
Hydractinia symbiolongicarpus Buss & Yund, 1989
|
publication ID |
https://doi.org/ 10.5281/zenodo.556851 |
|
publication LSID |
lsid:zoobank.org:pub:985C0239-D00C-457D-B593-76A3081BCEEA |
|
DOI |
https://doi.org/10.5281/zenodo.6015987 |
|
persistent identifier |
https://treatment.plazi.org/id/03A787C7-4936-FF9C-FF58-F8BCFE32FA89 |
|
treatment provided by |
Plazi |
|
scientific name |
Hydractinia symbiolongicarpus Buss & Yund, 1989 |
| status |
|
Hydractinia symbiolongicarpus Buss & Yund, 1989 View in CoL
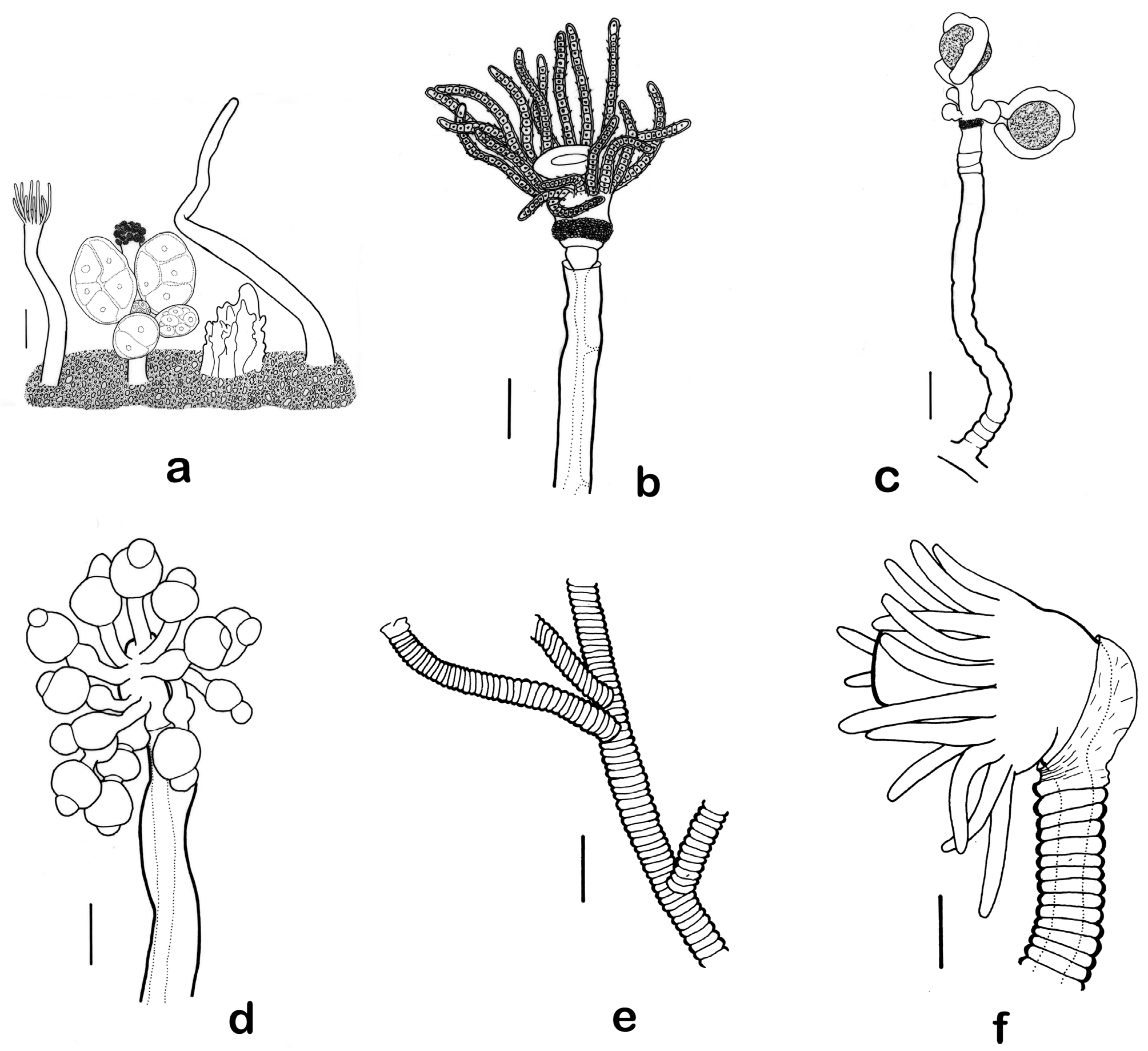
Fig. 13 View FIGURE 13 a
Hydractinia symbiolongicarpus Buss & Yund, 1989: 863 View in CoL , figs. 1, 2.
Type locality. USA: Long Island Sound, Connecticut, Guilford , Harrison Point , 41°16'N, 72°44' W ( Buss & Yund 1989: 868). GoogleMaps
Material examined. NS: Kingsport, 19.vii.1966, low tide, one colony, on gastropod shell occupied by Pagurus longicarpus , with gastrozooids and gonozooids, gonozooids with female gonophores, coll. J.S. Bleakney & K.H. Bailey, ROMIZ B643.
Description. Hydroid encrusting a gastropod shell occupied by pagurid crab Pagurus longicarpus . Colony stolonal, with a dense aggregation zooids arising from an encrustation of naked coenosarc dotted by innumerable small perisarcal spines and a lesser number of much larger, jagged perisarcal spines, grooves of these spines filled with naked coenosarc. Zooids naked, polymorphic, in poor condition but with distinguishable gastrozooids, gonozooids, and dactylozooids. Gastrozooids columnar, varying in height but about 2–3 mm high, terminating in an elongated hypostome; tentacles in a staggered but narrow ring at distal end, tapering from base to tip, about 20 in number, appearing amphicoronate, with ones in upper whorl larger than those in lower whorl. Gonozooids columnar, shorter, lacking tentacles, terminating with a cap of bulbous nematocyst batteries. Dactylozooids tentacle-shaped, tapering from proximal to distal end, located inside orifice of shell adjacent to hermit crab. Colony female, with each gonozooid bearing several gonophores in a whorl.
Gonophores fixed sporosacs lacking tentacles and canals, varying from nearly spherical to oval to irregular in shape, each one with several eggs.
Cnidome. Undescribed.
Remarks. Difficulties identifying species of Hydractinia Van Beneden, 1844a on the basis of traditional morphology alone have long been apparent. For example, Nutting (1901a) compared hydroids from Plymouth, UK, and Woods Hole , USA, and concluded that European populations assigned to H. echinata ( Fleming, 1828) were readily distinguishable from American populations referred to H. polyclina L. Agassiz, 1860 based on the much larger size of their polyps and the smaller number of tentacles on gastrozooids. Fraser (1912) came to a different conclusion after examining specimens from Massachusetts (Vineyard Haven, Tarpaulin Cove, Woods Hole) , Maine ( South Harpswell ) , North Carolina (Beaufort) , Nova Scotia (Canso), and New Brunswick (Grand Manan), and comparing them with colonies from Devon , England (Plymouth) . No consistent differences of taxonomic importance could be found by him between populations from the two sides of the North Atlantic, and he referred them all to a single species, H. echinata ( Fleming, 1828) .
Alternative methods have helped in resolving the taxonomy of this difficult genus. Utilizing breeding experiments, electrophoretic analyses, and morphometrics, Buss & Yund (1989) demonstrated that hydroids commonly identified as Hydractinia echinata from northeastern North America comprised a sibling species group. Moreover, none of these sibling species was conspecific with the European H. echinata . One of the American populations, found as epibionts on shells inhabited predominantly by the hermit crab Pagurus longicarpus between Freeport, Maine, and Guilford, Connecticut, was established as a new species, H. symbiolongicarpus Buss & Yund, 1989 .
Two other species were recognized in the complex. A population sampled between Starboard , Maine, and Nahant , Massachusetts, growing as epibionts on shells of Littorina littorea inhabited by the hermit crab Pagurus acadianus , was referred by Buss and Yund (1989) to H. polyclina L. Agassiz, 1860 . A lectotype of that species, designated as MCZ 9010 View Materials -b, was selected by them from what is taken to have been Louis Agassiz’s syntype material at the Museum of Comparative Zoology ( MCZ). While collection localities of the species were not specified in the original accounts of L. Agassiz (1860, 1862) , A . Agassiz (1865) listed material at MCZ from Grand Manan Island, NB, in the Bay of Fundy , as well as from Massachusetts (Chelsea Beach, Swampscott, Nantucket Shoals, and Nahant). The lectotype specimen was taken from an original collection ( MCZ 55 View Materials ) obtained at Grand Manan during 1859 by A. E. Verrill. Remaining material from that collection, together with specimens in another lot obtained at Grand Manan ( MCZ 56 View Materials ), form part of the paralectotype series. The second species, also recognized as new ( H. symbiopollicaris Buss & Yund, 1989 ), occurred on shells occupied by hermit crabs identified as P. pollicaris . Colonies of that species were collected at Woods Hole, Massachusetts, and Guilford , Connecticut . Of the three species distinguished by Buss & Yund, H. polyclina appears to have a stronger boreal affinity. Hydractiniid hydroids identified earlier from cold waters of the lower Bay of Fundy as H. echinata were most likely based on H. polyclina , a conclusion supported by the collection locality of the lectotype of that species. Hydroids of Hydractinia symbiolongicarpus occurring northwards to estuaries of central Maine were thought to represent either recent introductions or disjunct populations of a species occurring predominantly south of Cape Cod ( Folino & Yund 1998). The species has been shown here to extend even further north into Minas Basin, NS, in the upper Bay of Fundy , where it is also likely disjunct. As for Hydractinia symbiopollicaris , it is not known to occur north of Cape Cod ( Buss & Yund 1989; Cunningham et al. 1991).
