
Ichthyophis multicolor, Wilkinson & Presswell & Sherratt & Papadopoulou & Gower, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3785.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:7EF35A95-5C75-4D16-8EE4-F84934A80C2A |
|
DOI |
https://doi.org/10.5281/zenodo.4910682 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD8F79-7602-1808-20E2-799C5B9E54E3 |
|
treatment provided by |
Felipe |
|
scientific name |
Ichthyophis multicolor |
| status |
sp. nov. |
Ichthyophis multicolor View in CoL sp. nov.
urn:lsid:zoobank.org:act:AAE62A6A-B689-4B18-9A58-599C2993EDED
Figs. 1 – 3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Tables 1 View TABLE 1 – 2
Holotype. CAS 212264 View Materials , an adult female, collected by villagers for Jens V. Vindum on 25 April 2000 from the vicinity of Mwe Hauk Village (26°16’39.2’’N, 94°45’32.5’’E; c. 10 m asl), Ayeyarwady Region, Myanmar. GoogleMaps
Paratypes (n = 13). CAS 212254 – 63 View Materials and 212265 – 67 (eleven females and two males) collected along with holotype .
Diagnosis. A striped Ichthyophis that differs from all other striped congeners except I. tricolor in having a pale venter, typically with whitish markings mid-ventrally, bordered by a darker ventrolateral longitudinal stripe immediately below the pale yellowish lateral stripe on each side. Differs from I. tricolor in having many more AGs (> 325) and vertebrae (> 120) and TAs nearly or more than twice as far from nares than from eyes (N–TA/E–TA> 1.8).
Description of holotype. Some morphometric and meristic data are given in Table 1 View TABLE 1 . Mostly good condition, damaged skin dorsally c. 55 mm anterior to terminus, small depressed scar on right about eight annuli anterior to the vent, several scale pockets opened, small (<15 mm) midventral longitudinal incision 125 mm behind ST with some viscera protruding including bean-shaped eggs (9 x 6.5 mm). Head, nuchal region and trunk dorsoventrally compressed. Girth maximal throughout midbody, increasing very gradually over first 60 mm, decreasing less gradually over last c. 30 mm, L/W = c. 25. Tail upturned towards tip, short (about as long as ST–TA). Head somewhat more V- than U-shaped in dorsal view. In lateral view, distance of CM from top of head a little greater than distance from bottom of head. In ventral view, lower jaws inset from upper jaws, not notably more so anteriorly than at level of TAs. Eye equidistant from lip and top of head in lateral view, not surrounded by narrow whitish ring, eye diameter (c. 0.7 mm) distinctly larger than that of naris and TA, about equal to TP. TAs more than twice as distant from nares as from eyes, fairly close to lips (0.6 mm), on imaginary lines between nares and CMs. Naris approximately level with AM; in lateral view equidistant from top and bottom of head and ST; close to but inset from side of head in dorsal view. Teeth slender, strongly recurved, OM series extends approximately one quarter to one third the length of IM series (with three to five OMs on each side) behind posteriormost IMs. Bluntly tipped tongue strongly plicate (with approximately a dozen longitudinal ridges and grooves) posteriorly, lacking a distinct longitudinal medial groove, margin overlying all except anteriormost IMs. Choanae very narrow, distance between them six or seven times each of their greatest widths. Collar region more massive than adjacent head and body, delimited by strong constrictions. C2 slightly longer than C1 measured laterally. NG1 incomplete dorsally, not pale ventrally. NG3 with gentle middorsal anterior flexure. Four anteromedially flexed, evenly spaced TGs on C2 dorsally, first two short and third poorly developed on right, last longer, crossing midline, not extending as far as lateral stripes.
The first three AGs widely incomplete on venter, otherwise mostly complete midventrally with five (on left) or six (on right) widely incomplete near (i.e. interrupted by) the vent, and five more narrowly incomplete on tail. Dorsally, AGs are mostly complete, those on first two thirds of the body and last eight curve anteromedially, the former gently, the latter very strongly angulate and narrowly incomplete (giving the appearance of a notch) at midline. Ventrally, anterior AGs angulate, degree of posteromedial curvature increasing from two to 2.5 times the length of an annulus at midbody, decreasing posteriorly, orthoplicate by 36th AG anterior to vent. The last few annuli shorter. The terminus ends in a short (1.3 mm) cap, approximately the length of the three preceding tail annuli or two body annuli.
Small (0.6 x 0.4 mm) scattered scales present in a single row in very shallow pocket of first AG on dorsum; posteriorly, four to five rows of large (1.7 x 1.8 mm) scales present in very deep pockets on dorsum, reducing to deep pockets with three to four scale rows on the venter. Margins of vent slightly elevated, formed of twelve main denticulations, six on each side, with some irregular subdivisions. No associated papillae.
