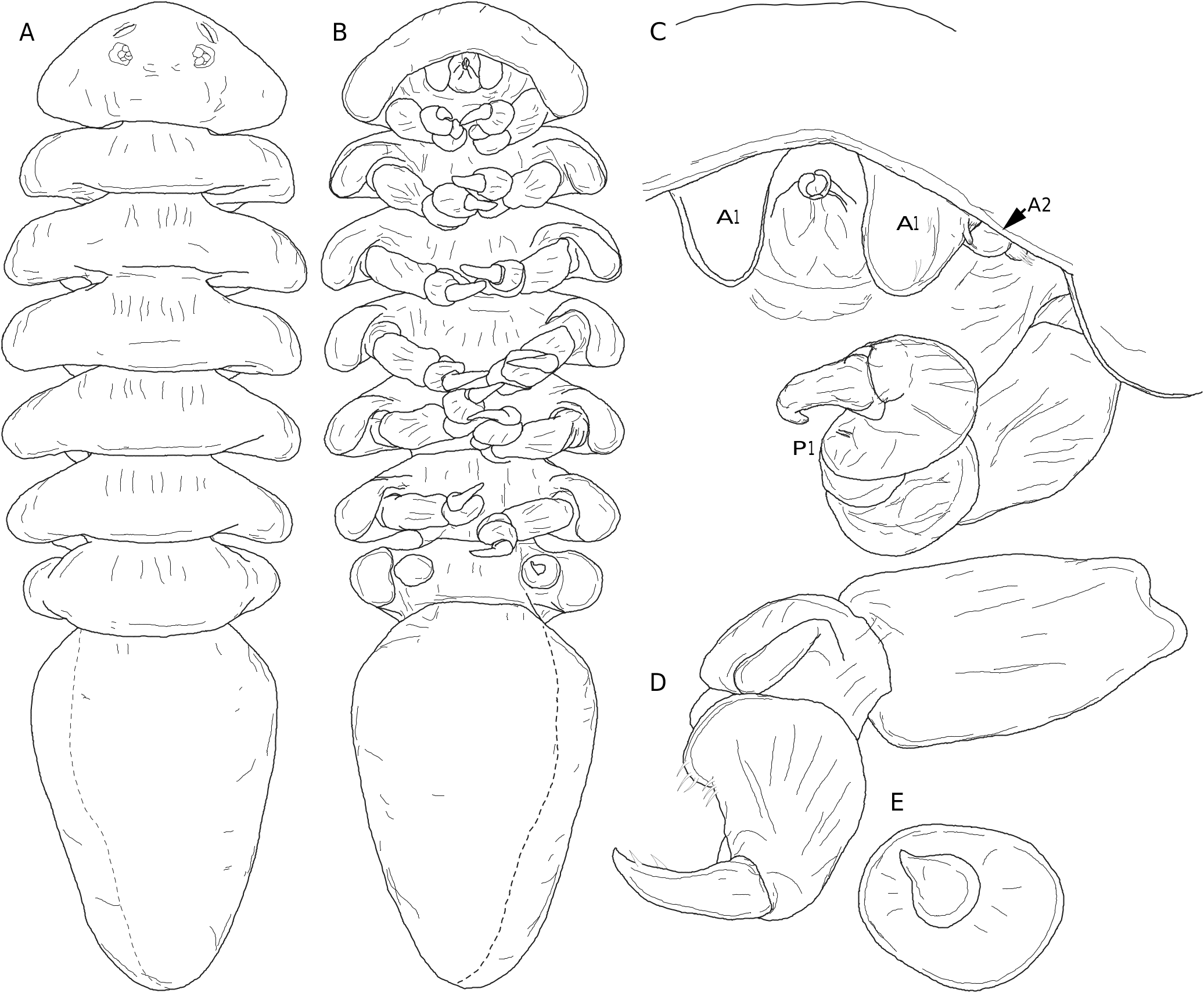
Body not recurved ventrally ( Fig. 8A, B View FIG ). Cephalon fused with pereomere 1 ( Fig. 8 View FIG A-C), anterior margin subtriangular, posterolateral margins evenly rounded; faint unpigmented eyes, cephalic slits present. Antennules each as single ovate plate lateral to and extending posterior to oral cone, with minute lateral projection bearing terminal setae ( Fig. 8C View FIG ); antennae lateral to antennules, of two segments each with distal setae ( Fig. 8C View FIG ), flagella absent. Oral cone subtriangular ( Fig. 8B, C View FIG ). Pereomeres 2-7 distinct, 2-6 subequal in width, 7 slightly narrower, lateral margins recurved ventrally ( Fig. 8A, B View FIG ). Pereopods 1-6 subequal in size and shape ( Fig. 8 View FIG B-D), ischia and bases fused, carpi rounded, ischia/bases elongate; dactylus and propodus with isolated marginal setae; pereopod 7 reduced, single rounded stub on right side, rounded stub plus dactylus on left side. Pleon elongate ( Fig. 8A, B View FIG ), tapering to rounded tip, all segments fused, distinct from pereomere 7; anal slit and pleopods lacking.