Trophosome morphology was found by Buss & Yund (1989) to be of less utility in distinguishing the three sibling species than the identity of the pagurid crabs occupying their substrate shells. Nevertheless, there was a tendency for the chitinous skeleton of the hydroid to grow beyond the shell margin in H. polyclina but not in the others. Colonies of both H. polyclina and H. symbiopollicaris also occasionally included unusual gastrozooids having 3–5 tentacles with significantly thickened bases, alternating with 2-4 shorter tentacles of normal shape. Such polyps were not seen by them in H. symbiolongicarpus .
Questions remain over the taxonomy and biogeography of these putative species. According to Miglietta et al. (2009), preliminary molecular studies suggest that H. symbiopollicaris may be conspecific with H. polyclina . Confirmation of this conclusion is warranted, and both species are currently recognized as valid in WoRMS.
Moreover, Miglietta et al. proposed that H. polyclina is in fact an amphi-Atlantic species, occurring on the coasts of Belgium and Denmark as well as northeastern North America. At those locations in Europe, the species was thought by the authors to have been misidentified previously as H. echinata . That conclusion, too, merits reexamination. Hydractinia polyclina was not included in an account of northwest European anthoathecates by Schuchert (2012).
The occurrence of Hydractinia symbiolongicarpus in Minas Basin is biogeographically noteworthy in that its known distribution elsewhere is entirely to the south of the region. Nevertheless, the basin is hydrographically distinct from much of the rest of the Bay of Fundy, in part because of significantly warmer surface water temperatures during summer. As a result, several species of marine invertebrates, isolated from their primary populations along the Atlantic coast of the United States, are known to occur there ( Ganong 1890b; Bousfield & Leim 1960: 26). Notably, one of the species included in the list of Bousfield and Leim was the hermit crab Pagurus longicarpus . To the list of disjunct species in Bousfield and Leim may be added the relatively warm-water hydroids H. symbiolongicarpus and Obelia bidentata S.F. Clark, 1875 , both reported herein. Eudendrium album Nutting, 1896 , identified during this study from Minas Basin, may be another. Although reported earlier by Fraser (1918) from perpetually cold waters of the lower Bay of Fundy, its actual distribution in the bay may be restricted to areas having seasonally warmer water temperatures. Other similar species occurring near the mouth of the Bay of Fundy ( E. capillare Alder, 1856a ; E. bleakneyi , sp. nov.), could easily be mistaken for E. album .
As noted elsewhere (e.g., Schuchert 2001a, 2008a; Cartwright et al. 2008; Calder et al. 2014), molecular studies have revealed that the genera Hydractinia and Clava Gmelin, 1791 are closely related and should be referred to the same family. A nomenclatural problem arises from this discovery because widely used family names ( Clavidae McCrady, 1859 ; Hydractiniidae L. Agassiz, 1862 ) have been based on the two genera. Prevailing usage so far this century has been to combine them in Hydractiniidae and to abandon Clavidae . While this is counter to the Principle of Priority in nomenclature, it has been justified given the greater number of species assigned to Hydractiniidae and to the supposed greater familiarity of that name. A ruling on the issue from the ICZN is needed in the interests of nomenclatural stability. Meanwhile, prevailing usage has been maintained here.
The binomen H. polyclina has often been attributed to L. Agassiz (1862). Although the species was first described in that publication, the specific name was made nomenclaturally available two years earlier. The name Hydractinia polyclina was used by L. Agassiz (1860) in association with illustrations of the species, thereby meeting criteria of availability (ICZN Art. 12.2.7).
A substantial literature exists on the genetics of allorecognition, and rejection or fusion, of merging conspecific colonies of sessile invertebrates. Hydractinia symbiolongicarpus has been used extensively a model organism in such studies (see Rosengarten et al. 2011, Powell et al. 2011).
Recorded distribution. Bay of Fundy: recorded for the first time.
Eastern North America: Bay of Fundy (this study) to Beaufort, North Carolina ( Miglietta et al. 2009).
Worldwide: As above.
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hydractinia symbiolongicarpus Buss & Yund, 1989
| Calder, Dale R. 2017 |
Hydractinia symbiolongicarpus
| Buss 1989: 863 |