In preservation, dorsum brownish grey, more grey on head. A distinct pale spot, wider than long (4.5 x 4 mm), forming a disc around the vent. Narrow, irregular, pale yellowish bilateral stripes, extend unbroken from just in front of CMs to the fifth or sixth last annulus, fading gradually on first annulus behind the vent, almost broken on C2, not visible ventrally on collars, not connecting with or curving towards disc. Immediately below each lateral stripe, running from C2 to the level of the vent, a mostly slightly narrower brown line, a little paler than the dorsal colour, with a gradual but rapid transition to a much paler tan colour that predominates on the venter but which is interrupted by small whitish patches midventrally. Narrow, pale lines along upper margins of lower jaw. TPs and ST pale. Pale midventral line extends across nuchal region to the level of CMs. Except where crossing pale lateral stripes, AGs appear mostly paler than adjacent skin, especially on venter.
Variation. Morphometric and meristic data for the type series are given in Table 1 View TABLE 1 . The sample is trimodal in total length, with eight specimens less than 200 mm, five greater than 350 mm and one at 256 mm. Relative overall proportions (L/W = 25.1 to 28.4) and some features of the head (N– TA /E – TA = 1.9 to 2.6; E – ST/E – E = 1.0 to 1.1) show no correlation with total length and are seemingly unaffected by growth. In contrast, there are some substantial differences related to size. For example, W/S ranges from 4.4 to 5.4 in the eight smallest specimens and from 6.1 to 7.2 in larger specimens, indicating that the width of the stripe does not grow as fast as the width of the body. Similar allometries are apparent in the relative lengths of the head (L/H) and of the tail (L/ T), and the tail appears to lengthen more slowly than the head. The sample includes only two males (both small), and we can reject the hypothesis that it is an unbiased sample from a population with an even sex ratio (p = 0.0129, two-tailed binomial test). The five largest females all have large yolky eggs and all other females have small, non-yolky eggs (<0.5 mm) and might therefore be considered subadult .
