
Gryllus vocalis Scudder
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4705.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:F534C43A-AB09-4CB3-9B08-FD5BDFD90298 |
|
persistent identifier |
https://treatment.plazi.org/id/182387A8-09F9-FF32-51F6-F904027CF80D |
|
treatment provided by |
Plazi |
|
scientific name |
Gryllus vocalis Scudder |
| status |
|
Damp-Loving Field Cricket
Figs 155-163 View FIGURE 155 View FIGURE 156 View FIGURE 157 View FIGURE 158 View FIGURE 159 View FIGURE 160 View FIGURE 161 View FIGURE 162 View FIGURE 163 , Table 1 View TABLE 1
1901 Gryllus vocalis Scudder, Psyche 9: 268. Lectotype male ( Fig. 158 View FIGURE 158 ), courtesy of J. Weintraub) designated by Weissman et al. (1980): “ L. Angeles, Calif., July 29, 1897. Gr. vocalis , Scudder’s type 1901. Red type label, type 14070.” Deposited in ANSP.
1902 Gryllus alogus Rehn. Proc. Acad. Nat. Sci. Philadelphia 54: 726. Holotype female: “Albuquerque, 1902. N. M. T.D.A. Cockerell/Red type label Gryllus alogus Rehn Type No. 5067.” Adult type ( Fig. 159 View FIGURE 159 , courtesy of J. Weintraub) with black head and pronotum, pronotum hirsute, tegmina tan, hind wings short, all legs orange brown. Head narrower than pronotum. Some brown-red markings in area of lower face. Body 17.2, hind femur 10.9, ovipositor 14.8, head width 5.2, pronotum width 5.7, pronotum length 3.4.
Because the holotype is a female without intact cerci, we don’t have the benefit of male file characters or cerci length to help in determining which species this is. The following five Gryllus species occur either in the city of Albuquerque or its vicinity (we considered a geographical area wider than Albuquerque since many old collecting localities were frequently “assigned” to the nearest town): G. armatus , G. longicercus , G. veletis , G. lightfooti , and G. alogus . We eliminate G. armatus because the body colors of the holotype of G. alogus are different, her pronotum is too long and hirsute, and she has short hind wings. We eliminate G. longicercus because the ovipositor to hind femur ratio in New Mexico specimens ( Fig. 160 View FIGURE 160 ) is wrong, the pronotum is hirsute, and the head is narrower than the pronotum, the last two qualities not seen in G. longicercus . We eliminate both G. veletis and G. lightfooti because their pronotums are not hirsute or minimally so, and the body, tegmina, cerci and legs of the latter two taxa are generally black and not orange brown, as in the holotype. This elimination process leaves only G. alogus . Deposited in ANSP. New synonymy.
1981 ‘ Gryllus IV’, Rentz & Weissman (1981).
‘ Gryllus #10, #18’, ‘arizonensis’ and alogus of DBW notebooks.
‘Regular stutter-triller’ of Sakaguchi & Gray (2011).