Cryptoniscus larva
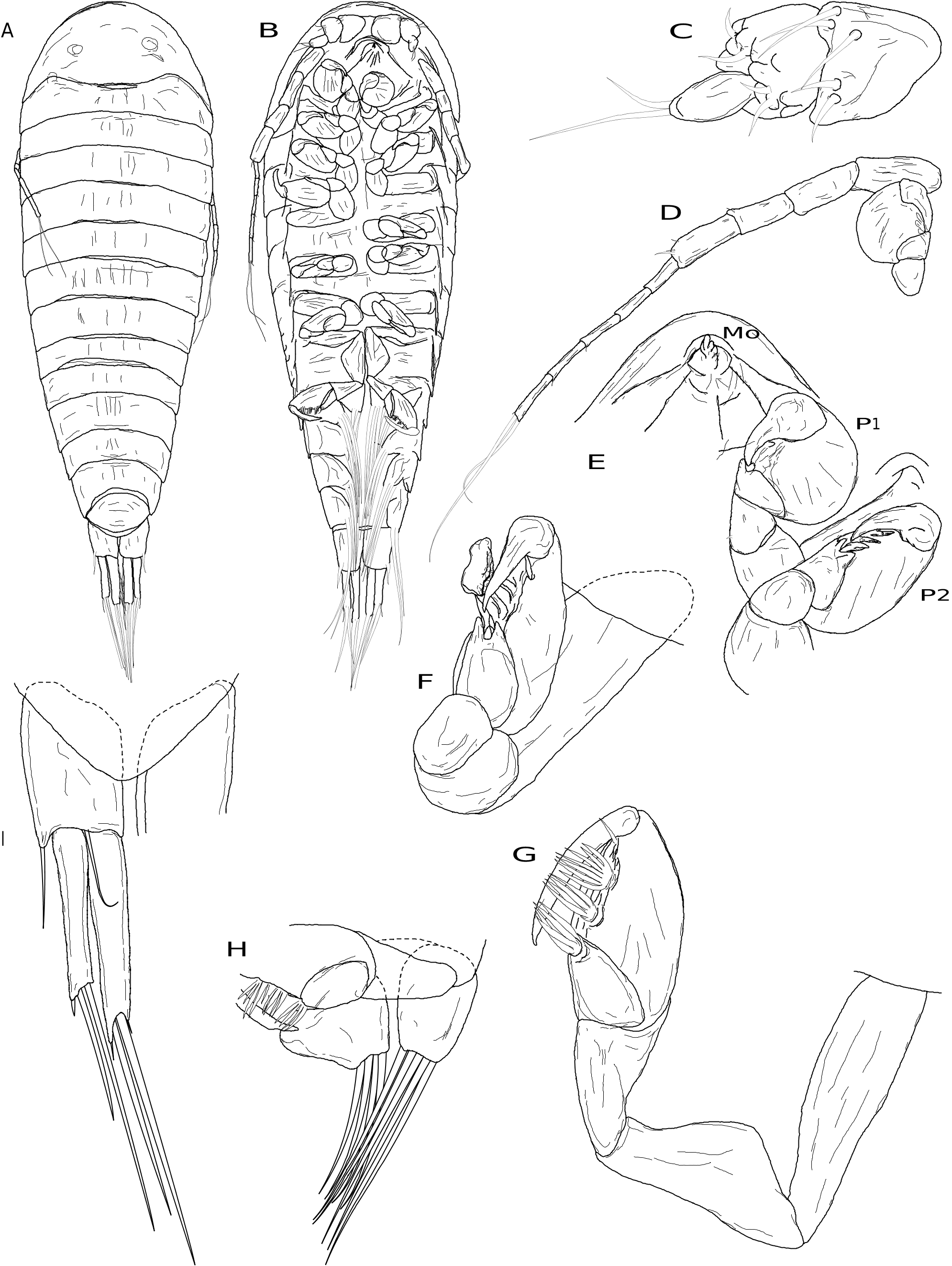
Body tear-drop shaped ( Fig. 9A, B View FIG ), length 0.9 mm, maximum width at pereomere 3. Cephalon anterior margin round, medial region of posterior margin convex, lateral regions concave, posterolateral margins extended posteriorly ( Fig. 9A View FIG ); eyes round, unpigmented. Body pigmentation lacking. Antennules of three articles each ( Fig. 9C View FIG ), basal article triangular with five stout distal setae, article 2 quadrate with four stout distal setae and several low, rounded bumps, article 3 digitiform, inserted into article 2 distoventrally, less than half width of article 2, with two distal setae ( Fig. 9C View FIG ). Antennae of nine articles each (four peduncular and five flagellar) ( Fig. 9B, D View FIG ), all articles cylindrical, peduncular articles subequal in size with minute, distal setae (at least on articles 3 and 4); flagellar articles approximately half width of peduncular articles, with minute terminal setae, distalmost article with two long terminal setae ( Fig. 9D View FIG ). Oral cone triangular, anteriorly directed, lacking oral sucker (see Remarks) ( Fig. 9B, E View FIG ). Pereomeres 1-7 with entire (not toothed) coxal plates ( Fig. 9B View FIG ). Pereopod 1 with short, slightly curved dactylus, propodus semi-spherical with