The tail is upturned only in the holotype. All paratypes have the stripe beginning closer to the eye than in the holotype, with some slight downturn of the stripe near the vent, and pale markings on the throat are not apparent in CAS 212255 View Materials . Ventral colour varies substantially. All specimens have dark stripes immediately below the pale lateral stripes that give way to a paler tan ground colour on the venter, and all but one ( CAS 212257 View Materials ) have some white mid-ventrally, either as patches or forming an irregular but unbroken stripe that may be very broad (e.g. CAS 212267 View Materials ). In general, the colours are a little more intense in the smaller specimens. Scales are less well developed in the smallest specimens, with no scales in anteriormost annuli and only a single scale row in shallow pockets at midbody and posteriorly in the smallest paratype ( CAS 212259 View Materials ). The right naris of the largest paratype ( CAS 212267 View Materials ) is malformed and especially large. Larger specimens have more teeth in all series but the relative numbers between series does not vary much with size .
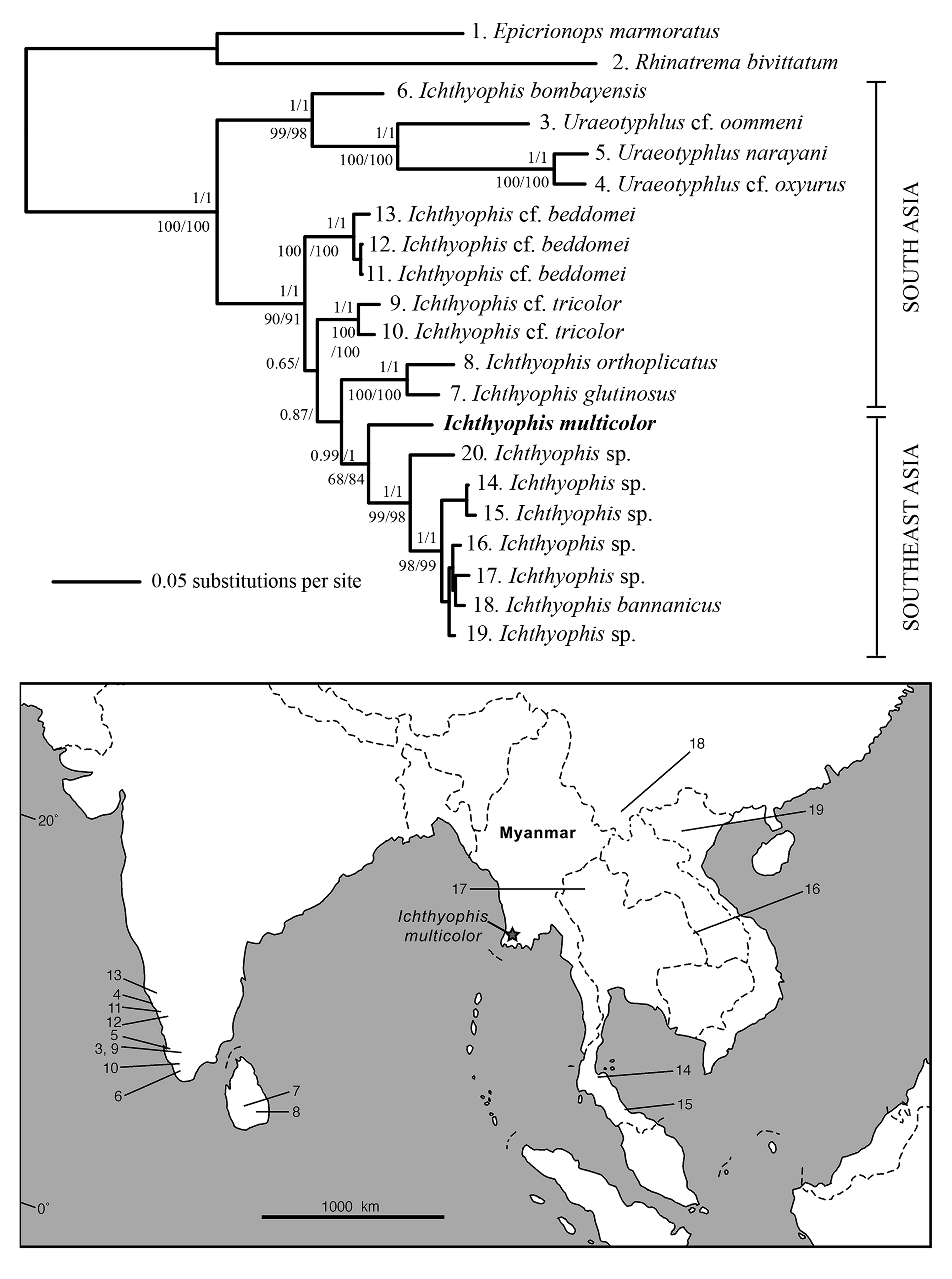
Molecular systematics. There is very little variation among the eight specimens of Ichthyophis multicolor sequenced for the three mt gene fragments (mean nucleotide p-distance 16s: 0.0008, cytb: 0.0010; no variation in 12s), consistent with the interpretation that they are conspecific. Visual inspection of transition and transversion plots (not shown) indicated substantial saturation of cytb third codon positions across the multiple alignment. Partitioning by gene and codon position was favoured over partitioning by gene only in both ML and BI analyses, but the different partitioning schemes had little effect on the phylogenetic results ( Fig. 2 View FIGURE 2 ) which are very similar to those reported by Gower et al. (2002) in that sampled South Asian Ichthyophis are paraphyletic with respect to the monophyletic Southeast Asian Ichthyophis and the relationships among sampled peninsular Indian ( I. beddomei Peters 1879 , I. tricolor ) and Sri Lankan ( I. glutinosus L, I. orthoplicatus Taylor 1960 ) Ichthyophis are not particularly well-supported. I. multicolor is distinct genetically from other sampled species, and is recovered as sister to all other sampled Southeast Asian Ichthyophis .