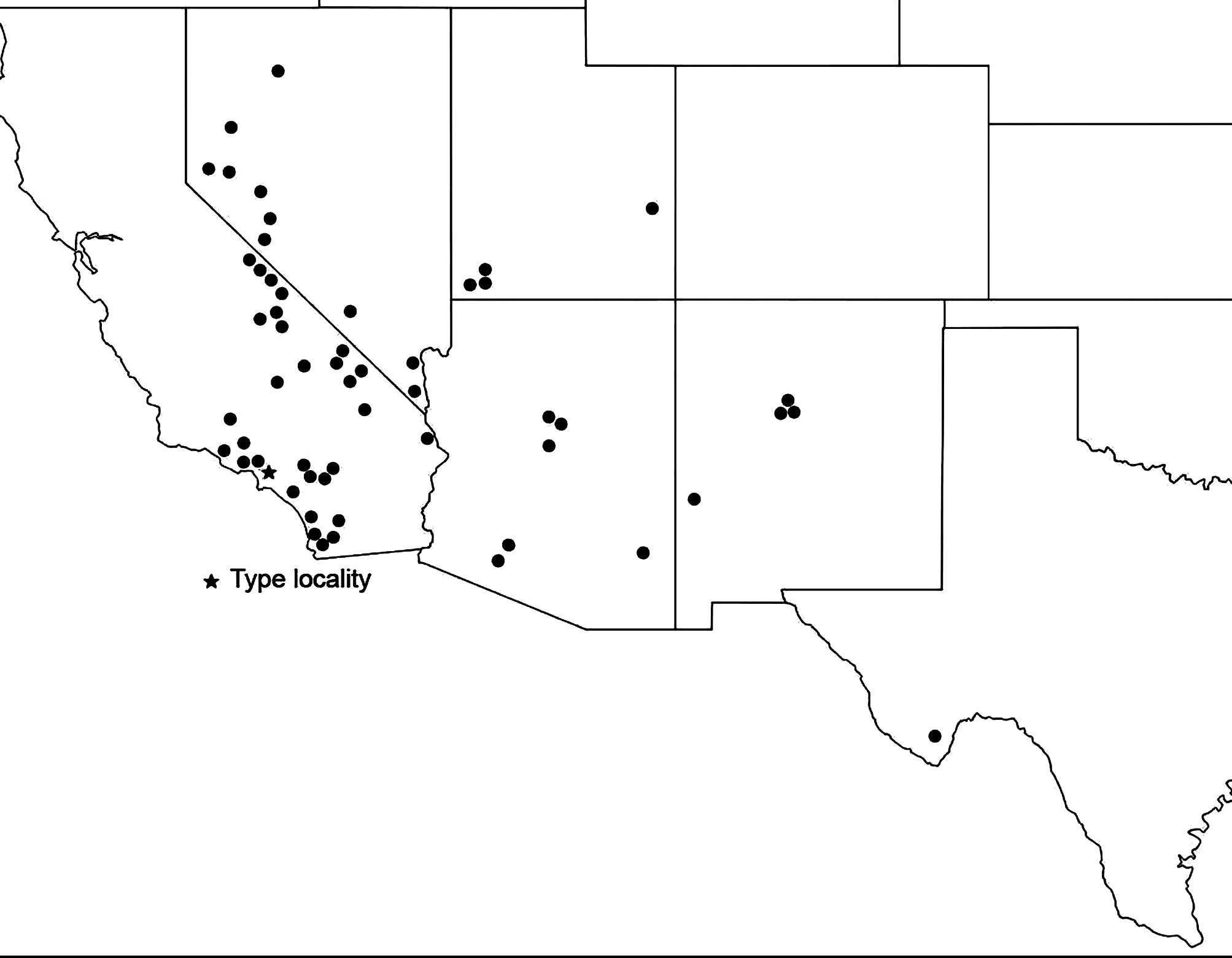
Distribution. Widespread ( Fig. 163 View FIGURE 163 ) across the Southwestern US from southern California up along the eastern side of the Sierras, through southwestern Nevada, southern Utah, Arizona, New Mexico, and east to Big Bend, Texas.
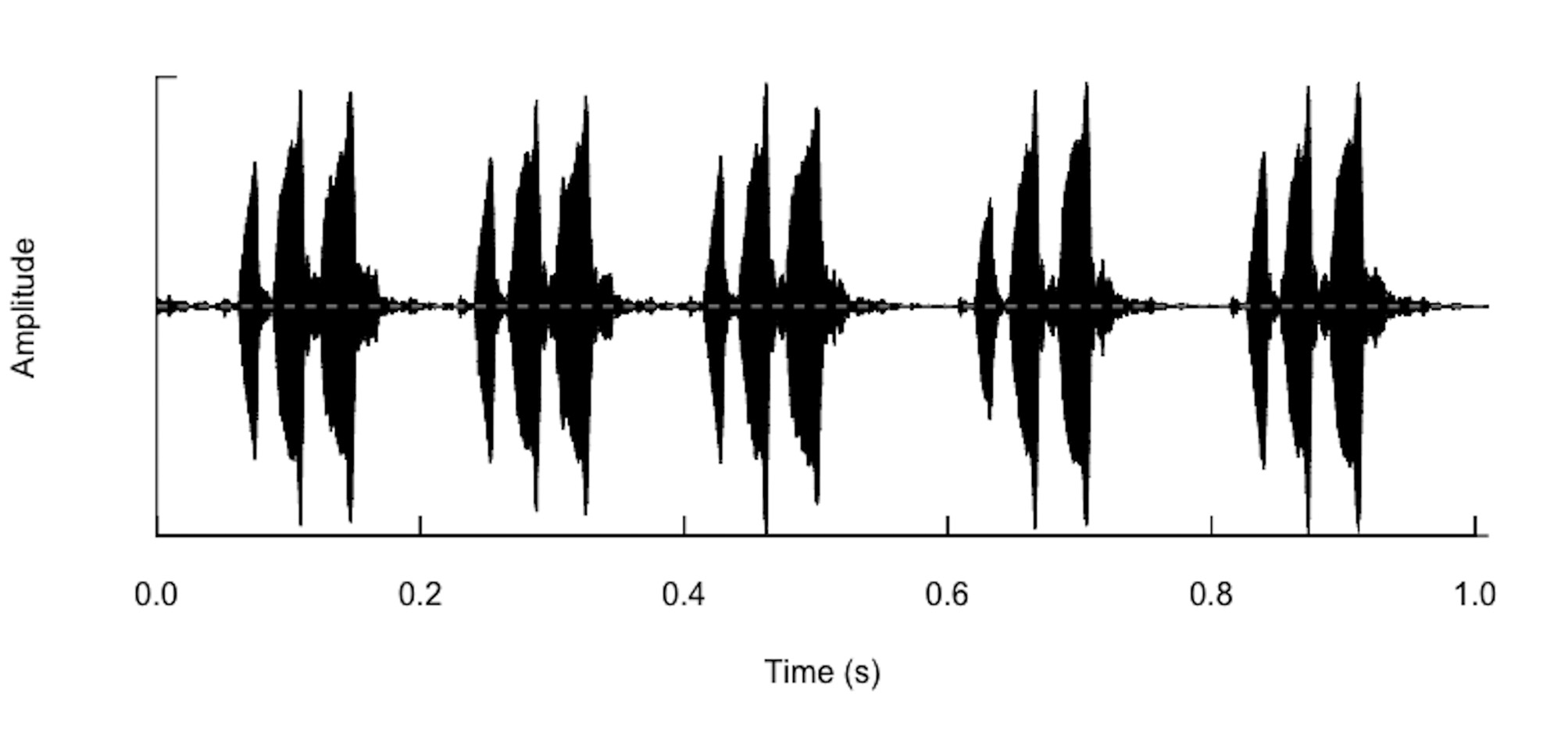
Recognition characters and song. While geographically widespread, G. vocalis has a narrow microhabitat and is almost always associated with wet, verdant habitats such as coastal salt marshes (Cardiff by the Sea, S80-48, 49), along seasonal (Whitewater, S09-21 and Agua Fria, S13-14) and year-round streams (Furnace Creek, Death Valley, S80-32, S83-60, S03-36; and Zion National Park, S01-33), in seasonal ponds with Distichlis salt grass (7.2 km N Beatty, NV, S80-34), and in gardens ( Gila Bend, S 15-111) and artificially watered areas. Medium to large crickets that almost invariably have a dark head, pronotum and tegmina ( Fig. 161 View FIGURE 161 ) from coastal California east to the borders of the Mohave Desert, while those from Owens Valley (Big Pine and Bishop) are also dark. East of the Mohave Desert, individuals in most populations usually with dark heads and pronotum but light colored tegmina. Song ( Fig. 162 View FIGURE 162 , R09-17) unique within the western US: 3 p/c (range 2-4), PR 23-45 at 25°C, 5-10 c/s evenly delivered. Their song can only be confused with certain western US individuals of G. cohni from central-southern Arizona and we extensively discuss how to separate the 2 taxa on p. 162. Males singing at 30+°C in Gila Bend , sounded more like G. armatus than G. vocalis because of their increased chirp rate and shortened pulse period when that warm. G. vocalis and G. cohni are apparently only microsympatric at Ajo, Arizona (S98-72). Song of 3p/c also similar to more eastern G. fultoni and G. vernalis but CR faster in G. vocalis and nowhere sympatric. Distinguished from allopatric G. veintinueve by faster chirp rate and almost non-overlapping and faster pulse rate in G. vocalis , in addition to different DNA.
