
Leptolalax (Lalos) bourreti Dubois, 1983
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3147.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/A83A879E-FFB5-9312-5AE3-F92CFB30FF03 |
|
treatment provided by |
Felipe |
|
scientific name |
Leptolalax (Lalos) bourreti Dubois, 1983 |
| status |
|
Leptolalax (Lalos) bourreti Dubois, 1983 View in CoL
( Fig. 10–13 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 )
Megophrys pelodytoides View in CoL — Bourret 1937: 17; Bourret 1942: 208, Fig. 45 (a2).
Leptolalax bourreti Dubois, 1983: 150 View in CoL .
Onymophoront, by original designation as “ holotype ”: holophoront, MNHN 1938.0094 View Materials , subadult male, SVL 36.2 mm. Onymotope : Chapa [Sa Pa] (22°21’ N, 103°52’ E), Lao Cai Province, Vietnam. Collected by René Bourret , before 1937. Material examined. Vietnam. Sa Pa, Hoang Lien Nature Reserve (22°19’N, 103°47’E): MNHN 1938.0094 View Materials , subadult male, holophoront, coll. René Bourret GoogleMaps , before 1937; MNHN 1938.0093 View Materials , MNHN 1938.0095 View Materials , adult females, other protaxonts (paratypes) ; MNHN 1999.5659 View Materials – 5660 View Materials , 1999.5665 View Materials , MNHN 1999.5669 View Materials – 5671 View Materials , adult males, coll. Annemarie Ohler, Stéphane Grosjean & Steven Swan , 8 November 1997, 20 January 1998, 19 February 1998 and 28 July 1998; MNHN 1998.8702 View Materials – 8742 View Materials , tadpoles , 19 and 21 July 1998; MNHN 1998.8743 View Materials – 8763 View Materials , tadpoles , 28 July 1998, in the quiet part of a small torrent; MNHN 1998.8764 View Materials – 8769 View Materials , tadpoles , 02 August 1998; MNHN 1998.8770 View Materials – 8774 View Materials , tadpoles , 5 August 1998, at 1860 m a.s.l. in a large torrent more than 5 m wide with a strong current, coll. Stéphane Grosjean.
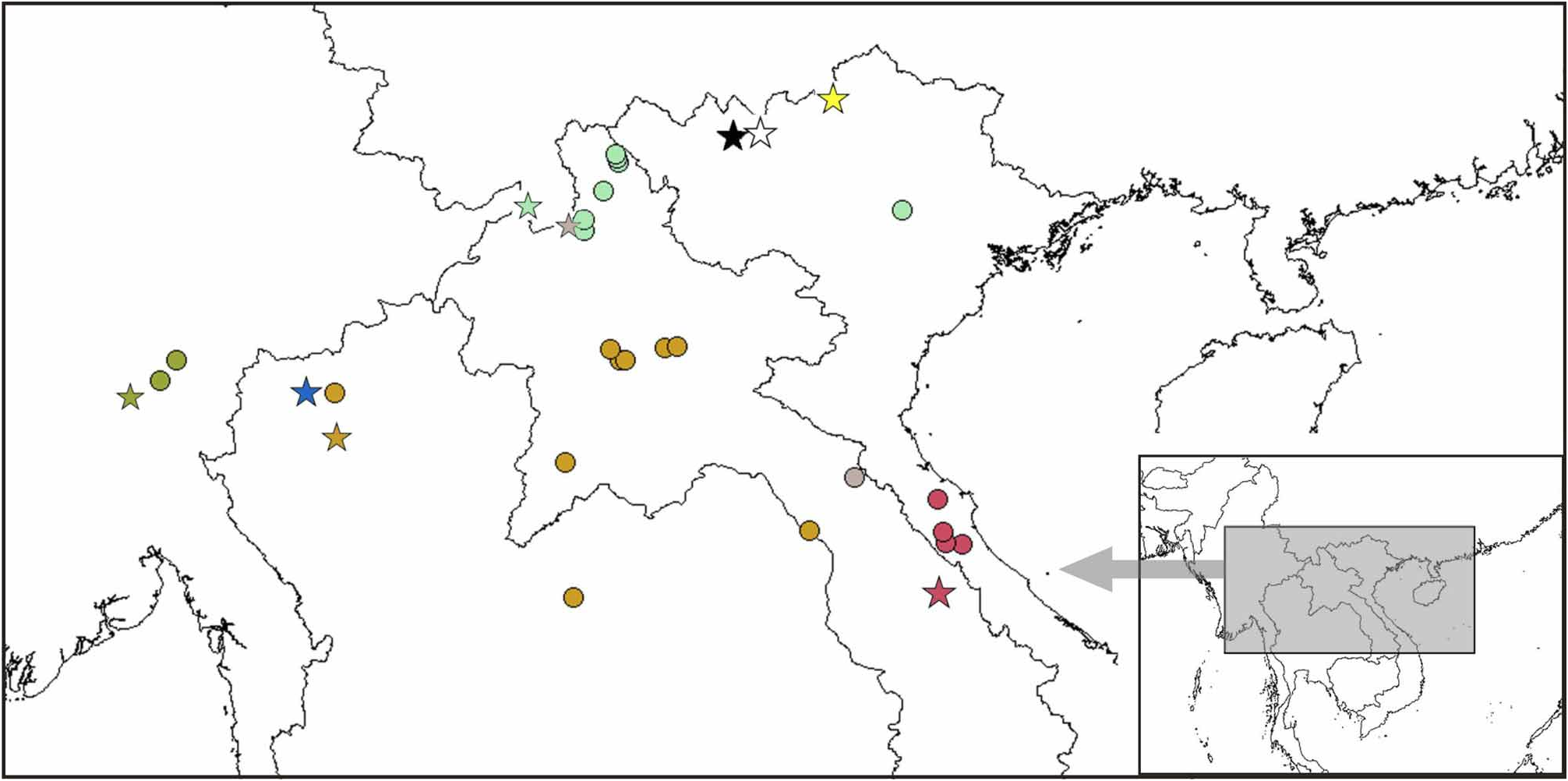