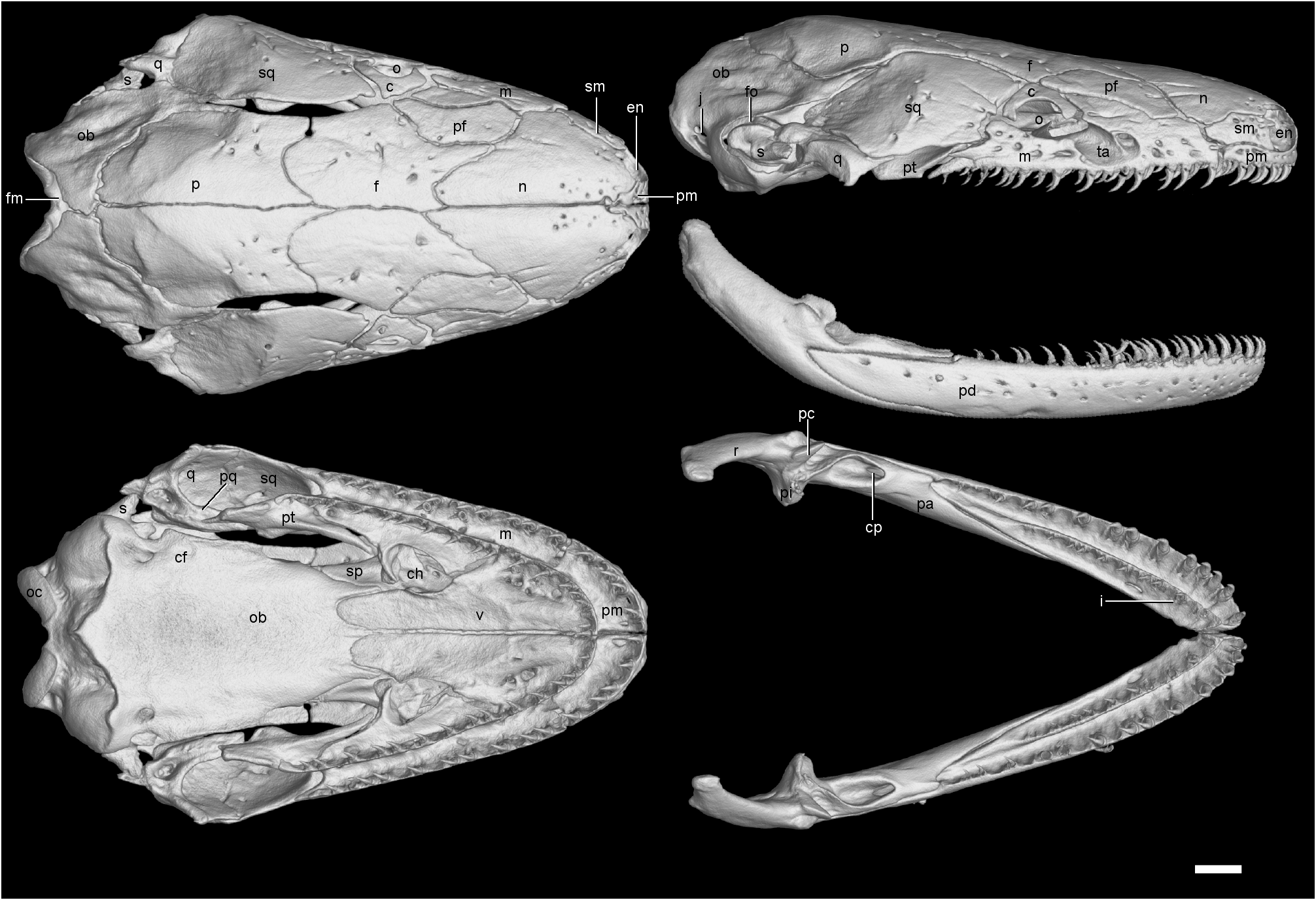
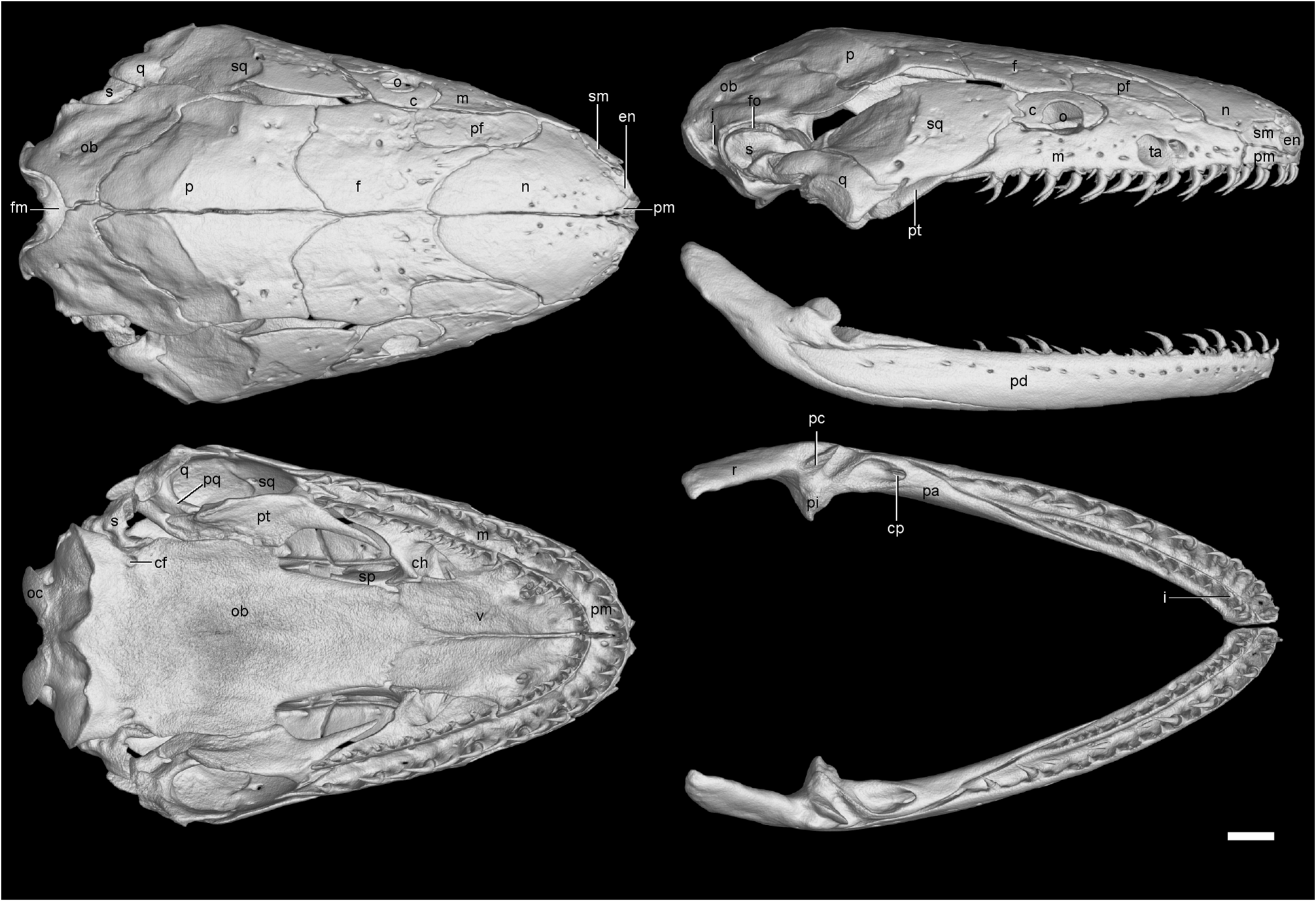
Cranial and mandibular osteology. A reconstruction of the skull and mandible of one large, female paratype (CAS 212266) of Ichthyophis multicolor is shown in Figure 3 View FIGURE 3 and similar illustrations of Ichthyophis kohtaoensis Taylor 1960 (a ‘typical’, or at least not obviously unusual, Southeast Asian congener) and I. tricolor (the most similarly coloured congener) are shown in Figures 4 View FIGURE 4 and 5 View FIGURE 5 . There are no differences between these species and the type species of the genus, I. glutinosus , in terms of the number of separate skull bones and their general arrangement ( Wilkinson et al. 2011: fig 5). Comparison of these three reconstructions ( Figs. 3 – 5 View FIGURE 3 View FIGURE 4 View FIGURE 5 ) immediately suggests several notable differences among the three species. However, some of these differences do not stand scrutiny as interspecific because they are variable among the other specimens examined. Thirteen characters that, based on our sampling, do represent interspecific differences separating one of the three species are reported in Table 2. Several of these differences are substantial and obvious, such as the relative size of teeth in the outer rows, and whether the tentacular canal is open within the maxillopalatine or roofed with bone.
