
Cyrtodactylus dumnuii, Bauer, Aaron M., Kunya, Kirati, Sumontha, Montri, Niyomwan, Piyawan, Pauwels, Olivier S. G., Chanhome, Lawan & Kunya, Tunyakorn, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.197372 |
|
DOI |
https://doi.org/10.5281/zenodo.5685489 |
|
persistent identifier |
https://treatment.plazi.org/id/03933D30-6B20-FF95-60B3-9CA61AE1DA46 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus dumnuii |
| status |
sp. nov. |
Cyrtodactylus dumnuii sp. nov.
Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 .
Holotype. THNHM 15904 [formerly Montri Sumontha field series (MS) 302], adult male ( Figure 1 View FIGURE 1 ); Thailand, Chiang Mai Province, Chiang Dao District, Mae-Na Subdistrict, Ban Thakilek, Tham (Cave) Phabartmaejon, 19°20’34” N, 99°01’37”E, 486 m a.s.l., collected by Kirati Kunya, 26 December 2005.
Paratypes. THNHM 15905 [MS 201], adult female, CUMZ R 2009-6-24-5 [MS 202], adult male; same data as holotype. KZM 0 0 2 [MS 181], adult female, CUMZ R 2009-6-24-6 [MS 182], adult female ( Figure 2 View FIGURE 2 ); same locality as holotype, collected by Kirati Kunya, 30 March 2005.
Etymology. We are pleased to name this new species in honor of Mr. Sophon Dumnui, Director of the Zoological Park Organization under the Royal Patronage of His Majesty The King of Thailand. We suggest the following common names: Tuk kai Dumnui ( Thai), Dumnui’s bent toed gecko (English), Cyrtodactyle de Dumnui (French), Dumnuis Bogenfingergecko ( German), Kromvingergekko van Dumnui (Dutch).
Diagnosis. A moderately sized Cyrtodactylus , snout-vent length to at least 84 mm; body slender, limbs and digits long, slender, tail much longer than SVL; one pair of greatly enlarged postmental scales in broad contact with one another; no internasal scales; dorsal scalation with 18–22 regularly arranged rows of keeled tubercles; approximately 40 ventral scales across belly between well-developed, non-denticulate ventrolateral folds; no precloacal groove, 5–6 precloacal pores separated by a diastema of 10–11 poreless scales from a series of 6 small femoral pores borne on a row of enlarged femoral scales in males, femoral pores absent and tiny precloacal pores variably present in females; nine broad basal lamellae and ten narrow distal lamellae beneath 4th toe of pes; median subcaudal scales enlarged to form broad transverse plates; dorsal pattern of alternating light and dark transverse bands on occiput, nape, and body, 6–7 such dark bands between shoulder and sacrum, dark band on occiput usually broken medially.
Description of holotype. Adult male. SVL 81.2 mm; TailL 100.2 mm (91.4 mm regenerated). Head relatively long (HeadL/SVL ratio 0.29), wide (HeadW/HeadL ratio 0.69), not markedly depressed (HeadH/ HeadL ratio 0.44), distinct from slender neck. Loreal region weakly inflated, canthus rostralis not prominent. Snout elongate (SnEye/HeadL ratio 0.41), rounded; longer than eye diameter (OrbD/SnEye ratio 0.63); scales on snout small, rounded to oval, granular to weakly conical, mostly homogeneous, larger than those on crown, interorbital and occipital regions. Eye large (OrbD/HeadL ratio 0.26); pupil vertical with crenelated margins; supraciliaries short, those at posterior of orbit bearing small conical spines. Ear opening vertically elliptical, large (EarL/HeadL ratio 0.09); eye to ear distance greater than diameter of eyes (EyeEar/OrbD ratio 1.10). Rostral much wider (3.46 mm) than deep (2.16 mm), rostral crease very short (<¼ height of rostral). Two enlarged supranasals in broad contact with one another, no internasals. Rostral in contact with first supralabials, nostrils, and supranasals. Nostrils oval, more-or-less laterally directed, each surrounded by supranasal, rostral, first supralabial and two enlarged postnasals. Two to three rows of small scales separate orbit from supralabials. Mental triangular, wider (3.33 mm) than deep (2.82 mm). A single pair of greatly enlarged postmentals in broad contact behind mental, each postmental bordered anteromedially by mental, anterolaterally by first infralabial and anterior portion of second infralabial, posterolaterally by an enlarged lateral chinshield (or second postmental), and posteriorly by three granules, the medialmost of which also contacts the other postmental. Supralabials to midorbital position 9/9; enlarged supralabials to angle of jaws 13/12. Infralabials 10/11. Interorbital scale rows across narrowest point of frontal bone 15.
Body slender, elongate (TrunkL/SVL ratio 0.44) with well defined, non-denticulate ventrolateral folds. Dorsal scales weakly heterogeneous, domed to conical; regularly distributed tubercles (4-5 times size of adjacent scales) extending from shoulder region on to tail base, smaller tubercles on postocular region, crown, occiput, and nape; most tubercles bearing a keel, those on flanks conical, often lacking a distinct keel, those on posterior trunk and sacral region most prominent; tubercles in 22 regular rows at midbody, typically separated from one another by 1–2 dorsal granules, three or more mid-dorsally ( Figure 3 View FIGURE 3 A). Ventral scales larger than dorsals, smooth, oval and subimbricate, largest on posterior abdomen and in precloacal region. Midbody scale rows across belly to lowest rows of tubercles 40. Gular region with homogeneous, smooth, juxtaposed granular scales.
Five precloacal pores in continuous series. No precloacal groove. A continuous row of enlarged femoral scales, each about twice the size of adjacent anterior femoral scales, extending along length of femur and continuous with the precloacal pore-bearing scales. Six (left) to seven (right) tiny femoral pores in distal scales of enlarged femoral scales, separated by 10 (right) to 11 (left) poreless scales from precloacal series ( Figure 3 View FIGURE 3 B). Postcloacal spurs each bearing three enlarged, smooth scales.
Scales on palm and sole smooth, rounded to oval or hexagonal, slightly domed. Scalation on dorsal surfaces of limbs similar to body dorsum with enlarged, conical tubercles interspersed among smaller scales; tubercles separated from one another by 1–2 small scales, or in direct contact with one another. Fore and hindlimbs moderately long, slender (ForeaL/SVL ratio 0.16; CrusL/SVL ratio 0.20). Digits long, slender, inflected at interphalangeal joints, all bearing robust, slightly recurved claws. Basal subdigital lamellae broad, ovoid to rectangular, without scansorial surfaces (5-7-7-6-8 right manus; 6-7-7-9-8 right pes); narrow lamellae distal to digital inflection and not including ventral claw sheath: 9-9-11-10 -8 (right manus), 10-9-11-10-9 (right pes); very weakly developed interdigital webbing between digits (except IV and V). Relative length of digits: IV>III>II>V>I (manus); V>IV~III>II>I (pes). Mostly regenerated tail, long, slightly depressed, gently tapering to pointed tip; longer than SVL (TailL/SVL ratio 1.23). Tail with enlarged median subcaudal plates.
Coloration in ethanol. Dorsum of alternating whitish to grayish-brown and chocolate brown transverse bands; chocolate bands with paler centers, grayish-brown bands often with darker centers. Six dark bands from shoulder to sacrum, another on tail base. Additional incomplete dark bands on nape and occiput. Dark bands fusing near ventrolateral margin of trunk. Most dorsal tubercles, except those on darkest brown areas, whitish. Dorsum of head medium brown with irregular cream to grayish markings between occipital band and posterior margin of orbit. A chocolate brown stripe extends from lower posterior corner of orbit, forms a right angle above the tympanum, and passes medially to form the broken occipital band. A vague pale canthal stripe is present. Supra- and infralabials mottled brown and gray, darkest anteriorly. Dorsal surfaces of limbs with alternating chocolate and gray-brown markings, palms grayish. Dorsum of regenerated tail light brown with irregular beige to cream markings. Dorsal trunk coloration ends abruptly at ventrolateral folds. Venter beige with brown speckling, densest under chin, on lower abdomen and precloacal region, under thighs, and at lateral margins. Tail venter mottled gray-brown.
Color in life (based on photographs of uncatalogued specimens, Figure 4 View FIGURE 4 ) similar to that in preservative. Iris green, rims of orbits and posterior supralabials pale yellowish to white.
Variation. Comparative mensural data for the holotype and paratypes are given in Table 1 View TABLE 1 . Meristic characters of the paratypes mostly correspond to those of the holotype except as noted. Rows of tubercles 18– 20 in paratypes (Tabler 1). Male paratype with six precloacal pores separated by 11 poreless scales from a series of six femoral pores on each thigh. Female paratypes with no femoral pores and 0–7 precloacal pores ( Table 1 View TABLE 1 ); if present, precloacal pores minute. Original portion of tail of CUMZ R 2009-6-24-6 weakly differentiated into annuli, with two subcaudal plates corresponding to eight rows of small, rectangular, subimbricate dorsal scales, decreasing to seven, then six such rows posteriorly. Dorsally original tail with cream blotches separated by more extensive brown markings. THNHM 15905 with seven dark transverse bands between shoulders and sacrum. CUMZ R 2009-6-24-6 with occipital band complete, partly complete in CUMZ R 2009-6-24-5. In living non-type specimens ( Figure 4 View FIGURE 4 ) the slender original tail exhibits 13–14 ashy blotches or rings on a chocolate to purplish-brown background. Caudal tubercles are limited to the pygal portion of the tail only.