Comments. Dubois (1983) described this large species from a series of frogs collected by Bourret in 1937 and considered by the collector as Megophrys pelodytoides ( Bourret 1937: 17; 1942: 208). Subsequent collections of Leptolalax in Sa Pa ( Ohler et al. 2000: 74) consisted of specimens of slightly smaller body size but which can be allocated to L. bourreti by coloration and morphological characters. A specimen from Ha Giang, Vi Xuyen, Cao Bo (AMNH A163810) mentioned as L. bourreti by Frost et al. (2006), sequence available on Genbank (accession number DQ283381 View Materials ), is morphologically quite different from the L. bourreti hypodigm ( Dubois 1983) and groups in a distinct clade from L. bourreti onymotopic specimens (see description below).
As described below under Variation, this species apparently exhibits variation in iris colour and other chromatic characters. Especially iris colour is often species-specific in frogs and we cannot exclude that these differences point to the existence of more than one species currently subsumed under bourreti . However, for the time being we have no evidence that there is more than one species as the iris colour differences in Sa Pa cannot be correlated to other morphological or colour differences, and insufficient molecular data are available to analyse this question further.
Diagnosis. Member of the subgenus Lalos based on the presence of a lateroventral gland and molecular phylogenetic relationships ( Fig. 4 View FIGURE 4 ), distinguished from other species included in this subgenus by the following combination of characters: large-sized species (males 28.0– 36.2 mm; females 42.0–45.0 mm) ( Table 3) with small webbing and distinct fringes on toes; tympanum distinct; dermal ridges under toes poorly distinct; finger tips slightly enlarged; dorsal skin smooth (Table 4); dorsum reddish, greenish or brown with dark spots; moderate sized dark spots on flanks; ventral side whitish; iris colour variable, clearer above, copper, green or brownish ( Table 5).
Description of the holophoront, MNHN 1938.0094. Size and general aspect. (1) Specimen of moderate size (SVL 36.2 mm), body slender.
Head. (2) Head of large size, narrower (HW 13.4 mm) than long (HL 14.1 mm; MN 12.1 mm; MFE 9.5 mm; MBE 4.9 mm), flat. (3) Snout not protruding, its length (SL 5.5 mm) longer than horizontal diameter of eye (EL 4.7 mm). (4) Canthus rostralis rounded, loreal region slightly concave, obtuse. (5) Interorbital space flat, as large (IUE 3.4 mm) as upper eyelid (UEW 3.4 mm) and internarial distance (IN 3.5 mm); distance between front of eyes (IFE 5.8 mm) about half of distance between back of eyes (IBE 10.1 mm). (6) Nostrils closer to eye (EN 2.7 mm) than to tip of snout (NS 2.9 mm). (8) Tympanum (TYD 1.9 mm) distinct, rounded; about two-fifths of eye diameter, tympanum-eye distance (TYE 1.6 mm) slightly smaller than its diameter. (11) Tongue large, rounded.
Forelimbs. (12) Forearm short, thin (FLL 9.5 mm), longer than hand (HAL 8.7 mm), not enlarged. (13) Fingers long and thin (TFL 5.7 mm). (15) Tips of fingers slightly enlarged. (16) Fingers II and III with dermal fringe.
Hindlimbs. (19) Shanks about four times longer (TL 17.8 mm) than wide (TW 3.7 mm), longer than thigh (FL 16.0 mm) and distance from base of internal metatarsal tubercle to tip of toe IV (FOL 16.8 mm). (20) Toes rather short and rather strong; toe IV (FTL 9.9 mm) longer than one-third of distance from base of tarsus to tip of toe IV (TFOL 26.8 mm). (22) Tips of toes not enlarged. (23) Webbing present, very small; fringes present, distinct: I 2 – 3 II 2 – 3 III 2 ½ – 4 IV 4 – 2 ½ V (MTTF 6.5 mm; MTFF 6.3 mm; TFTF 9.6 mm; FFTF 10.3 mm). (24) Dermal ridge along toe V present, from tip of toe to base of finger, poorly developed. (26) Inner metatarsal tubercle short, distinct; its length (IMT 1.6 mm) 2.4 times in length of toe I (ITL 3.8 mm).
Skin. (29) Dorsal and lateral parts of head and body: snout, region between eyes, side of head, back and flank smooth (possibly due to preservation). (30) Supratympanic fold poorly prominent, from eye to above upper forelimb. (31) Dorsal parts of limbs: smooth. (32) Ventral parts of head, body and limbs: smooth. (33) Presence of macroglands: a suprabrachial and a large oval axillary gland present; femoral glands and lateroventral gland indistinct.
Coloration. In alcohol (pattern and coloration modified due to preservation): (34) Dorsal and lateral parts of head and body: light grey brown. (35) Dorsal parts of limbs: light grey brown with indistinct brown crossbands; posterior part of thigh light grey brown. (36) Ventral parts of head, body and limbs: greyish; webbing greyish.
Male secondary sexual characters: (38) Vocal sacs: no opening on mouth floor.
Variation. Bourret (1937) described the coloration in life of the Sa Pa specimens mentioned as Megophrys pelodytoides as follows (translation): “Patterns and colour very variable; back grey brown verging more or less to red, dark grey green, more or less clear or dark brown, dirty green or blackish; transversal spots on lips and hindlimbs dark. Underside whitish. Dark spots more or less distinct, marbling, or symmetrical markings sometimes with light outline. Often fine light dots, greenish white above, yellow on the flanks and on the shanks. A light vertical spot on the tip of snout seems to be constant; an orange tympanic spot occurs frequently. The iris is also variable, dotted with dark green-brown, a little clearer above and below than on the sides; yellow above, grey brown below; light yellowish brown; light brown-yellow reticulated with dark brown; coppery, green or brownish green, finely reticulated with black, spotted with red in its upper part.” The first author noted life coloration of MNHN 1999.5659 in the field: Iris copper on upper part, golden in lower part separated by a black horizontal band. Dorsum and upper part of flank brown with darker spots, lower part lighter brown with black brown spots; loreal and tympanic region brown with a dark black brown band on canthus rostralis and tympanic fold; tympanum black brown; upper lip brown with black brown bands; forelimbs and hindlimbs brown with darker crossbands; posterior part of thigh brown with darker spots; throat ivory white, transparent; margin of throat brownish with white spots; chest light reddish; belly yellow getting orange in posterior part; webbing light brown; macroglands light orange.
All the seven males with large testes collected from Sa Pa in 1998 considered as adults, have vocal sac openings in the mouth. As in all other species of Leptolalax the adult males have internal vocal sacs, we consider that the male holophoront of Leptolalax bourreti , not exhibiting such structures, is a subadult.
Females have large (diameter 2.01 mm), yellowish white oocytes in ovary. The specimen MNHN 1938.0093, adult female, part of the hypodigm, exhibits a still distinct colour pattern: Dorsum fawn with dark brown spots on back and upper flank. A dark brown line on canthus and supratympanic fold; dark bands on upper lip, forelimbs and hindlimbs. The specimens originally mentioned by Bourret (1937) vary in coloration, but show spots and markings on back, limbs and head.
The snout-vent length of nine adult females varies from 41 to 54 mm. Six adult males collected in 1998 are smaller and measure 29.5–34.0 mm and are smaller than the two males measured by Bourret (36.2 in the holophoront). The only adult female in the hypodigm ( Dubois 1983) measures 43.8 mm and falls into the range given by Bourret (1937).
All recently collected specimens of L. bourreti exhibit macroglands. The lateroventral glandular ridge is composed of a series of glands that form a continuous line. Femoral and axillary glands are rounded or oval; whereas the femoral glands are always clearly distinct, the axillary glands are poorly distinct in a few specimens. The suprabrachial glands are of round or oval shape and smaller than the femoral and axillary glands. In specimens reaching metamorphosis, the lateroventral, femoral and suprabrachial glands are distinct, but the axillary gland becomes distinct only when metamorphosis is completed.
Tadpole. Presently two species of Leptolalax are known to occur in the Hoang Lien mountain range: L. bourreti and L. pluvialis . They can be distinguished based on extent of webbing and on colour pattern as these characters are already differentiated when tadpoles enter metamorphosis. On this basis we ascribed the tadpoles collected from Sa Pa to L. bourreti although more species could be found in this area. The tadpoles described below are common in the Hoang Lien Nature Reserve (Sa Pa) and were collected in three different places. On one occasion, no adult Leptolalax was observed in the area, on the second occasion adult of L. bourreti were collected with the tadpoles and in the third occasion tadpoles were collected in the onymotope of L. pluvialis (the two species are known to occur syntopically in this area).
The external morphology is based on a specimen in stage 31 (MNHN 1998.8737; TOL 59.2 mm and BL 18.9 mm).
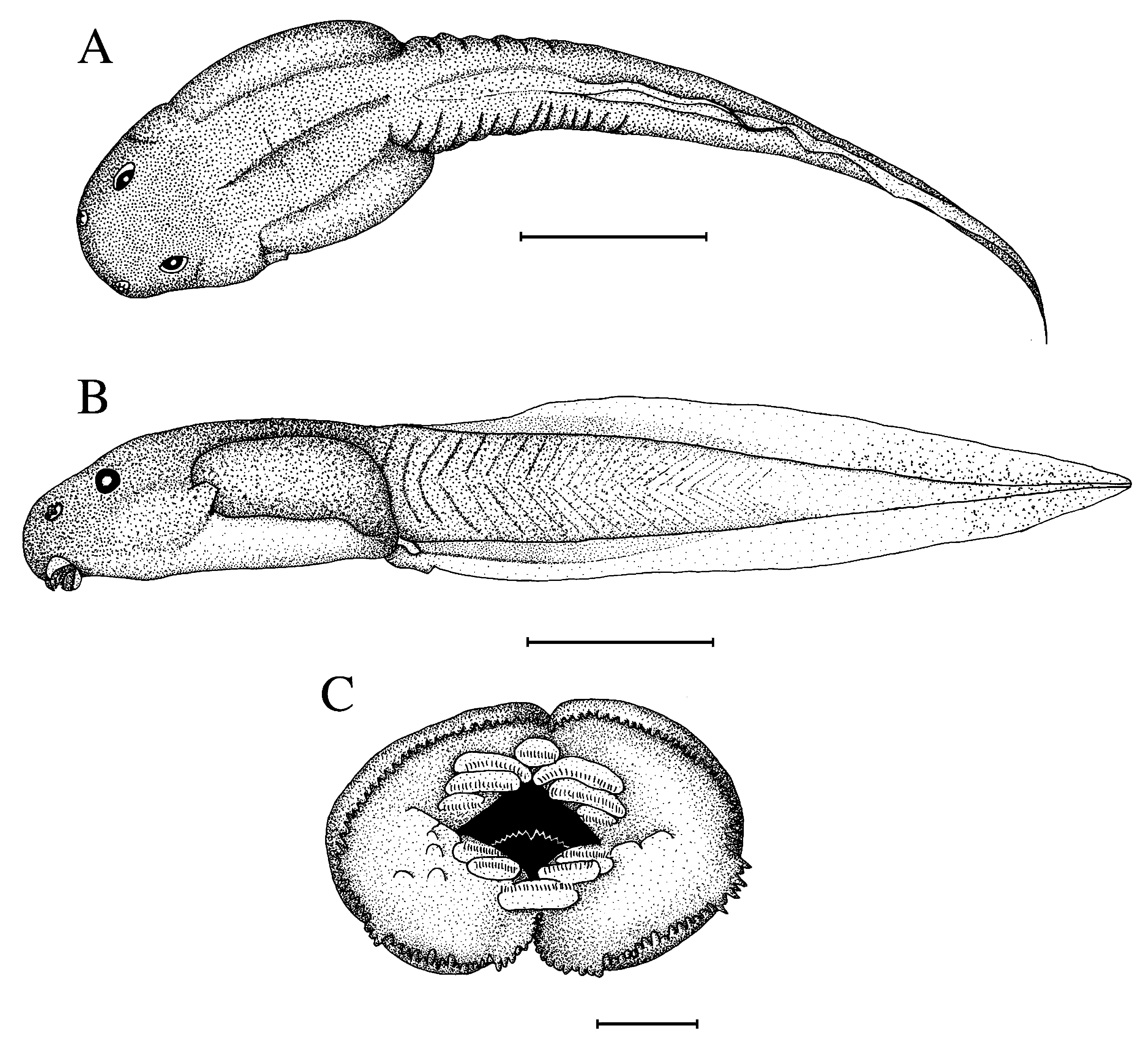
In dorsal view ( Fig. 13A View FIGURE 13 ), body elliptical, widest at the middle of the intestinal coil, snout semi-circular, a pair of large lateral lymphatic sacs extending from spiracle to end of body. In lateral view ( Fig. 13B View FIGURE 13 ), body depressed, BW 143 % of BH, highest just behind the spiracle; snout rounded. Eyes moderately small, ED 7.5 % of BL, slightly bulging and not visible in ventral view, positioned almost dorsally and directed more laterally than anterodorsolaterally. Pineal ocellus not visible; nasolacrymal ducts not visible. Nares round with an oval opening, moderately large, rimmed with four small lobes, positioned almost dorsally but directed anterolaterally with an opening direction dorsolateral, closer to snout than to pupils, RN 40 % of NP; NN 82 % of PP. Spiracle sinistral, square, moderately small, internal wall fused with body wall except its tip, positioned just below the lateral axis and situated at equal distance from snout to end of body, SS 52 % of BL, oriented posterodorsally; its opening closed against body wall, directed posterodorsally, at the height of the apex of myotomes of the caudal muscle. Tail long, TAL 209 % of BL; tail musculature well developed, TMH 71 % of BH and 61 % of MTH, TMW 55 % of BW, the lower part slightly bulging in the proximal quarter then gradually tapering, reaching tail tip. Upper fin shallow in its proximal third then moderately shallow, UF 30 % of MTH, not extending onto body, SU 97 % of BL, almost nonexistent in its proximal sixth then increasing rather abruptly, the remaining part of about equal height, slightly convex and following caudal muscle; lower fin shallow in proximal third then moderately shallow, LF 26 % of MTH, straight and so increasing in size anteroposteriorly; point of maximum height of tail located just before the proximal third, MTH 116 % of BH, tail tip finely rounded. Anal tube moderately large, as a short tube, dextral and directed posteriorly, entirely attached to ventral fin; opening large, lateral, bevelled, posterolaterally directed. Lateral line system consisting of a dorsal body line which begins at the rear of eye and passes just above the lateral sac, a middle body line which joins the dorsal body line at midway of tail; on the body, the angular line curves behind the eye and passes on the upper surface of the lateral sac, the ventral body line, parallel and posterior to the latter, passes laterally on the lateral sac, an unnamed line in the terminology of Lannoo (1987) starts from the spiracle and passes ventrally on the lateral sac; anteriorly, anterior oral line, longitudinal oral line, supraorbital line and infraorbital line present, a short line joinning the supraorbital and the infraorbital lines anterior to eye and another joining these two lines anterior to naris; a short unnamed line lying ventroposteriorly almost parallel to body; the system is symmetrical on both sides of the tadpole. Lateral line organs dash-shaped, all longitudinally oriented except those of the dorsal line which are perpendicular to the longitudinal axis. A pair of small axillary glands ventrally at the level of the gills; another one on the posterior part of flanks, at a short distance before the insertion of tail, halfway between the insertion of hind limbs and the apex of the myotomes of the caudal muscle; a unilateral gland on the left side just before the spiracle.
Oral disc ( Fig. 13C View FIGURE 13 ) cup-like, transversely elongate at rest, positioned and directed ventrally, not emarginated laterally, moderately large, ODW 27 % of BL and 45 % of BW. Lower labium divided by a medial notch, upper labium with four lobes, the median two separated by a deeper fold forming a median fold. A continuous row of marginal papillae all around the oral disc, papillae of small size, conical and pointed, the papillae of the upper labium with a larger base (triangular) than those of the lower labium, finer; eight submarginal papillae present laterally to keratodont rows of the lower labium and anteromedially to row P3. Denticulate papillae present on the lower labium. KRF 1:3+4/2+2:1, keratodont rows short, A3> A4> A2> A5> A1, A1 very short, A2 shortly interrupted in the middle, the other lower rows separated by the upper beak; P1>P2>P3; A1 very short, one-third of P3; P3 half of P2. Keratodonts scythe blade-shaped. Jaw sheaths strong, coarsely serrated, black with a brown blurring just above the serrations; upper sheath a large arch with a stronger and bigger median serration; lower sheath as an open V.
Coloration in life: body and caudal muscle brown olive with lighter spots on fore part of head; numerous spots formed by iridiophores. Fins light olive with a few darker spots on distal part.
Colour in preservative: Upper part of body and upper flanks grey brown, extension of caudal muscle onto body light brown; lower flanks in gradation from brown to grey. Ventral part grey transparent. Caudal muscle light brown mottled with ochre brown, especially in proximal third of upper part, ventral side lighter. Fins transparent grey, upper fin brownish proximally. Nares and hindlimbs white.
The variation is based on 10 other specimens, except where noted (MNHN 1998.8726–8727, 1998.8731, 1998.8738–8739, 1998.8742, 1998.8747, 1998.8762, 1998.8772, 1998.8774, in stages 25–36, TOL 38.2–56.0 mm and BL 10.9–20.8 mm). Only the ratio values different from those of the described specimen are given: BW 115– 138 % of BH (n = 9); ED 6.3–10.1 % of BL; RN 41–61 % of NP; NN 75–88 % of PP; SS 45–53 % of BL; TAL 157–261 % of BL; TMH 59–74 % of BH; TMH 54–67 % of MTH; TMW 52–59 % of BW (n = 8); UF 29–35 % of MTH; LF 24–28 % of MTH; SU 101–133 % of BL; MTH 102–120 % of BH; ODW 22–28 % of BL; ODW 44–48 % of BW. The more the tadpole is in an advanced stage, the more the upper fin begins close to body end. The KRF vary from 1:3+3/2+2:1 to 1:4+4/3+3:1, the additional keratodont rows appears between stages 27 and 29.
Distribution. Vietnam: Lao Cai Province ( Fig. 9 View FIGURE 9 ).
Natural history. One of the female specimens of hypodigm (MNHN 1938.0095) has large oocytes, but those of the other females are half size. Adult males from Sa Pa (MNHN 1999.5659–5660, 1999.5665–5666, 1999.5669– 5671) were collected in January, February, July and November and all have vocal sac openings on the mouth floor.
Specimens collected in 1998 were found in tropical montane deciduous forest at altitudes from 1150 to 1815 m near streams, sitting on leaves, on tree trunks or on the ground on dead leaves at a height of 0–0.5 m and at a distance of 0–50 m from the stream. When breeding, these frogs were usually in or near (0.4 m) the water. This species was abundant as compared to the other species observed in Sa Pa at the study period (more than 26 records). Few were observed in agricultural land, scrub and savannah, most were observed in submontane and montane forest near rivers and streams.
The tadpoles live in mountain streams of variable size and current strength, but in the low-flow parts, in pools often created before small dams formed by dead vegetation lying across streams, but always in parts directly connected to the main water and where current is present. Other tadpoles found in the same streams belong to Leptobrachium ailaonicum , Xenophrys sp. , Quasipaa verrucospinosa and Amolops sp.
Etymology. Dedicated to René Bourret (1884 – 1957). This nomen is a patronym in the genitive.
Placement in molecular phylogeny. Specimens allocated to L. bourreti apear in a highly supported clade that groups L. eos , L. “ oshanensis ” and Leptolalax sp. from Doi Chiang Dao. They are included in a poorly supported subclade with L. “ oshanensis ” AY561306 View Materials and Leptolalax sp. From Doi Chiang Dao which is sister group of L. eos and L. “ oshanensis ” AY526215 View Materials . The molecular differentiation within this cluster of forms is not high, and the relationships and status of the various lineages included in this clade require further study.
Conservation status. Recent observations show that the species is abundant in its habitat. It has been observed in a rather restricted area, but this is clearly due to lack of exploration and difficulty in identifying the species. We assume that this species should be qualified as Least Concern but suggest to keep it as Data Deficient until the expected additional locality records become available and its taxonomic status confirmed.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leptolalax (Lalos) bourreti Dubois, 1983
| Ohler, Annemarie, Wollenberg, Katharina C., Grosjean, Stéphane, Hendrix, Ralf, Vences, Miguel, Ziegler, Thomas & Dubois, Alain 2011 |
Leptolalax bourreti
| Dubois, A. 1983: 150 |
Megophrys pelodytoides
| Bourret, R. 1942: 208 |
| Bourret, R. 1937: 17 |