Derivation of name. “voco” is Latin for call, perhaps referring to the distinctive fast chirp rate and loud calling song of this species. Which raises the interesting question of who collected the lectotype and did this person subsequently relate such information to Scudder? In 1896, Scudder developed Parkinson’s disease (aka paralysis agitans) and started to prepare for his decline and demise ( Mayor 1919), such that we assume that he did not visit Los Angeles in July, 1897 when the lectotype was collected. Scudder retired in 1902 and died in 1911. No further collection information is available at ANSP (J. Weintraub, pers. comm. to DBW, June, 2018).
Geographic range. Fig. 163 View FIGURE 163 . Also into northern Baja California Norte, Mexico.
Habitat. Individuals with an impressive temperature tolerance and almost always associated with wet habitats: Salt marshes (Cardiff by the Sea, S80-48, 49) and freshwater marshes (Shoshone, S85-27), irrigated gardens, cultivated ivy, along desert streams and ponds and, rarely, in railroad track rocks (near UC Riverside, S03-40). In San Diego (S97-60), 1 male singing 3m above ground in hole in tree with a long hind winged female near him. In a nearby palm tree, another male singing from 1.5m above ground. At coastal Ballona Wetlands (Los Angeles Co., S92-58) in ice plant on sandy substrate. While easy to approach, singing males can be very difficult to collect because they sing from dense vegetation. Oatmeal trails and checking under available rocks and trash can assist in such situations.
Life cycle and seasonal occurrence. No egg diapause: Arizona: S15-111 ( Gila Bend ). California: S92-60 ( Shoshone ), S95-112 (Big Pine), S03-73 (Barstow). Nevada: S90-44 (Cottonwood Cove). Apparently one or two generations/year depending on locality. In coastal California, a definite late winter—early spring species probably with one generation/year as those March , Orange Co. specimens (S05-28 and S05-29) were collected as late instars and no adults were heard after June at these localities .
S. Gershman (pers. comm. to DAG, 18-xi-2014) reports for inland and warmer Riverside, California: “I found a few calling males (of G. vocalis at the UCR Botanic Garden—S03-39) even in the winter. Here’s what I think is happening. Eggs do not diapause under summer conditions in the lab. Eggs are fairly constant in development time. Adults do not differ much in longevity. But, even in the lab under constant light and temperature, there is a huge range in development times of nymphs. Under lab summer conditions, some G. vocalis go from hatching to adult in 2 months. Others take more than 6 months. I assume that this spread causes individuals in nature to emerge as adults at random times throughout the year.”
Nevertheless, we have the following late summer collections in southern California: 3 adult males on 7-ix-2001 at Sepulveda Basin Wildlife Refuge in Los Angeles Co.; 1 adult male 11-ix- 1977 in Orange in Orange Co.; and 2 adult males on 8-ix-2005 at Afton Canyon in San Bernardino Co. We wonder if certain California localities might have sporadic second-generation adults, as discussed for G. veletisoides (see p. 195), which make no genetic contribution to future generations.
In contrast, interior western US G. vocalis are often abundant in late summer and fall, e.g. at Zion National Park, Utah, 5-7-ix-2008, 5♂ 11♀. DAG recorded the following data for near Los Lunas, NM (S94-44 and S94-100), in 1994, which suggests 2 generations/year: Collecting in the last 2 weeks of June and the first 2 weeks of July showed adults abundant with some 80 collected. On July 25 th , only one adult female seen; on August 8 th, 8 adults; August 10 th, 12 adults; August 23 rd, 20 adults; August 24 th, 35 adults; September 6 th, 36 adults. In central Arizona, DAG has also observed both spring and fall adults: Agua Fria National Monument, Bloody Basin Road at Agua Fria river , Yavapai Co ., 2 males 27-iii-2005, 2 females 1 nymph 16-iv-2010; Wet Beaver Creek Campground, Yavapai Co ., 3 males 9 females 8-viii-2007; Oak Creek Canyon, Manzanita Campground, Coconino Co ., 2 males 18-viii-2004.
Variation. Pronotal hirsuteness: variable resulting in shiny to dull surfaces within many populations. Color: varies as discussed under “Recognition characters.” Of 10 males and 3 females from Death Valley (S80-32 & S83- 60), all 10 males and 1 female had light colored tegmina with male pronotums ranging from reddish to solid black. In a large 1994 survey at Las Lunas, NM, DAG found most individuals with dark brown pronotums, although some definitely reddish. Hind wing length: All 80 first generation field (i.e. those collected before 8-viii-1994), Las Lunas, NM, individuals, with short hind wings. Second generation field individuals collected there after 8-viii-1994, showed 26 of 58 males and 22 of 53 females with long hind wings. All 7 individuals from Cottonwood Cove (S90- 44) had long hind wings. Of 33 males and 19 females from the type locality of G. alogus (Albuquerque, NM), all but 2 females had short hind wings. All 16 males and 49 females from along permanent stream at Agua Fria (S13-14) had short hind wings. All 8 males and 9 females from the watered garden area at McDonald’s in Gila Bend , Arizona, had long hind wings. One female was dealate. Song: males in some populations have long series of chirps (Albuquerque, S85-52) while others ( Gila Bend, S 15-111) have more broken up songs. Tegmina length: variable within and between populations.
Specimens examined. (Total: 333♂ 216♀). Arizona: Coconino Co., Sedona, 4500’, 15-vi-1990 (S90-49) 1♂; 5-viii-1991 (S91-78) 5♂; 30-vi-1994 (S94-35) 1♂. Graham Co., 4.5 m S Safford , 3180’, 10-vi-2012 (S12-20) 1♂ . Maricopa Co., Buckeye , 840’, 18-ix-2011 (S11-102) 1♂. Gila Bend , garden area at McDonald’s , 32° 56’ 38.5” - 112° 43’ 57.5”, 712’, 1-viii-2009 (S09-103) 3♂ ; 18-ix-2011 (S11-101) 1♂ ; 30-vii-2015 (S15-111) 4♂ 10♀. Hwy 85 E Goodyear, 980’, 31-vii-1981 (S81-46) 3♂ . Yavapai Co., Agua Fria National Monument , stream crossing 4.9 m SE of pavement end, 989m, 34° 15’ 27.57” -112° 03’ 50.5”, 27-iii-2005, 2♂ ; 16-iv-2010, 2♂ ; 12-vi-2012 (S12-26) 1♀; 31-v-2013 (S13-14) 16♂ 49♀. Wet Beaver Creek Campground, 8-viii-2007, 3♂ 9♀ . Hwy 179 eastern side Sedona, 4000’, 31-v-2013 (S13-16) 2♂ . California : Inyo Co., Big Pine , 10-ix-1995, 4000’ (S95-112) 1♂ 1♀ . Bishop , 5-vi- 1983, 4450’ (S83-56) 1♂ ; 28-viii-2005 (S05-99) 3♂. 9 m N Bishop on Hwy 395, 1430m, 4-vi-1983, (S83-55) 13♂ 1♀ ; 26-vi-1992 (S92-64) 4♂ 5♀ . Death Valley National Park , Furnace Creek, -190’, 23-vi-1980 (S80-32) 5♂ 3♀ ; 5-vi-1983 (S83-60) 5♂ ; 5-v-2003 (S03-36) 1♀. Little Lake 20 m S Olancha, 3400’, 5-v-2003 (S03-33) 8♂ 7♀ . Lone Pine , 3650’, 5-viii-1978 (S78-117) 1♂ , 29-viii-2005 (S05-105) 1♂ . Shoshone , 12-iii-1985 (S85-27) 2♂ 6♀ ; 25-vi- 1992 (S92-60) 10♂ 10♀ ; 5-v-2003 (S03-37) 3♂ 3♀. Hwy 127 0.6 m N Shoshone , 1600’, 6-vi-1983 (S83-61) 13♂ 6♀ . 3 m W Lone Pine on Whitney Portal Rd , 5-viii-1978 (S78-119) 1♂ . Kern Co., Frazier Park , 16-vi-1978, 4600’, 1♂ . Kern River area across from CSU Bakersfield, 5-viii-1980 (S80-70) 1♂ . Mojave , 2757’, 5-viii-1988 (S88-70) 2♂ . Los Angeles Co., Ballona Wetlands , 24-vi-1992 (S92-58) 6♂ 2♀ . CSU Northridge , 810’, 8-v-2003, (S03-47) 3♂ ; early April, 2004 (S04-27) 2♂ 1♀. Malibu near Big Rock and Rockport Rds., 26-vi-1976, 2♂ 1♀ ; Malibu Canyon Rd. near Pacific Coast Hwy, 1-vii-1976, 1♂ . Santa Monica Mts. , Cold Creek, 2-v-2002, 3♂ 2♀ . Sepulveda Basin Wildlife Refuge , 7-ix-2001, 3♂ . Sherman Oaks , 28-vi-1976, 2♂ . Mono Co., 9 m N Laws at western base of White Mts., 26-vi-1992, 4500’ (S92-63) 5♂ 4♀ . Orange Co., Costa Mesa , Talbert Nature Preserve, 15-iii-2005, 20’ (S05-28) 1♂ . Irvine near UCI Medical Center, 6-iv-1978, 2♂ 2♀ . Newport Beach , Backbay Rd., 7-vii-1976, 5♂ 6♀ . Orange , 9-iv-1978, 7♂ 1♀ ; 11-ix-1977, 1♂ . Peters Canyon Regional Park , 16-iii-2005, 620’ (S05-29) 1♂ . Riverside Co., Riverside , mid-July, 2001 (S01-96) 2♂ 1♀ . UC Riverside Botanic Garden , 6-v-2003 (S03-39) 6♂ . Near UC Riverside, railroad tracks, 6-v-2003 (S03-40) 2♂ . Whitewater Canyon ca. 2.5 m N I10, 1-iv-1978, 1300’ 2♂ 2♀; 26-v-2009 (S09-21) 1♂. San Bernardino Co., Afton Canyon , Mojave River, 1400’, 35.03920° -116.38260°, 2-v-2004, 4♂ 4♀ ; 1-v-2005 (S05-50) 7♂ 4♀ ; 8-ix-2005, 2♂. Barstow , 2300’, 16-viii-1998 (S98-58) 1♂ ; 28-vi-2003 (S03-73) 1♀ . Cherry Valley , 4-iv-1991, 4000’ (S91-12) 1♂ 1♀ . Rancho Cucamonga , 10-iii-1998 (S98-9B) 1♂ . San Bernardino Mts. , Mill Creek Ranger Station, 25-vii-1981 2800’ (S81-27) 1♂ . San Diego Co., Camp Pendleton , 10- vii-1976, 4♂ 1♀ . Cardiff by the Sea, Manchester Ave., 27 & 28-vi-1980 (S80-48, 49) 28♂ 15♀ ; 7-v-2003 (S03-42) 3♂ . San Diego , Mission Bay Hilton, 18-vi-1994 (S94-34) 3♂ 1♀ ; 5-vi-1997 (S97-60) 5♂ 1♀ . Hwy 79 0.8 m S Warner Springs , 26-v-2009, 3000’ (S09-18) 2♂ . Hwy 79 23.8 m E 15, 17-iii-2005, 2820’ (S05-32) 1♂ 1♀. Hwy S 22 2.1 m W Anza Borrego , 4000’, 17-iii-2005 (S05-34) 2♀ . Ventura Co., Ojai Valley Swim and Spa Club, 3-iv-1999, 1000’ (S99-5) 2♂ . Nevada : Churchill Co., Fallon , Churchill Community Hospital, 4000’, 16-ix-1998 (S98-95) 5♂ 1♀ . Hwy 95 2.5 m N Fallon , 3900’, 21-vi-2005 (S05-69) 1♂ 1♀ . Hwy 50 Alt. 12.8 m NW Fallon at Bench Rd intersection, 19-iii-1993, 3900’ (S93-7) 2♂ raised from 9 collected mid-instars. Clark Co., Cottonwood Cove , 800’, 14-vi-1990 (S90-44) 3♂ 4♀ . Echo Bay , Lake Mead, 1600’, 7-vi-1989 (S89-33) 2♂ . Lyon Co., 0.5 m N Fernley on Hwy 50, 4-vi-1983, 1311 m (S83-51) 1♂ . Mineral Co., Hwy 95 near Mina, 4600’, 3-vii-2005 (S05-68) 2♂ . Nye Co., Hwy 95 0.5 m N Beatty, 20-vi-2005 ( DL05-16 ) 1♂ 1♀ . Hwy 95 4.5 m N Beatty , 3500’, 24-vi-1980 (S80-34) 1♂ . New Mexico: Bernalillo Co., Albuquerque , 5300’, late May, 1984 (S84-25) 6♂ ; 6-vi-1985 (S85-52) 5♂ 6♀; 18-vii-1985 (S85-95) 4♂; 13-vi-1986 (S86-34) 12♂ 9♀; 23-vi-1993 (S93-59) 7♂ 5♀. Rio Grande Nature Center , June, 2005, D.C. Lightfoot, 1♂ ; 11-vii-2012, DC Lightfoot, 5♂. Catron Co., Village of Reserve , 5770’, 3-vii-1994 (S94-50) 4♂ 2♀ . Sierra Co., Caballo Reservoir State Park , 7-vi-1986 (S86-38) 3♂ 1♀ . Valencia Co., Los Lunas , 1 m S Los Lunas Bridge, 1-vii-1994 (94-44) 2♀ ; early September, 1994 (S94-100) 6♂ 5♀. Texas : Brewster Co., Big Bend National Park , Rio Grande Village, 2100’, 9-vi-1985 (S85-56) 5♂ 2♀ ; 5-vi-1991 (S91-43) 3♂ 2♀ . Utah : Grand Co., Moab, 25-iv-2006 (S06-21) 4♂ . San Juan Co., 12 m W Farmington , 19-vi-1987 (S87-80) 1♂ . Rest sta- tion 12 m S Moab , 21-vi-1987 (S87-59) 4♂ 4♀ . Washington Co., Hurricane , 3420’, 20-iv-1999 (S99-12) 1♂ 3♀ . La Verkin , 3420’, 11-ix-2004 (S04-121) 1♂ . Springdale , 4000’, 9-viii-1991 (S91-94) 2♂ . St. George , 3040’, 19-v-1995 (S95-39) 6♂ 1♀ . Zion National Park , near turnoff to Zion Narrows, 4200’, 19-v-1995 (S95-38) 1♂ ; Zion Museum , 3980’, 19-v-2001 (S01-33) 2♀ . 1 m W entrance to Zion National Park , 4060’, 10-vi-1996 (S96-56) 1♀ .
Uncertain placement. Arizona: Pima Co., Ajo , plaza area, 520m, 20-viii-1998 (S98-72) 1 male (R98-28). This long hind winged male has 3p/c delivered evenly and probably represents G. vocalis . If true, this site is the only documented locality where G. cohni and G. vocalis occur microsympatrically. Hwy 85 13 m N Ajo, 1255’, 30-vii- 2015 (S15-110) 1 female. This female has ITS2 DNA that agrees with G. vocalis despite the absence of water. No males heard singing in area.
Song records only. Nevada: Pershing Co., Rye Patch State Recreational Area , 4050’, 27-vi-1992 (S92-67) 3 males heard. California: Ventura Co., McGrath State Beach , 24-vi-2007 .
DNA. ITS2 and multilocus 2016-036, Los Angeles Co., type locality of G. vocalis ; G3335, Albuquerque, NM, type locality of G. alogus ; and G3227 from Gila Bend , AZ (S15-111), locality of ‘G. arizonensis’ all map together (Gray et al. 2019) and are all sister species to multilocus G. cohni G101, Baja California Sur, type locality (S95-81); G2776 Mazatlán, Mexico (S14-53); and 2016-041 Agua Fria National Monument. We caution that G. cohni type locality leg G101, used in both 16S and multilocus sequencing, was removed, in 2003, from a pinned specimen, eight years after collection. In both sequences, the leg mapped consistent with other G. cohni and near sister species G. vocalis specimens. A singleton male from 7.2 km S Safford (S12-20, G2244) had a unique 16S haplotype but mapped with all other G. vocalis for ITS2 gene.
Discussion. We initially divided G. vocalis into four groups mainly separated by geographic regions. We discuss our early thinking here so that future researchers can revisit these results and reexamine some of our uncertainties. G. vocalis (with a type locality of Los Angeles, California) was defined by Weissman et al. (1980) as those dark 3p/c crickets in southern coastal California that extended through the Mohave Desert and into the Owens Valley east of the Sierras. G. alogus (aka ‘G. #18’) (with a type locality of Albuquerque, New Mexico) is a light colored 3p/c cricket of the Rocky Mountains that extends southeast into Big Bend, Texas. ‘G. #10’ is the light colored 3p/c cricket that occurs in the northern Mojave Desert, Great Basin Desert and Arizona between G. vocalis and G. alogus . ‘G. arizonensis’ was restricted to Gila Bend , AZ, where all individuals had long hind wings. While 1 out of 7 Gila Bend males had a broken song, although still with 3p/c, his ITS2 DNA, along with 5 other adults from there, all clustered with G. vocalis ( Fig. 157 View FIGURE 157 ). Thus, after failing to find physical or genetic characters to reliably separate these initial groupings, we merged them. On the other hand, we still see a definite dichotomy in that almost all individuals of G. vocalis discussed in Weissman et al. (1980), are dark colored with probably one generation/year; while almost all specimens to the east, including those from the Mohave Desert, are light colored and some may have two generations/year. We were also able to cross one male of G. vocalis from Riverside Co. (S03-39) with a female of ‘G. #10’ from Inyo Co. (S03-37) and got good egg hatches.
At Furnace Creek in Death Valley National Park, there were good numbers of G. vocalis singing in both 1980 and 1983, but only one male was heard in 2003 ( Weissman et al. 2012). While G. vocalis , and sympatric G. staccato , G. saxatilis , and G. armatus , which we also heard on earlier visits, were all declining there, non-native Acheta domesticus , which was rare in the early 1980’s, had increased significantly and suggests competitive exclusion.
Tachinid Ormia ochracea emerged from adult males collected at S91-78 (Sedona), S92-58 (Ballona Wetlands), S98-95 (Fallon—with 2 of 5 males parasitized), and S13-14 (Agua Fria—2 of 12 males parasitized). Most interestingly, from a mid-instar nymph collected under a cow pie on 19-iii-1993 (S93-7 at 20.6 km NW Fallon), a dead adult tachinid Exoristoides johnsoni was present in the rearing container on 6-v-1993. It is unknown when the larvae emerged from the cricket but this may represent a mechanism whereby tachinids can pass the winter at localities that experience freezing winter temperatures.
Gershman (2009, 2010) discussed aspects of mating in G. vocalis . Sandford (1987) studied aggression with crickets from Oklahoma: his ‘ G. alogus ’ is uncertain, but not G. vocalis or G. alogus as understood here (because out of range) while his ‘ G. integer ’ is probably G. texensis .







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.