Comparisons with other species. As discussed by Bauer et al. (2009), a number of geographically coherent and/or phenotypically similar clusters of species of Cyrtodactylus have recently been recognized in the literature. However, a comprehensive phylogenetic frame work for the group is still pending. We therefore provide comparisons of C. dumnuii sp. nov. to all of the Cyrtodactylus species described as of 1 April 2010 (exclusive of the distinctive South Asian Geckoella , sometimes considered as a subgenus of Cyrtodactylus , and a number of bent-toed geckos of questionable generic allocation occurring in Nepal, northern India, and Tibet [Xizang Autonomous Region]).
Rösler and Glaw (2008) provided an extensive table summarizing a diversity of features across nearly 100 species of Cyrtodactylus sensu lato. Because of the incredible rate at which new Cyrtodactylus have been described from southeast Asia, this set of comparisons is already somewhat outdated. Data from additional taxa and corrections to Rösler and Glaw’s (2008) data were derived from Oliver et al. (2008), Grismer & Norhayati (2008), Grismer et al. (2008), Linkem et al. (2008), Nazarov et al. (2008), Ngo (2008), Ngo & Bauer (2008), Ngo et al. (2008), Rösler et al. (2008), Bauer et al. (2009), Geissler et al. (2009), Welton et al. (2009, 2010), Chan & Norhayati (2010), Sumontha et al. (2010), and Ziegler et al. (2010).
The presence in males of separate series of femoral pores (versus no femoral pores or continuous precloacal-femoral pores) in C. dumnuii distinguishes it from C. angularis (Smith) , C. chanhomeae Bauer et al. , C. intermedius (Smith) , C. jarujini Ulber , C. oldhami (Theobald) , C. papilionoides Ulber & Grossmann , C. peguensis (Boulenger) , C. quadrivirgatus Taylor , C. sumonthai Bauer et al. , C. thirakhupti Pauwels et al. , and C. variegatus (Blyth) , among Thai congeners and from the following extralimital species: C. adleri Das , C. annulatus (Taylor) , C. aurensis Grismer , C. ayeyarwadyensis Bauer ; C. badenensis Nguyen et al. , C. batucolus Grismer et al. , C. brevidactylus Bauer , C. cattienensis Geissler et al. , C. cavernicolus Inger & King , C. chauquangensis Hoang et al. , C. condorensis (Smith) , C. consobrinoides (Annandale) , C. cracens Batuwita & Bahir ; C. cryptus Heidrich et al. , C. deveti (Brongersma) , C. edwardtaylori Batuwita & Bahir ; C. elok Dring , C. epiroticus Kraus , C. feae (Boulenger) , C. fraenatus (Günther) , C. fumosus (Müller) , C. gansi Bauer , C. halmahericus (Mertens) , C. hontreensis Ngo et al. , C. ingeri Hikida , C. irianjayaensis Rösler , C. irregularis (Smith) , C. khasiensis (Jerdon) , C. klugei Kraus , C. lateralis (Werner) , C. malayanus (de Rooij), C. malcolmsmithi (Constable) , C. matsuii Hikida , C. nigriocularis Nguyen et al. , C. nuaulu Oliver et al. , C. pantiensis Grismer et al. , C. papuensis (Brongersma) , C. paradoxus (Darevsky & Szczerbak) , C. phongnhakebangensis Ziegler et al. , C. pseudoquadrivirgatus Rösler et al. , C. pubisulcus Inger , C. ramboda Batuwita & Bahir , C. robustus Kraus , C. roesleri Ziegler et al. , C. rubidus (Blyth) , C. seribuatensis Youmans & Grismer ; C. sermowaiensis (de Rooij), C. serratus Kraus , C. soba Batuwita & Bahir ; C. stresemanni Rösler & Glaw , C. subsolanus Batuwita & Bahir ; C. sworderi (Smith) , C. tautbatorum Welton et al. , C. tiomanensis Das & Lim , C. tuberculatus (Lucas & Frost) , and C. yoshii Hikida.
The presence of precloacal pores in males distinguishes C. dumnuii from C. eisenmanae Ngo ; C. gordongekkoi (Das) , C. jellesmae (Boulenger) , C. laevigatus Darevsky , C. semenanjungensis Grismer & Leong , and C. wallacei Hayden et al. It differs from its Thai congeners C. consobrinus (Peters) , C. interdigitalis Ulber and C. tigroides Bauer et al. , as well as C. aaroni Günther & Rösler , C. aequalis Bauer , C. baluensis (Mocquard) , C. caovansungi Orlov et al. , C. louisiadensis (de Vis), C. mimikanus (Boulenger) , C. russelli Bauer , C. salomonensis Rösler et al. , C. slowinskii Bauer , and C. tripartitus Kraus in a lower number of precloacal pores (5–6 versus 7 or more [usually 9 or more]).
Cyrtodactylus dumnuii may be distinguished from C. macrotuberculatus Grismer & Norhayati , C. philippinicus (Steindachner) , C. spinosus Linkem et al. , and the Thai species C. pulchellus Gray , by the absence of a longitudinal or steeply angled precloacal sulcus or groove, from C. marmoratus (Kuhl) and C. sadleiri Wells & Wellington by the absence of a recessed transverse precloacal groove, from C. biordinis Brown & McCoy by the presence of a single, versus double row of femoral pores, and from C. chrysopylos Bauer by the absence of an enlarged pore-bearing scale posterior to the precloacal pore row. An enlarged series of femoral scales separates the new species from C. annandalei Bauer , C. buchardi David et al. , C. grismeri Ngo , C. murua Kraus & Allison , and C. wakeorum Bauer.
The new species may be distinguished from the Thai species C. brevipalmatus (Smith) and the extralimital congeners C. agamensis (Bleeker) , C. agusanensis (Taylor) , C. capreoloides Rösler et al. , C. darmandvillei (Weber) , C. derongo Brown & Parker , C. gubernatoris (Annandale) , C. huynhi Ngo & Bauer , C. jarakensis Grismer et al. , C. loriae (Boulenger) , C. novaeguineae (Schlegel) , C. papilionoides Ulber & Grossmann , C. wetariensis (Dunn) , C. ziegleri Nazarov et al. , and C. zugi Oliver et al. by the presence of enlarged subcaudal plates in the original tail and from C. redimiculus King and C. takouensis Ngo & Bauer by its greater number of rows of dorsal tubercles (18–22 versus 14–16 and 10, respectively). Finally, C. dumnuii also differs from its most recently described Thai congeners; from C. erythrops Bauer et al. it is distinguished by its greater number of scales across the venter at midbody (40 versus 28), and from C. auribalteatus Sumontha et al. it differs in having 6–7 (versus 4) dark bands from the shoulders to the sacrum.
Distribution and natural history. Cyrtodactylus dumnuii has been collected only from a single locality near the Myanmar border in Chiang Mai Province in the northwest of Thailand ( Figure 5 View FIGURE 5 ). Specimens were collected inside the entrance and up to 30 m within Tham (Cave) Phabartmaejon ( Figure 6 View FIGURE 6 ), during daylight hours.


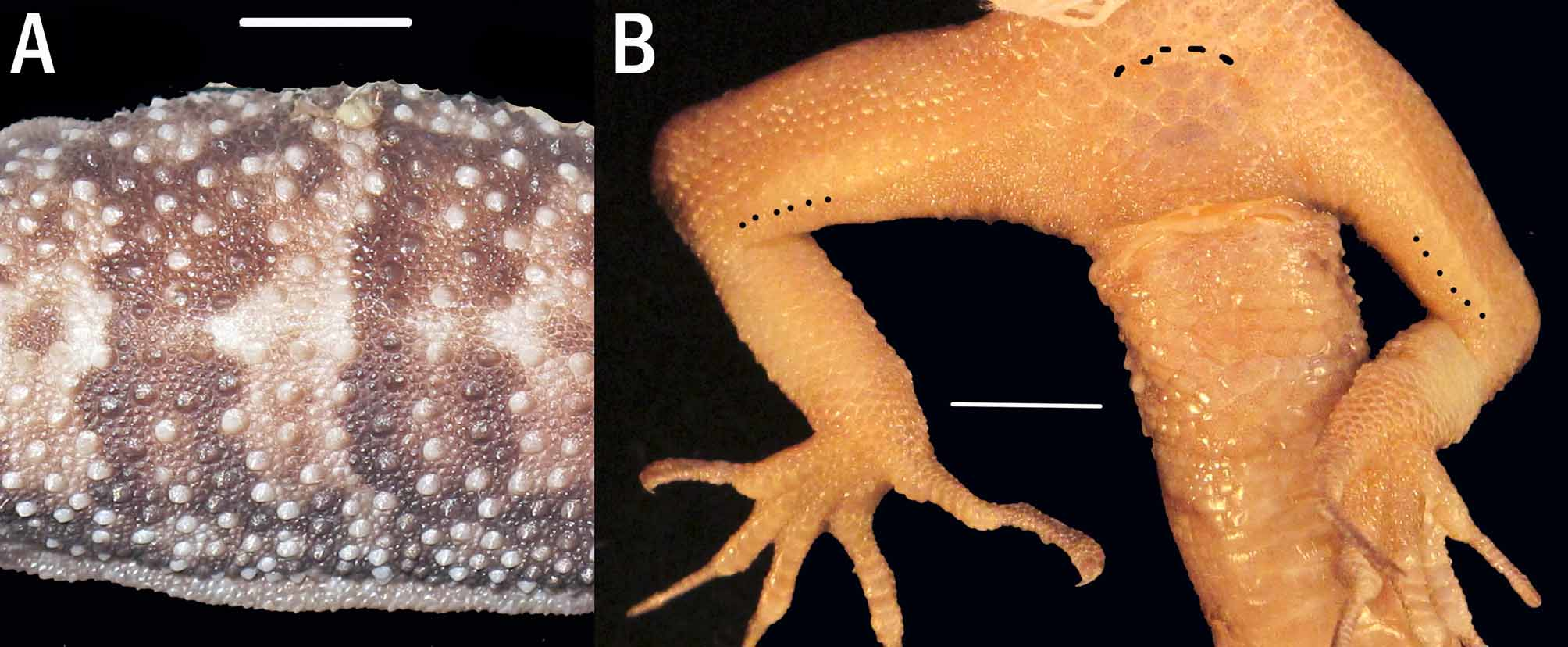



TABLE 1. Mensural and selected meristic data for the type series of Cyrtodactylus dumnuii sp. nov. DorTub = Longitudinal Rows of Dorsal Tubercles, PreclP = Precloacal Pores, FemP = Femoral Pores (Left – Right), other abbreviations as in Materials and methods. All measurements in mm. * = head damaged, reliable measurements not possible.
| THNHM 15904 Holotype | CUMZ R 2009-6-24-5 Paratype | KZM 0 0 2 Paratype | CUMZ R 2009-6-24-6 Paratype | THNHM 15905 Paratype | |
|---|---|---|---|---|---|
| Sex | male | male | female | female | female |
| SVL | 81.2 | 84.2 | 83.6 | 76.2 | 76.5 |
| ForeaL | 12.8 | 14.5 | 13.8 | 13.6 | 12.9 |
| CrusL | 15.9 | 17.2 | 16.3 | 15.3 | 15.4 |
| TailL | 100.2 (8.8 original) | 54.0 (17.4 original) | 95.1 (9.1 original) | 33.6 (broken) | 7.1 |
| TailW | 7.8 | 7.5 | 6.7 | 4.4 | — |
| TrunkL | 35.6 | 36.1 | 35.4 | 34.7 | 33.3 |
| HeadL | 23.4 | 23.4 | 23.1 | 22.8 | 23.3 |
| HeadW | 16.1 | 16.4 | 16.0 | 14.8 | 14.6 |
| HeadH | 10.4 | * | 10.0 | 10.2 | 8.5 |
| OrbD | 6.0 | 5.2 | 5.7 | 6.1 | 6.0 |
| EyeEar | 6.6 | 6.9 | 6.6 | 6.2 | 5.7 |
| SnEye | 9.7 | 10.2 | 9.7 | 9.4 | 9.1 |
| NarEye | 7.0 | 7.2 | 6.8 | 7.0 | 6.5 |
| Interorb | 3.6/7.5 | * | 3.7/7.7 | 3.3/7.6 | 3.4/7.2 |
| EarL | 2.1 | 2.2 | 2.7 | 2.4 | 2.7 |
| Internar | 2.6 | 2.7 | 2.7 | 2.5 | 2.6 |
| DorTub | 22 | 20 | 18 | 18 | 20 |
| PreclP | 5 | 6 | 5 | 0 | 7 |
| FemP | 6–7 | 6–6 | 0 | 0 | 0 |
| CUMZ |
Chulalongkorn University Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.