distoventral ridge corresponding to dactylus tip position bearing stout, simple setae; carpus with distal seta; merus and ischium rounded, basis cylindrical ( Fig. 9D, E View FIG ). Pereopods 2-6 with more elongate curved dactyli, propodi progressively more elongate with distoventral ridge corresponding to dactylus tip position bearing stout, simple setae; each carpus with distal seta; meri and ischia rounded, bases cylindrical ( Fig. 9E, F View FIG ). Pereopod 7 with long, slightly curved dactylus, propodus elongate with distoventral ridge corresponding to dactylus tip position, with two large multifid setae and one simple seta near base of dactylus; carpus with distal large multifid seta; merus triangular with anterodorsal edge extending into spine-like extension closely applied to propodus dorsal margin, ischium large, triangular, basis long, cylindrical ( Fig. 9G View FIG ). Pleon with five pairs of biramous pleopods, endopods cylindrical, exopods triangular, both with long terminal setae ( Fig. 9H View FIG ). Pleotelson oval, with rounded distomedial margin ( Fig. 9A, I View FIG ). Uropods biramous, composed of wide sympod with lateral projection and seta, endopod slightly longer than exopod, pair of long distal setae on endopods and exopods, short seta at distolateral margin of endopods and exopods ( Fig. 9I View FIG ).
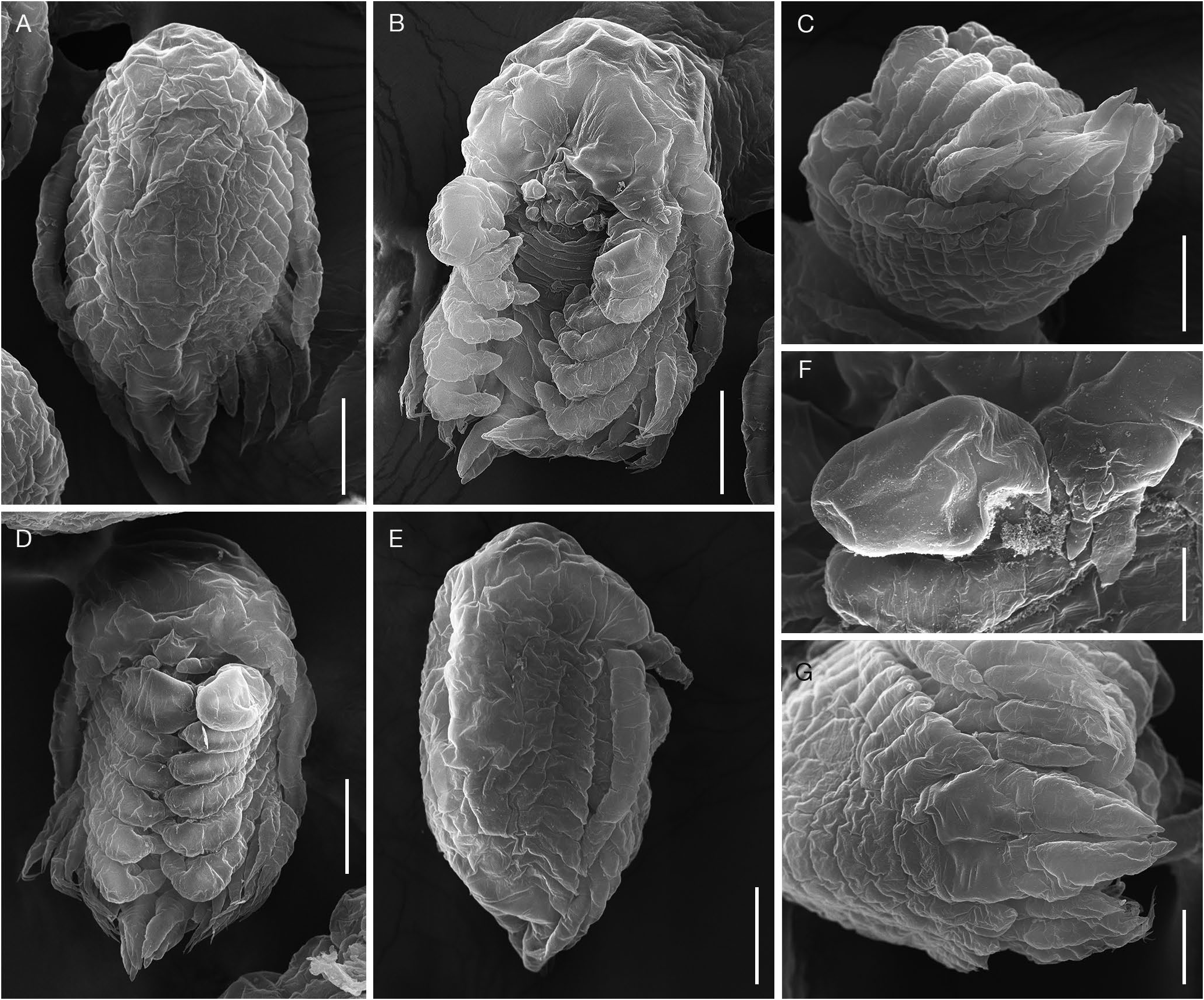
Pre-Molt Epicaridium Larva
Length 228.1 ± 13.7 µm (n= 30) from anterior margin of cephalon to end of pleotelson. Body ovoid ( Fig. 10A View FIG ), covered in wrinkled cuticle, obscuring segmentation of appendages. Cephalon large, rounded anteriorly; in ventral view, broad and extending laterally, appearing fused with antennules ( Fig. 10B, D View FIG ). Antennule triangular with large basal portion, two distal lobes and additional smaller extensions ( Fig. 10B, D F View FIG ). Antenna long, approximately ¾ length of body ( Fig. 10A, B, E View FIG ) with small distal lobes ( Fig. 10C, G View FIG ). Oral region inflated, no distinct oral cone, with pair of lobes (maxillipeds) anterior to pair of smaller lobes ( Fig. 10B, D View FIG ). Six pairs of rounded, broadly hooked pereopods, subequal in size, segmentation not visible ( Fig. 10B, D, E View FIG ). In lateral view, sides of pereomeres and pleomeres with thin extensions; anterior pleomeres 3-5 with crenulate margins ( Fig. 10E, G View FIG ). Pleon with 5 pairs of pleopods ( Fig. 10B, C, G View FIG ); pleopods 1-4 biramous, bearing three long terminal setae on each exopod and two long terminal setae on each endopod; pleopod 5 reduced, uniramous, lacking long terminal setae. Uropods biramous, endopods slightly shorter than exopods, both ending in two short lobes, terminal setae lacking ( Fig. 10D, G View FIG ).
REMARKS
See Remarks under Akrophryxus milvus n. gen., n. sp. for comparison with Telephryxus clypeus n. gen., n. sp. and other dajids. The maxilliped digitiform extension (“appendix” of Rustad 1935) appears homologous with the lateral lobes of the barbula found in bopyrids ( Markham 1985). As in bopyrids, the maxillipeds are used to pump water through the brood chamber, oxygenating the eggs or larvae ( Gilson 1909; Cericola & Williams 2015). However, in the case of females of species in the two new genera, the digitiform extension of the maxilliped also fits into the groove of oostegite 1, thus further aiding in oxygenation by moving the whole first oostegite. It may also aid in maintaining larvae in the brood chamber prior to release ( Cericola & Williams 2015).
The epicaridium larvae of Telephryxus clypeus n. gen., n. sp. were in pre-molt, having a wrinkled cuticle (particularly evident surrounding the cephalon, antennae and pereopods) similar to that shown in larvae of other dajid species (G. O. Sars 1898: pl. 94; Gilson 1909: figs 10, 11; Shimomura et al. 2005: fig. 12). As discussed by Gilson (1909), there are two stages of epicaridium larvae: the first is maintained within the brood chamber of the female and the second is released from the female as the planktonic phase prior to attachment to a copepod intermediate host ( Fig. 11 View FIG A-G). In addition to the yolk provisioned endogenously to the epicaridium larvae, they may also be provisioned with exogenous sources of nutrition taken up across the embryonic epithelial layer (as discussed by Strömberg [1971] and shown to occur in cryptoniscoids by Goudeau [1977]) that fuel their final development prior to release into the water column. After molting, the second stage (mature) epicaridium larvae of dajids show distinct segmentation of appendages (e.g. G. O. Sars 1898; Gilson 1909; Tattersal 1911; Taberly 1957a). The pleopods of the epicaridium larvae of T. clypeus n. gen., n. sp. all appear biramous (as they do in other pre-molt larvae previously described; e.g. Stephensen [1913], Shimomura et al. [2005]). However, it is not clear if any or all of these will remain biramous or become uniramous in the mature epicaridium larvae. Pleopods of mature epicaridium larvae of dajids have been described with both uniramous and biramous states: pleopods 1-5 uniramous ( Gilson 1909) or pleopods 1-4 biramous and 5 uniramous ( Taberly 1957a; Coyle & Mueller 1981).
The uropods of the pre-molt epicaridium larvae of T. clypeus n. gen., n. sp. are biramous, as are those of the mature epicaridium larvae described by Taberly (1957a). However, the orientation of the endopod and exopod of the uropods is ninety degrees rotated (“superposées” of Trilles 1999) from the lateral orientation that is typical for other epicarideans (“côte à côte” of Trilles 1999). The two segments can only be distinguished in lateral view ( Taberly 1957a: fig. 1; Coyle & Mueller 1981: fig. 1E). Some mature dajid larvae have been illustrated with what appear to be uniramous uropods (G. O. Sars 1898; Gilson 1909) but, as suggested by Taberly (1957a), these larvae may not have been observed in the proper orientation. Tattersall (1911) considered uniramous pleopods and uniramous uropods a defining characteristic for dajids; however, the key of Bourdon in Trilles (1999) is probably more accurate for dajid characters (pleopods and uropods both biramous).
The cryptoniscus larva of T. clypeus n. gen., n. sp. lacks an oral sucker, a characteristic found in most dajid species for which larvae have been examined ( Fig. 11J View FIG ). However, we think this represents a case where the attachment structure fell off prior to or during collection. As noted by Taberly (1957b), the oral sucker of dajids is easily detached. Other records of dajid cryptoniscus larvae lacking an oral sucker (e.g. Holophryxus alaskensis Richardson, 1905 , Zonophryxus quinquidens Barnard, 1914 ) should be considered as possible instances where this structure was detached. Although Coyle & Mueller (1981) collected several cryptoniscus larvae of H. alaskensis and reported they lacked an oral sucker, we suggest collection of new material of all such species is needed to confirm absence or presence of the oral sucker in cryptoniscus larvae across dajid taxa.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Telephryxus clypeus
| Williams, Jason D. & Boyko, Christopher B. 2021 |
Dajidae
| G. O. Sars 1883 |