Notable differences among the three specimens illustrated in Figs. 3 – 5 View FIGURE 3 View FIGURE 4 View FIGURE 5 which vary within species are: the presence or absence of prefrontal-septomaxilla contact; the relative size of the upper temporal fenestra; the relative breadth of the pterygoid in palatal view; the relative width of the skull roof level with the parietal-frontal suture. In a simple count of similarities summarized in Table 2, Ichthyophis multicolor and I. tricolor are about as similar to each other as are I. multicolor and I. kohtaoensis , with the least similar pair of species being I. tricolor and I. kohtaoensis .
Etymology. Named for its having more distinct colours than most other Ichthyophis . For nomenclatural purposes the specific epithet is considered to be a noun in apposition.
Suggested English name. Colourful Ichthyophis or Colourful Ich.
Distribution and natural history. This species is known only from the type locality. Specimens were collected on the surface after heavy rain from an area of secondary forest and agriculture. The soil was sandy and hard packed. Local people brought the field team to the locality in response to questions about where ichthyophiids could be found. Although the type series includes no larval specimens it seems very likely that the species, like all other Ichhyophis as far as is known, is oviparous with an aquatic larval stage. This life history is also suggested by the large size of the unlaid eggs (e.g. Wilkinson & Nussbaum 1998).


| V |
Royal British Columbia Museum - Herbarium |
| TA |
Timescale Adventures Research and Interpretive Center |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |