
Lateothela grandiflora ( Tixier-Durivault & d'Hondt, 1974 ) Moore & Alderslade & Miller, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015375 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87ED-3E1F-FFC7-FF4B-E59F7F3EDD76 |
|
treatment provided by |
Plazi |
|
scientific name |
Lateothela grandiflora ( Tixier-Durivault & d'Hondt, 1974 ) |
| status |
comb. nov. |
Lateothela grandiflora ( Tixier-Durivault & d'Hondt, 1974) View in CoL new combination
( Figs. 68–89 View FIGURE 68 View FIGURE 69 View FIGURE 70 View FIGURE 71 View FIGURE 72 View FIGURE 73 View FIGURE 74 View FIGURE 75 View FIGURE 76 View FIGURE 77 View FIGURE 78 View FIGURE 79 View FIGURE 80 View FIGURE 81 View FIGURE 82 View FIGURE 83 View FIGURE 84 View FIGURE 85 View FIGURE 86 View FIGURE 87 View FIGURE 88 View FIGURE 89 )
Anthothela grandiflora ( Sars, 1856) View in CoL : Broch 1912b (?part): 5–9, Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Molander 1918 (?part): 6–8, Fig. 1 View FIGURE 1 ; Kükenthal 1919 (?part): 17, 19, 26, 43–44, 672, 681–685, 730, 788, 796, Figs. 17 View FIGURE 17 , 315; Verrill 1922 (?part): 18–19, Fig. 2 View FIGURE 2 , Pl. VI Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Kükenthal 1924 (?part): 14–16, Figs. 13–14 View FIGURE 13 View FIGURE 14 ; Stiasny 1937 (?part): 20–23, Figs. F1, F2, Pl. I Figs. 6 View FIGURE 6 , 7 View FIGURE 7 ; Verseveldt 1940 (?part): 37–47, Figs. 13–15 View FIGURE 13 View FIGURE 14 View FIGURE 15 ; Carlgren 1945 (?part): 33–34, Fig. 8 View FIGURE 8 ; Grasshoff 1981 (?part): 745, Karte 1, 942.
Parerythropodium grandiflorum Tixier-Durivault & d’Hondt, 1974: 1373 , Figs. 9–10 View FIGURE 9 View FIGURE 10 ; Weinberg, 1977: 148
Alcyonium grandiflorum ( Tixier-Durivault & d'Hondt, 1974) : Stokvis & van Ofwegen 2006: 165–167; Figs. 1a View FIGURE 1 , 2–3 View FIGURE 2 View FIGURE 3 , 4 View FIGURE 4 a–c
Material examined. RMNH Coel. 33874, S of Raso, Cape Verdo, “ Tydeman ” Cape Verde Islands Expedition 1986, CANCAP-VII, stn. 7.140, 16.583°N 24.60°W, depth 1200 m, 4th Sept 1986 GoogleMaps ; NTNU-VM 63143 View Materials , Rødberg , Trondheimsfjord, Norway, 63.468°N, 10.0°E, depth 200–300 m, 5th December 2006 GoogleMaps ; NTNU-VM 68106 View Materials , Agdenesflua , Trondheimsfjord, Norway, 63.656°N, 9.766°E, depth 202–291 m, 12th June 2012 GoogleMaps ; NTNU-VM 67147 View Materials , Agdenesflua , Trondheimsfjord, Norway, 63.656°N, 9.766°E, depth 150– 50 m, 30th June 2011 GoogleMaps ; NTNU-VM 63140 View Materials , Agdenesflua , Trondheimsfjord, Norway, 63.651°N, 9.763°E, depth 84–147 m, 1st July 2008 GoogleMaps ; NTNU-VM 40341 View Materials (part), Dyrviknes 27, Trondheimsfjord, Norway, 63.603°N, 9.757°E, depth 120 m, 18th May 1965 GoogleMaps ; NTNU- VM 40336, Trondheimsfjord , Norway, determined by Broch as Anthothela grandiflora , depth and date unknown ; NTNU-VM 39877 View Materials , Rødberg , Trondheimsfjord, Norway, depth 400–500 m, 9th September 1911 ; ZMUB unregistered, Norway, Zoological Museum of Bergen , collected by Håkon Mosby-Møre, stn. 1, 10th October 2005 ; ZMUB 60246 View Materials , Handangerfjord , Norway, determined by A. Fosshagen as Trachymuricea kukenthali , 59.813°N, 5.586°E, depth 180–260 m, 6th June 1959 GoogleMaps ; ZMUB 17759 View Materials (part), Skarnsundet , Trondheimsfjord, Norway, determined by Storm as Anthothela grandiflora , depth unknown, August 1899 ; ZMUB 12120 View Materials , Totlandsholmen , Bryggen, Nordfjord, Norway, determined by Grieg as Anthothela grandiflora , depth 450 m, July 1899 ; ZMUB 3897 View Materials , Rødberg , Trondheimsfjord, Norway, depth and date unknown ; ZMUB 548 View Materials , Flora , Batalden, Norway, depth and date unknown ; NHM, UIOslo B1367, Rødberg , Trondheimsfjord, Norway, determined by Jungerson as Anthothela grandiflora , depth and date unknown ; NHM, UIOslo B1368, Between Rūltetangen and Solsvik, Norway, depth 400–500 m, 21st October 1949 ; ZMUC-ANT-000467, Rødberg , Trondheimsfjord, Norway, determined by Mortensen as Anthothela grandiflora , depth 300 m, 27th July 1911 ; ZMUC-ANT-000468, Trondheimsfjord , Norway, determined by Jungersen as Anthothela grandiflora , depth unknown, 27th April 1887 ; ZMB 5527 View Materials (part), Rødberg , determined as Anthothela grandiflora by Broch, Trondheimsfjord, Norway, depth 300–350 m, 1913 ; ZMB 5847 View Materials , Skarnsund , Trondhjemsfjord, Norway, Scholtlaener Expedition 1911, depth 150– 240 m, 1911 ; ZMB 2686 View Materials , Trondheimsfjord , Norway, depth 180 m, 1886 ; ZMB 2545 View Materials , Trondheimsfjord , Norway, depth unknown, 1881 ; NHMUK 1962.7 View Materials .20.210, Trondheimsfjord , Norway, H. Graham Cannon, depth and date unknown ; NHMUK 1898.5 View Materials .5.38, Rødberg , Trondheimsfjord, Norway, Norman Collection, depth 457 m, date unknown ; NHMUK 1917.6 View Materials .7.1, Pernambuco Plateau, east coast of Brazil, determined as Stereogorgia claviformis , 7.617°S, 34.433°W, depth 274 m, date unknown. GoogleMaps
Other material: USNM 1139021 View Materials , West Florida slope, Gulf of Mexico, USA, USGS Discovre GOM 2009, Lophelia II (JSLII-09-GOM-3722), DSRV Johnson Sea Link II, RV Seward Johnson, 26.204°N, 84.727°W, depth 498 m, 16th September 2009 GoogleMaps ; TMAG K4272, Gulf of Mexico , USA, Lophelia II, stn. 276 (JSLII-09-276), 28.441°N, 89.318°W, depth 541 m, 10th September 2009 GoogleMaps ; USNM 1207952 View Materials , Gulf of Mexico, USA, Lophelia II, (LII-10-312), ROV Jason , NOAA Ship Ronald H. Brown, 29.166°N, 88.017°W, depth 489 m, 22nd September 2010 GoogleMaps ; USNM 1207953 View Materials , Gulf of Mexico, USA, Lophelia II, (LII-10-352), ROV Jason , NOAA Ship Ronald H. Brown, 26.336°N, 84.756°W, 507 m, 1st October 2010 GoogleMaps .
The holotype of Parerythropodium grandiflorum is held at the MNHN, Paris and it was not possible to examine it during this project. However, Stokvis & van Ofwegen 2006 examined the holotype and assigned the specimen RMNH Coel.33874 to this species based on their comparison. We were fortunate to be able to examine their material and some figures are included here ( Figs. 68–71 View FIGURE 68 View FIGURE 69 View FIGURE 70 View FIGURE 71 ). A sequence of the mtMutS gene region had previously been obtained from this specimen by Dr Catherine McFadden (Harvey Mudd College, USA) and was able to be included in the phylogenetic analysis within (attempts to sequence igr1– cox1 were unsuccessful). Most of the more detailed description further below is based on a representative specimen from the Trondheimsfjord in Norway (NTNU-VM 63143).
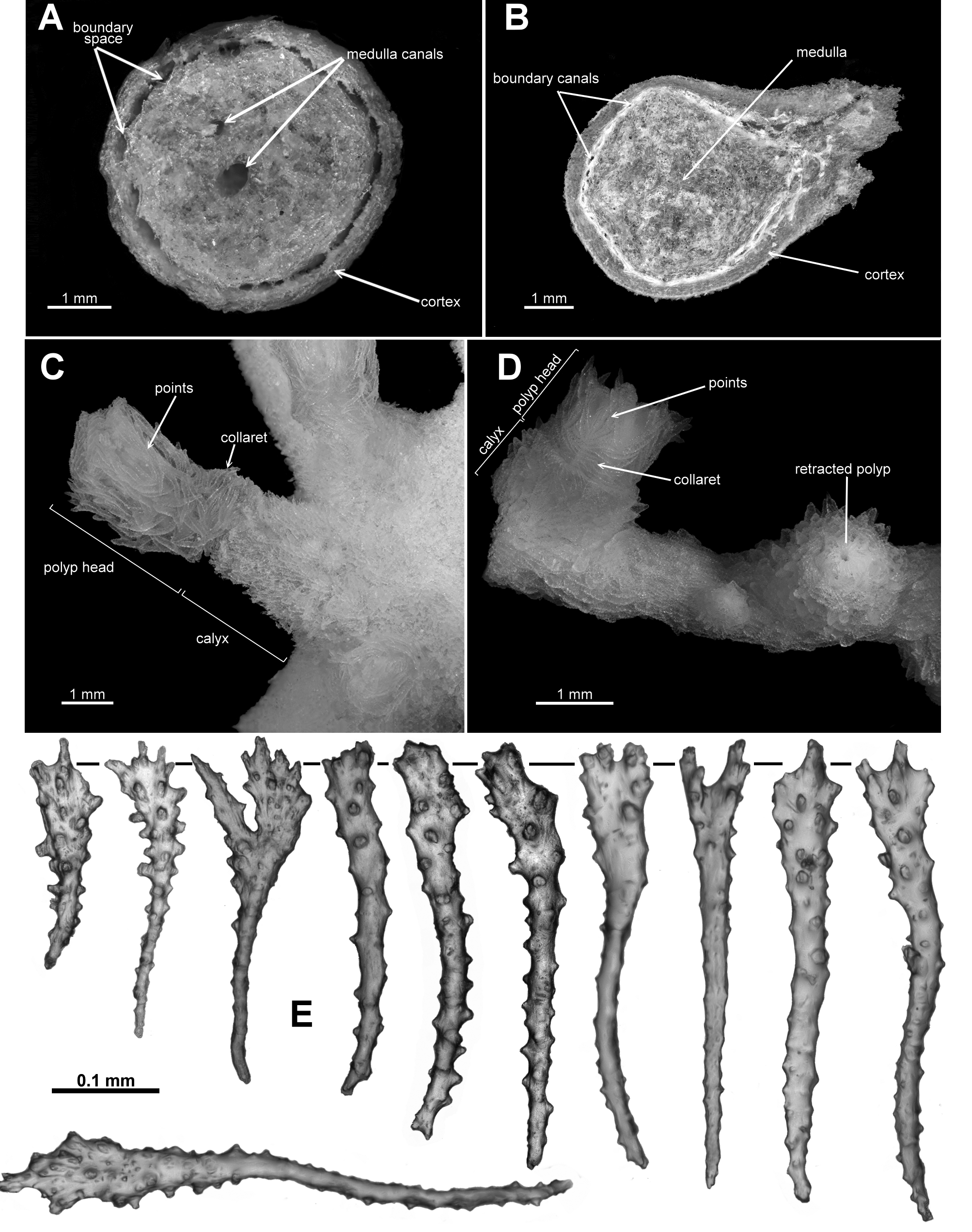
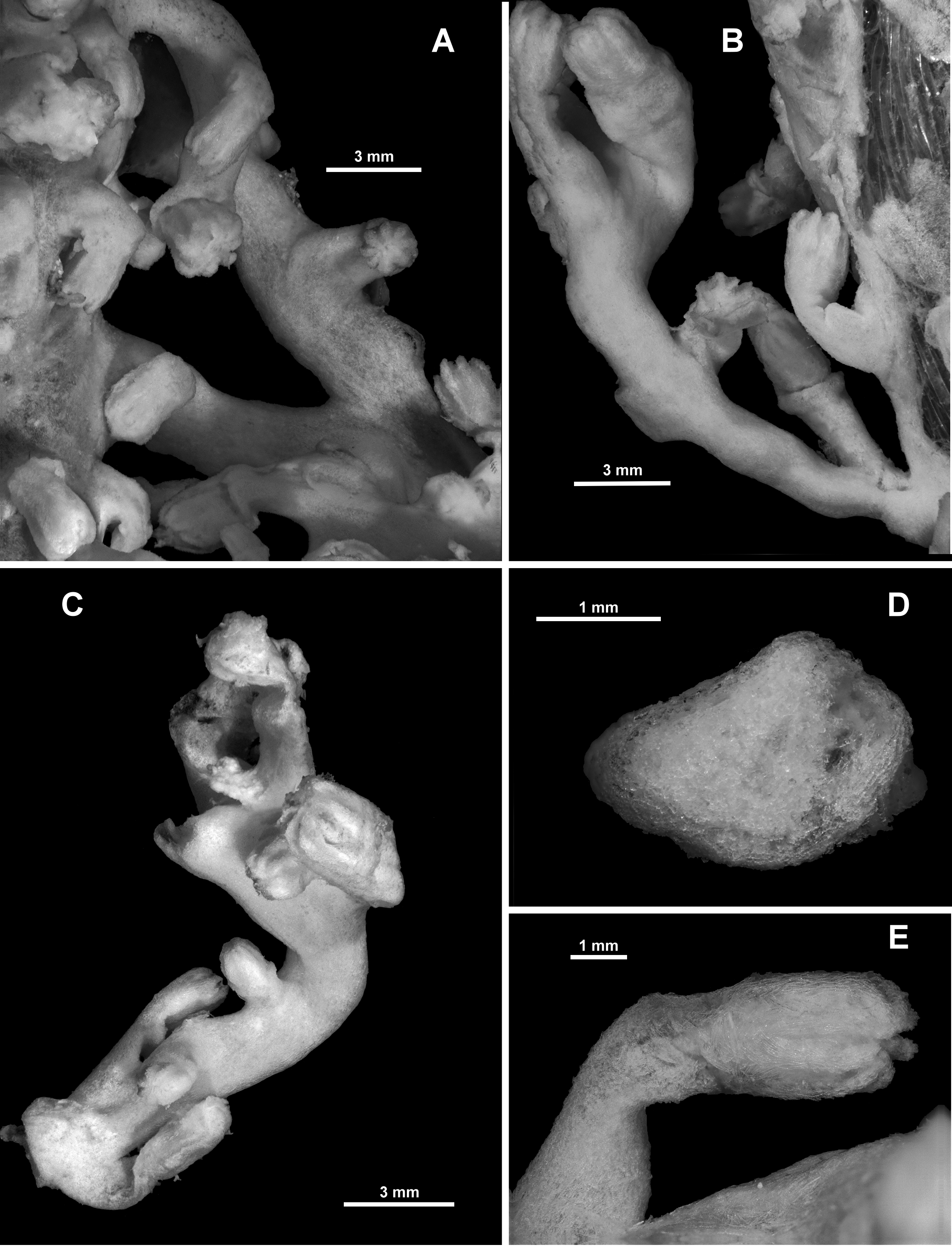
Description of RMNH Coel.33874. This specimen, collected from Cape Verde Islands in the western Atlantic Ocean, is a small colony which has encrusted a piece of green nylon rope (Stokvis & van Ofwegen 2006: Fig. 1a View FIGURE 1 ). Most of the colony is membranous and there are two distinct outgrowths where the colony has encrusted a dead hydroid, but there are also four or five instances where the colony has grown small but distinct scleraxonian branches with a number of polyps on each branch ( Fig. 68 View FIGURE 68 A–C). The branches available for dissection were all composed of a cortex with indistinct boundary canals surrounding a medulla ( Fig. 68 View FIGURE 68 D). There were no central medulla canals in the few cross-sections taken. As described by Stokvis & van Ofwegen, “the calyces are 2–6 mm high [and] the expanded polyps are 1.5–4 mm long and 1–3 mm wide”. Most polyps are partly or fully extended with the polyp neck exposed ( Fig. 68 View FIGURE 68 E). Sclerites are arranged into points and a diffuse collaret on the polyp head while sclerites on the neck are without a definite arrangement. The points consist of straight to slightly curved tuberculate sticks and spindles, ( Fig. 69 View FIGURE 69 A), up to 0.85 mm long (Stokvis & van Ofwegen 2006) which transition to tuberculate clubs (0.1–0.3 mm) and short, thick, warty rodlets (0.08–0.13 mm) along the rachis of the tentacles ( Fig. 69 View FIGURE 69 B). The warty rods are concentrated distally in the tentacle rachis and the clubs are orientated longitudinally along the tentacle. The base of the pinnules has many, longitudinally arranged, thorny josephinae clubs and smaller, lightly tuberculated rods ( Fig. 69 View FIGURE 69 C) which are 0.17–0.3 mm and 0.05–0.17 mm long respectively. The neck and the calyx have a dense covering of slender, tuberculate sticks and spindles (0.24–0.8 mm long) and short, thick, warty rodlets (0.1–0.26 mm long) ( Fig. 70 View FIGURE 70 A, B). The cortex has slender, tuberculate sticks and spindles, 0.13–0.5 mm long ( Fig. 71 View FIGURE 71 A), and the small piece of medulla sampled had mainly straight, lightly tuberculate sticks and spindles and sticks ( Fig. 71 View FIGURE 71 B), many of them broken during sampling, with some examples of fusion occurring.
Colour: The colour was recorded as “whitish” and the colony appears cream in Fig. 1a View FIGURE 1 in Stokvis & van Ofwegen 2007. The colour of the holotype was not recorded, however, a specimen from Norway (NTNU-VM 67147) was creamy peach to light apricot when alive ( Fig. 72 View FIGURE 72 C). Other specimens are recorded as being “peach” and “rose pink”.
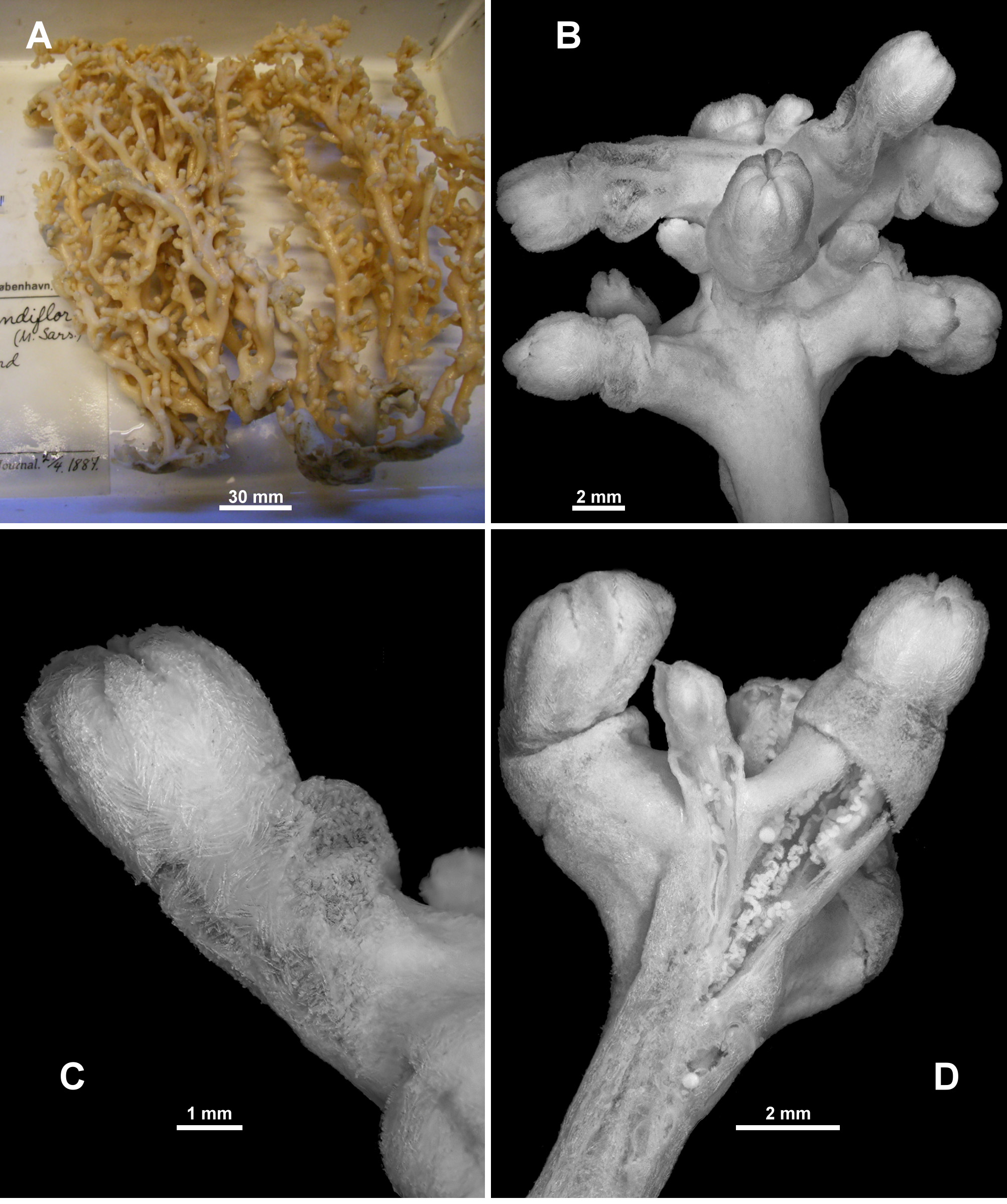
Description of NTNU-VM 63143: Colony form: The specimen consists of seven fragments of what was probably one colony ( Fig. 72 View FIGURE 72 A). The main holdfast is an encrusting, membranous part of the colony, growing over coral rubble. From this, five branches emanate basically perpendicular to the membrane and roughly parallel to each other. There are examples of anastomoses, predominantly at the bases of the branches. These upright branches are slightly flexible, but will snap if bent, are reasonably robust, a relatively consistent diameter (4–6.4 mm) and range from 46–83 mm long. Occasionally, the upright branches have secondary branching but usually only close to the distal end, and these branches are quite short, ranging from 3.3–4.6 mm only. In general, the branches range between oval to circular in cross-section, but at places where polyps occur and at points of bifurcation, the branches can be distorted.
Polyps are distributed throughout the colony, on all sides of the branches and also on the membranous part. There is a tendency for their calyces to crowd together in terminal bunches ( Fig. 72 View FIGURE 72 B). The largest distance where there are no calyces is only approximately 5 mm with most closer together.
The colony, although in a number of fragments, is in good condition with many intact polyps and much unbroken colony surface.
Colour: Colour was not recorded when alive but is now light cream in alcohol.
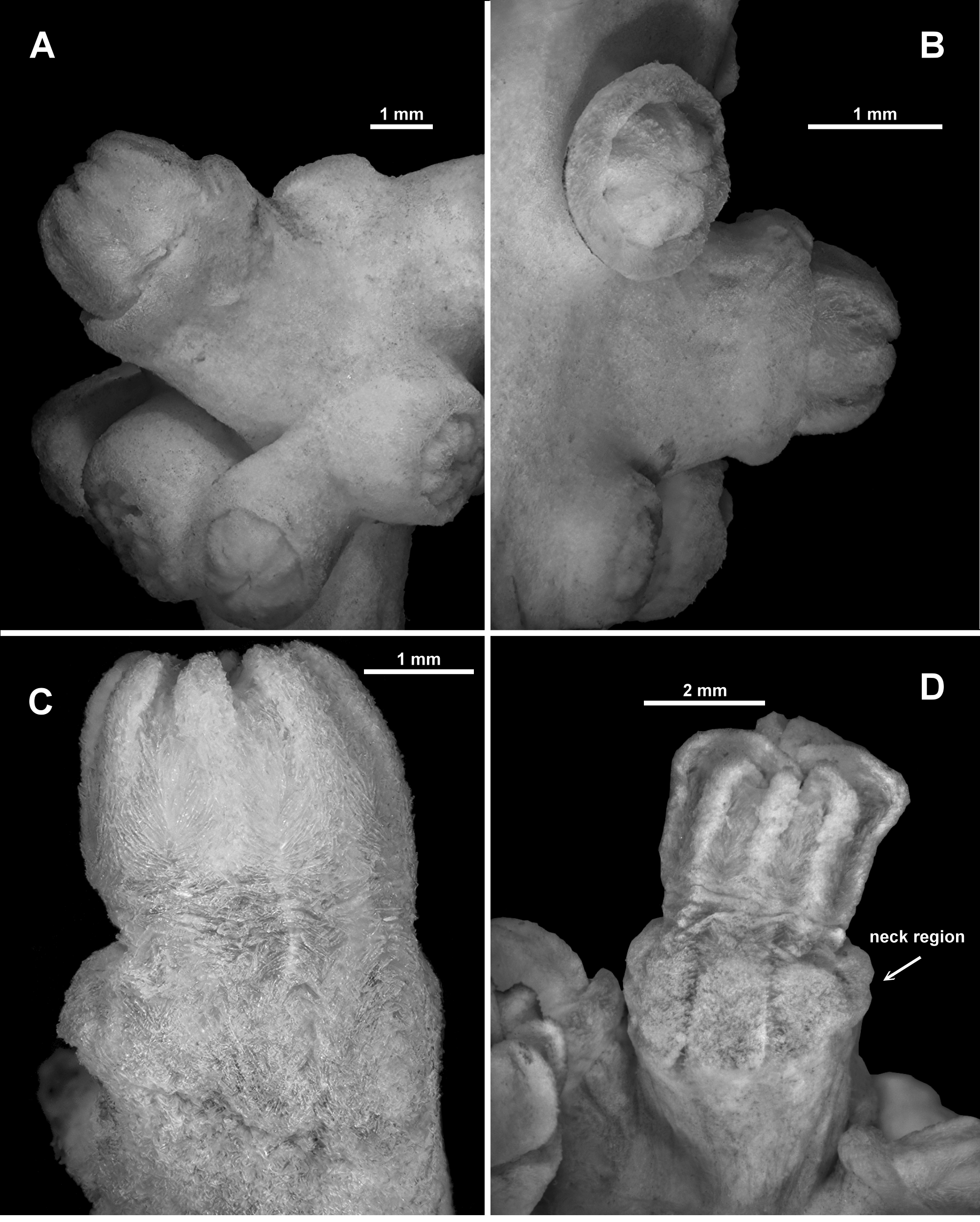
Polyps and calyces: Calyces are relatively short, flat-topped, conical projections which protrude, in general, at right angles to the branches except at branch tips where they crowd together at many angles ( Figs. 72 View FIGURE 72 B; 73A). They range from 1.5–2.5 mm tall and 2.5–3.8 mm wide. All are covered with a layer of tightly packed sclerites giving a very smooth appearance. Some of the calyces have eight minor bulges or longitudinal ridges at the lip of the calyx corresponding to the eight points on the polyp head. The majority of polyps are partly retracted such that the base of the polyp head sits on the lip of the calyx or is partly enclosed in the calyx ( Fig. 72 View FIGURE 72 B; 73B). There are examples of entirely retracted polyps where the calyx has closed over the top of the polyp head and rare examples of partially extended polyps where the polyp neck area is visible, which can be slightly swollen or ballooned out above the firm calyx ( Fig. 73 View FIGURE 73 C, D). The exsert part of the polyps can be 3–5 mm tall but most commonly just the polyp head is visible, approximately 2–2.5 mm tall. Polyp heads are approximately 2–3 mm across at the widest point. Occasionally juvenile polyps occur; these are approximately 0.8–1.3 mm tall and 1.3–1.5 mm wide. All polyps have the tentacles neatly and tightly closed within the mouth, giving the polyps a consistent starred appearance with eight rounded mounds. There is a single row of 8–10 very fleshy pinnules arranged along each side of the tentacles, some of which extend around the tip.
Medulla and Cortex: The upright branches have a medulla composed of tightly-packed, longitudinally or obliquely arranged sclerites surrounded by a thin cortex. There are many small canals between the medulla and cortex running longitudinally throughout the colony, creating a distinct boundary between the medulla and the cortex. They are clearly defined, and do not appear to anastomose ( Fig. 74 View FIGURE 74 A). There are no obvious coelenteric canals within the medulla, either at the base of the branches or at the tips. Small solenia penetrate the medulla and cortex, facilitating connections between the polyps and the main boundary canals.
Along the branches, polyp body cavities truncate evenly at the medulla while in the terminal bunches, polyp body cavities extend as gastric canals only slightly within the branches, merging into the boundary canals.
Sclerites: The calyces, polyps and colony surface are all covered in sclerites. The heads of the polyps are coated in a crowded layer, which are arranged en chevron at the points and longitudinal at the base of the tentacles ( Fig. 73 View FIGURE 73 C). There are also many intermediate sclerites between the points. The point and intermediate sclerites are mostly slightly curved sticks and spindles with a light to moderate covering of simple, often tall, tubercles ( Fig. 74 View FIGURE 74 B). Occasionally, smaller sclerites with a tendency for one of the tips to be clavate were noted but they are not common ( Fig. 74 View FIGURE 74 Ba). Lengths range from 0.19–0.65 mm but most are between 0.2–0.6 mm long. Immediately basal to the points, similar sclerites are arranged transversely to obliquely and form a diffuse collaret when the polyp is retracted.
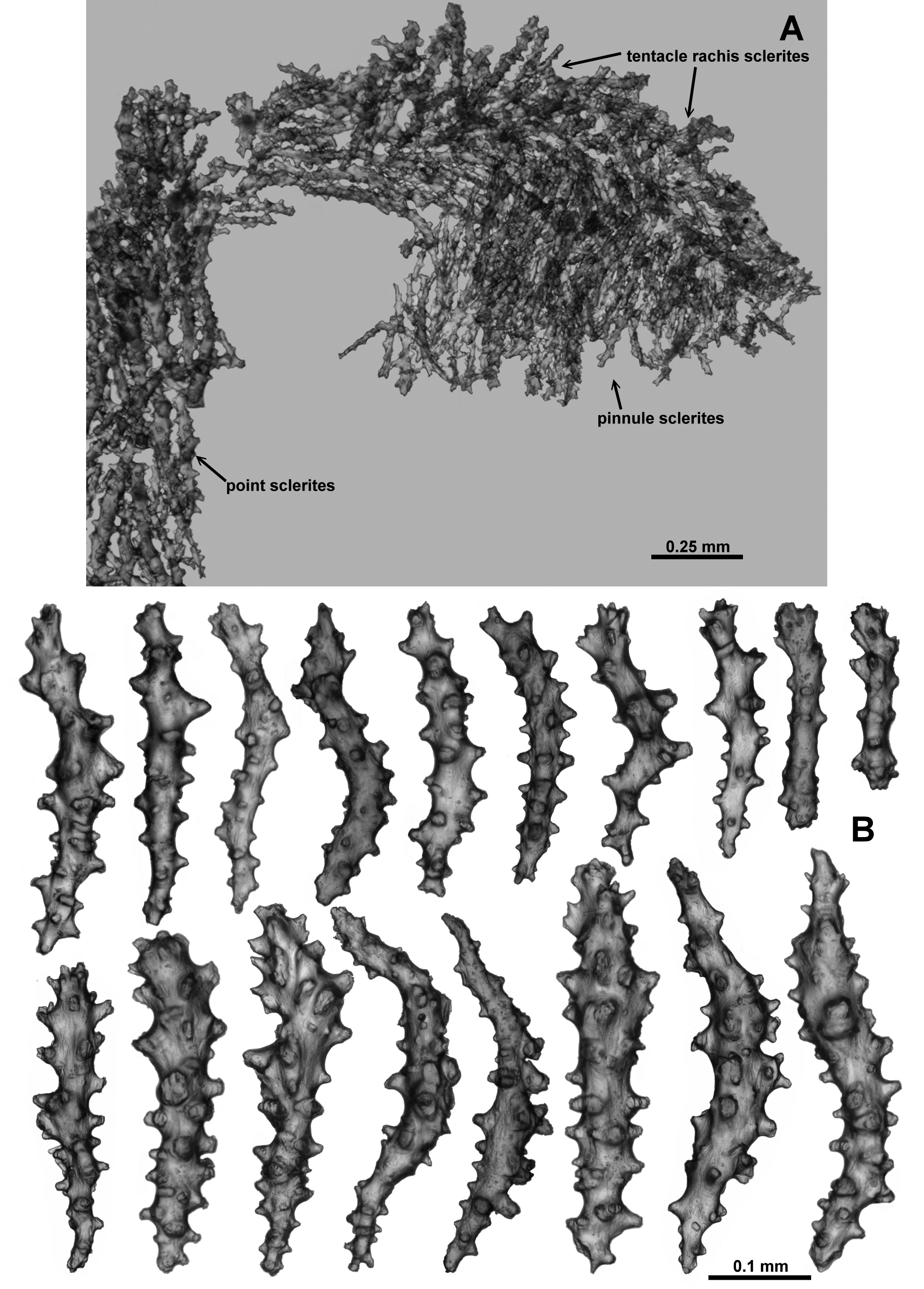
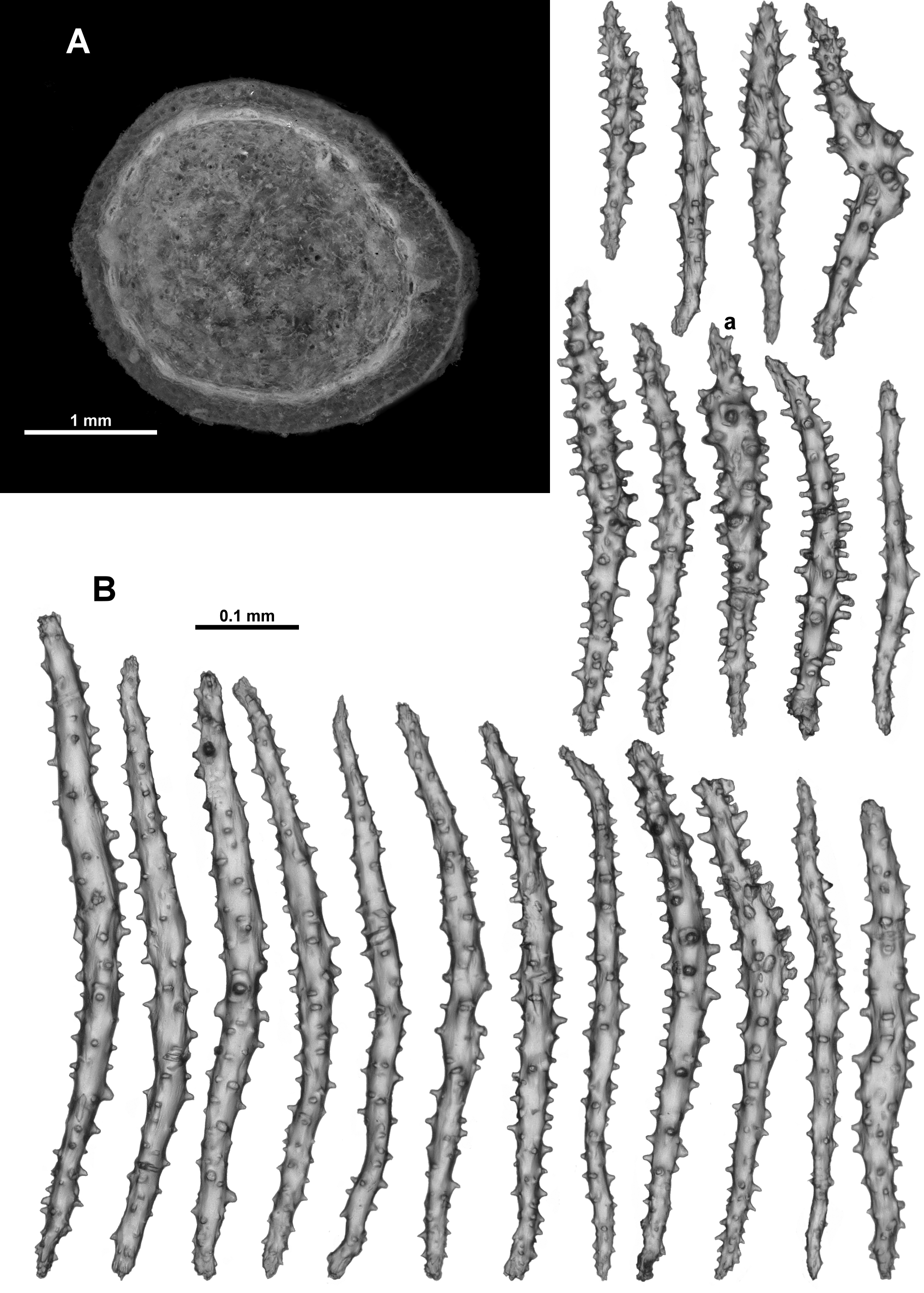
From the tip of the points, the crowded sclerites continue longitudinally along the tentacle rachis ( Fig. 75 View FIGURE 75 ). There are a mixture of forms: small clubs with flame-like spikes on the head (0.09–0.25 mm long) ( Fig. 76 View FIGURE 76 a); mostly straight, lightly tuberculate sticks and spindles (0.2–0.48 mm long) ( Fig. 76 View FIGURE 76 b) and short, straight, sparsely tuberculate flat rodlets (0.1–0.2 mm long) ( Fig. 76 View FIGURE 76 c). Most sclerites are arranged longitudinally along the tentacle rachis with any clubbed or spiny tips always arranged distad, although in the distal quarter of the tentacle the flat rods sometimes appear to lie transversely. The small clubbed sclerites ( Fig. 76 View FIGURE 76 a) are arranged along the centre of the tentacle rachis ( Fig. 75 View FIGURE 75 ) with the other types of sclerites occurring on the flanks of the tentacles. Sclerite length tends to decrease distally.
The pinnules are fleshy and relatively large with narrow sclerites crowded longitudinally ( Fig. 75 View FIGURE 75 ). However, this fleshy nature combined with few sclerites actually reaching the distal tip of the pinnules means it is possible to conclude, erroneously, that there are no sclerites in the pinnules. The sclerites include rodlets (0.1–0.2 mm long) and narrow, thorny josephinae clubs (0.2–0.4 mm long) ( Fig. 77 View FIGURE 77 ). Some of the thorny josephinae clubs are also found along the sides of the tentacles, extending into the top of the pinnules.
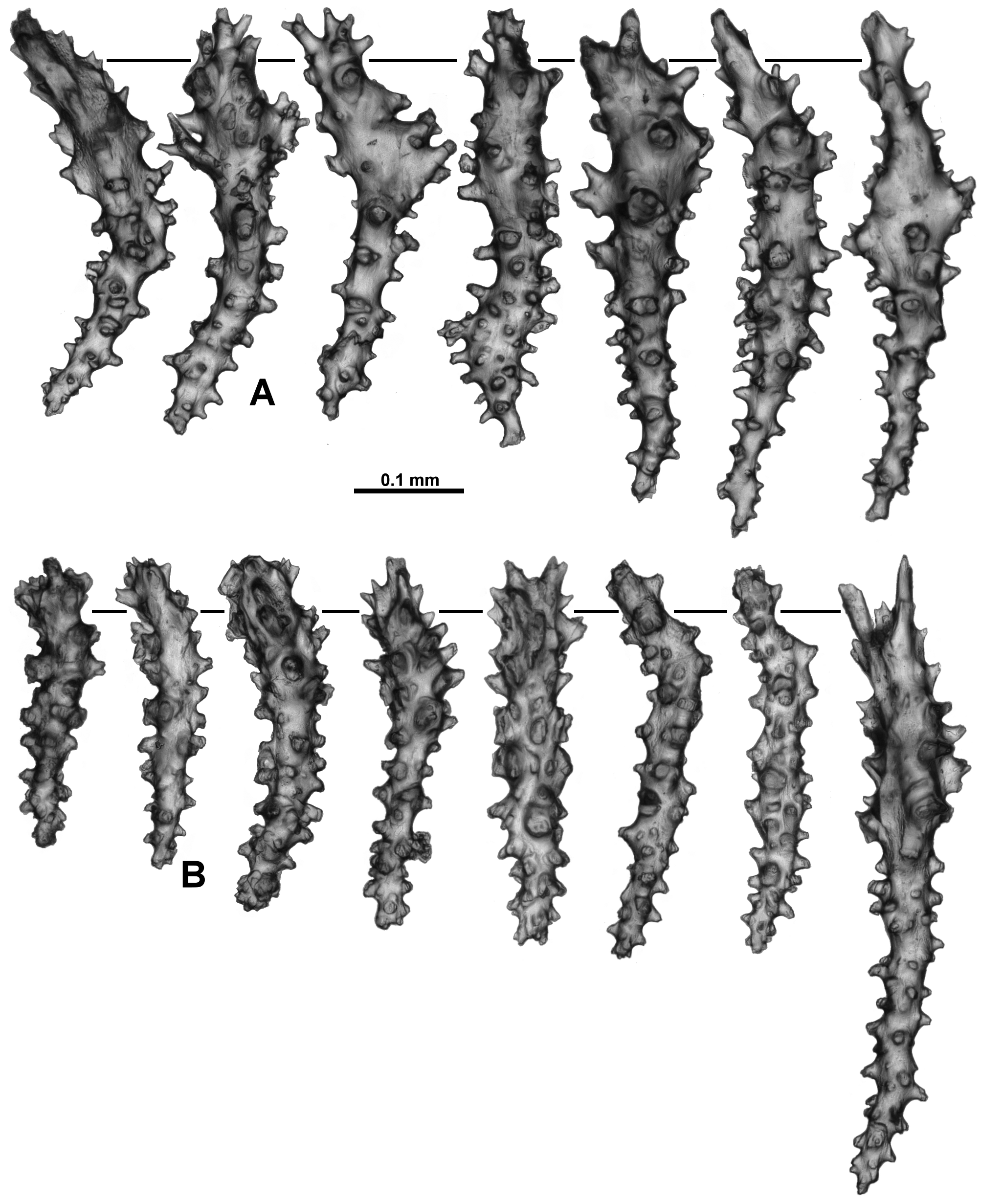
Below the polyp head and diffuse collaret, the polyp neck is covered in a crowded layer of haphazardly arranged sclerites ( Fig. 73 View FIGURE 73 C, D). The majority are short, straight, tuberculate sticks and spindles along with clubs and rodlets with relatively large warts and tubercles ( Fig. 78 View FIGURE 78 A). There are also occasionally small, tuberculate crosses, some of which are unevenly developed sticks and spindles, resembling those from the points. Lengths range from 0.2–0.45 mm for the sticks and spindles to 0.08–0.22 mm long for the small rodlets and clubs and the crosses are approximately 0.07–0.11 mm.
Numerous small sticks, spindles and rods with simple tubercles are found in the pharynx ( Fig. 78 View FIGURE 78 B). These are a relatively uniform size, with most only ranging from 0.09–0.15 mm long, although there are occasionally longer sclerites, up to 0.2 mm long. Small crosses are also occasionally found.
Calyces are covered in a dense, smooth layer of sclerites which are small enough to knit together to commonly create a mat-like appearance to the surface ( Fig. 73 View FIGURE 73 A, B), and it is difficult to see individual sclerites through a dissecting microscope. The sclerites comprise many short, thick, rodlets and clubs with relatively large tubercles and warts, and straight to slightly sinuous, tuberculate, narrow sticks and spindles ( Fig. 79 View FIGURE 79 ). The rodlets are a relatively consistent length, from 0.08–0.13 mm long while the clubs are from 0.09–0.19 mm long. Occasionally there are large spindles that can reach 0.65 mm long but most of them are approximately 0.14–0.5 mm long. Small crosses and irregularly branched forms with moderate tubercles occasionally occur.
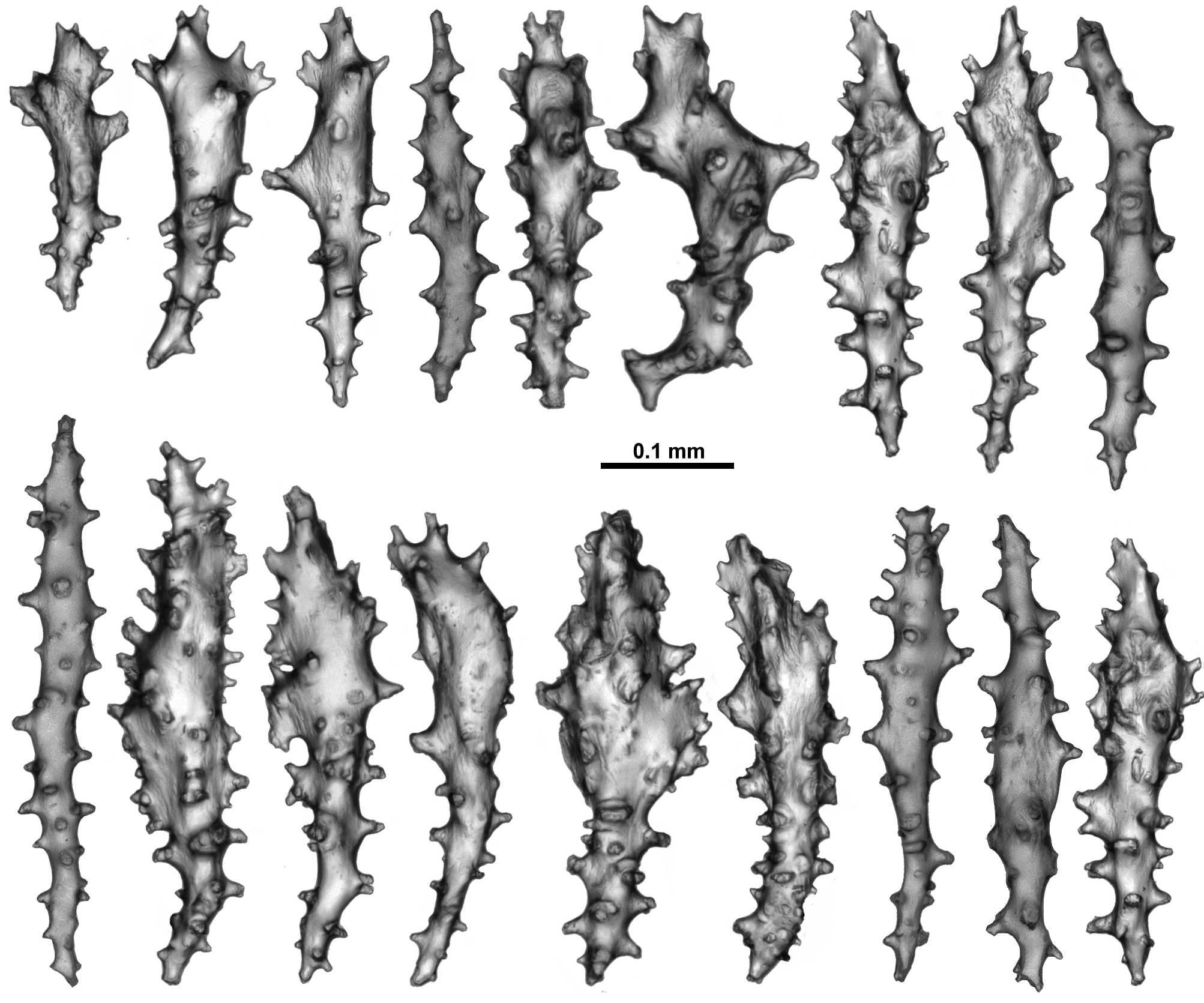
The surface of the colony has very similar sclerites to those in the calyces ( Fig. 80 View FIGURE 80 ). The short, warty rodlets (0.07–0.16 mm long) are again common as are clubs and predominately straight sticks and spindles. Most of the sticks and spindles grade from 0.16–0.35 mm long, but much larger sclerites, up to 0.7 mm, were found. Very occasionally tuberculate crosses also occur.
The medulla has narrow, slightly curved or straight, sparsely tuberculate sticks and spindles ( Fig. 81 View FIGURE 81 ). Many are brown in colour when viewed under transmitted light, and fragile so they tended to be broken during the sampling procedure. The brown sclerites were significantly more common in the centre of the medulla while the clear spindles were more likely to be from the outer part of the medulla. The clear spindles were usually from 0.09– 0.5 mm long while the fragile, brown sclerites from the centre of the medulla were estimated to be 0.12–0.62 mm long, which is probably an underestimate. Fusion and branching of sclerites were observed but these were not common.
Sclerites were all transparent under transmitted light except for some brown medulla sclerites.
Variation: Most of the specimens examined are generally consistent in colony form and sclerite shape. One of the notable exceptions in colony form and appearance is ZMUC-ANT-000468 ( Fig. 82 View FIGURE 82 A) which is a large, branching colony, with many anastomoses, the vertical branches joining and separating multiple times making it a much more complex colony than the one described in detail above. Additionally, most of the polyps are preserved exsert rather than retracted ( Fig. 82 View FIGURE 82 B), exposing the neck region so that the sclerite arrangement can be more easily assessed ( Fig. 82 View FIGURE 82 C). A longitudinal cross-section of a terminal polyp bunch ( Fig. 82 View FIGURE 82 D) shows the polyp body cavities do not extend very far into the medulla, that there are no obvious large canals, and that the cortex is firmly attached with only small boundary canals. The sclerites of this specimen are similar to those of the specimen described above but are larger and slightly more warty, particularly in the cortex where they include large, warty sticks and spindles.
Similarly, ZMUB 60246 is a large, robust colony. It consists of many pieces, but also has common anastomoses and most polyps exsert ( Fig. 83 View FIGURE 83 A). A small portion of the colony is peachy pink in colour ( Fig. 83 View FIGURE 83 B) while most is cream. Sclerites are similar to NTNU-VM 63143 with the exception of rare branched, warty sticks and spindles in the points. Most of the polyps are also exsert in the specimen ZMUB 3897 which again means the colony looks quite different to NTNU-VM 63143, but the usual colony form of multiple, vertical, anastomosing branches emanating from an encrusting membrane is still evident ( Fig. 83 View FIGURE 83 C). This specimen has some sclerites from the points, calyx and surface which are simple spindles with thorns rather than tubercles as in NTNU-VM 63143. Fragments of both L. grandiflora n. comb. and Anthothela grandiflora colonies were found in Lot ZMUB 17759, with both colonies looking very similar ( Fig. 83 View FIGURE 83 D). Another specimen, ZMUB 548 ( Fig. 83 View FIGURE 83 E), is very similar in form to NTNU-VM 63143 but other colonies have much more substantial anastomoses and membranous growths making the colonies complex ( Fig. 83 View FIGURE 83 F, G).
As mentioned above, in most of the examined specimens the surface of the colony between and on the calyces is a smooth mat of sclerites with most sclerites too small to be reliably discerned under a dissecting microscope. However, in the specimen ZMUB 12120 some calyces have obvious, long, longitudinally aligned spindles in an outer layer, while the short, warty rodlets are common in an inner layer ( Fig. 84 View FIGURE 84 A, B). This arrangement does not appear to be consistent, with some calyces having only a few long spindles obvious while others have only patches where long spindles are the dominant sclerite. As the polyp neck region invariably has a jumble of smaller sticks, spindles and rods, those polyps where long spindles dominate in the calyx show a clear demarcation between the calyx and polyp neck. There are also areas of the cortex between the calyces that have long spindles, but these again are not consistent. An unregistered ZMUB specimen has a similarly patchy distribution of long spindles although it appears to have the spindles in an inner layer of the calyx with the shorter sclerites forming an outer layer.
Specimens from Trondheimsfjord, Norway, recently obtained very close to the collection site of NTNU-VM 63143, were photographed soon after collection ( Fig. 85 View FIGURE 85 A, B). Their live colour varied from creamy pink to apricot, similar to those from the Gulf of Mexico. One of these samples was preserved with many polyps having the tentacles expanded ( Fig. 85 View FIGURE 85 C, D). An additional colony from Trondheimsfjord, photographed in situ ( Fig. 86 View FIGURE 86 A, B), appears likely to be an example of this species but was not available for examination, so this must remain unconfirmed.
A small piece of a colony collected from the east coast of Brazil (NHMUK 1917.6.7.1) has sclerites consistent with those from NTNU-VM 63143 and provides more evidence that the species is widespread.
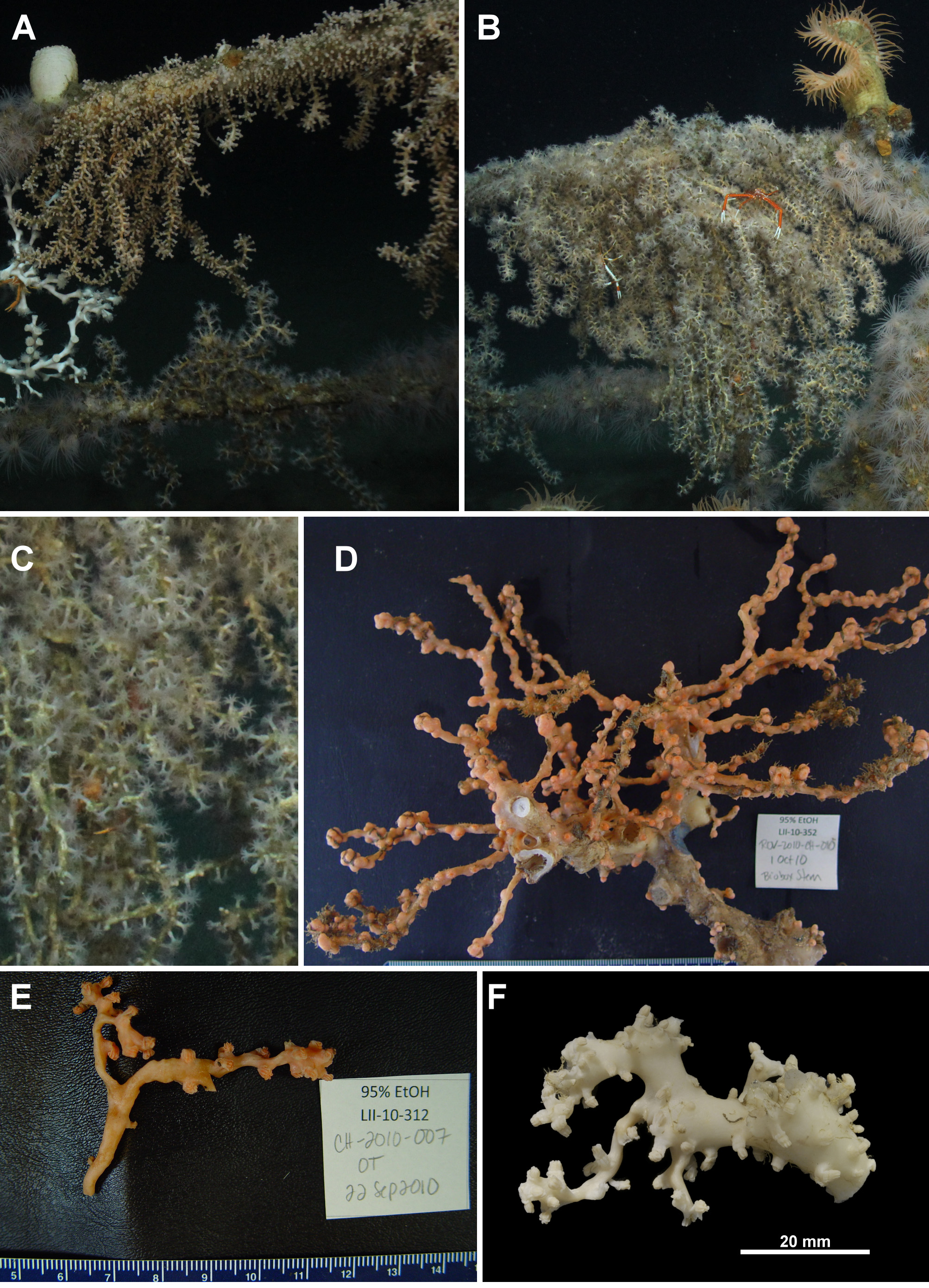
In situ photographs of the specimens TMAG K4272 and USNM 1207953 from the Gulf of Mexico show extensive colonies hanging down from artificial structures, with many branches emanating from a membranous base ( Fig. 87 View FIGURE 87 A, B). The polyps are crowded and tall with large tentacles ( Fig. 87 View FIGURE 87 C). Colonies are cream to beige in situ but are closer to apricot in bright surface light soon after collection ( Fig. 87 View FIGURE 87 D, E) but are bleached to almost white in ethanol ( Fig. 87 View FIGURE 87 F). Sclerites are similar to those of NTNU-VM 63143 in most areas ( Figs. 88 View FIGURE 88 ; 89), although the spindles from the calyx are slightly shorter (0.2–0.36 mm long in TMAG K4272 cf. 0.14–0.5 mm from NTNU-VM 63143) and sclerites from the cortex are generally stouter and have wider tubercles than NTNU- VM 63143 ( Fig. 89 View FIGURE 89 B). Considering the geographic distance separating the Gulf of Mexico from Norway and that there are genetic character differences between these geographic groups, the Gulf of Mexico specimens may represent a separate species within this genus. However, without more significant morphological differences and a stronger understanding of what is sufficient genetic variation within this group they remain within this species.
Distribution: Northern Atlantic—relatively common along the coast of Norway; Iceland; east coast of Brazil; Gulf of Mexico.
Depth: 50–550 metres.
Remarks: The description of Parerythropodium grandiflorum in Tixier-Durivault & d’Hondt (1974) is unfortunately brief and the sclerite depictions a little stylised. Given this and that the type specimen appears to be membranous it is perhaps understandable that this species has not before been associated with the much more substantial and structurally complex scleraxonian specimens included here. The taxonomic confusion around Anthothela has also contributed to specimens which were actually L. grandiflora n. comb. remaining unrecognised.
For over 150 years this species has been repeatedly mistaken for specimens of Anthothela grandiflora . This is understandable as the two species are sympatric, similar in colour, similar in colony form, and many of the sclerites in both species are tuberculate sticks and spindles. The initial mistake, made possibly by Sars not long after he described A. grandiflora , seems to have been compounded by Broch (1912b) in his influential re-description of A. grandiflora where he figured what appear to be the small, warty rodlets from the calyx and cortex common in L. grandiflora n. comb. Although stating that he viewed many specimens from a number of museums, including the holotype of A. grandiflora , he was not specific on exactly which specimen or specimens he figured. In his discussion he mentioned short, thick cylinders, only 0.1–0.2 mm long, as abundant in the calyces and in the colony surface ( Broch 1912b). The holotype of A. grandiflora does not have sclerites like that but as they were figured in Broch’s re-description the misconception that it does has continued for many years. Additionally, Broch mentioned that there are no sclerites in the pinnules of A. grandiflora ; this is clearly incorrect, as in the holotype of A. grandiflora the pinnules are packed with long spatulate clubs; however the sclerites in the pinnules of L. grandiflora n. comb. tend to be masked by the fleshiness of the pinnules and do not always reach the tip of the pinnule so it is feasible that Broch failed to notice them. The obvious hypothesis is that Broch used more than one specimen in his description and at least one of these specimens was L. grandiflora n. comb. This indeed seems to be the case, as sample lots labelled A. grandiflora that were investigated for this project (NTNU-VM 40341, ZMUB 17759, ZMB 5527 (determined by Broch)), had both species found together in the same jar. Subsequent researchers often used Broch’s 1912b re-description, sometimes finding the short, warty rodlets (so possibly actually investigating a specimen of L. grandiflora n. comb.) and sometimes failing to find them (so possibly with a true A. grandiflora specimen). For example, Verrill (1922) in his description of A. grandiflora stated the figures are “from the type described in 1869”, however Plate VI Fig. 1 View FIGURE 1 appears to depict the sclerites of L. grandiflora n. comb. with the small, warty rodlets from the calyx and cortex. His description—”the cortex of the calicles and coenenchyme is finely granulous under a lens. when dried, and the surface is filled with an abundance of very irregular and pop-corn shaped spicules, with roughly warted and mostly spindle-shaped spicules beneath, mixed with some irregular clubs, rods, and many small irregular forms of various shapes” ( Verrill 1922) is a good description of the sclerites found in the calices and cortex of L. grandiflora n. comb. instead of the holotype of A. grandiflora which has calyx and cortex sclerites that are chiefly long, narrow sticks and spindles. However, the colony depicted in Verrill’s Text Fig. 2 View FIGURE 2 is much like the holotype of A. grandiflora and not of L. grandiflora n. comb.
Stiasny (1937) mentioned two colony forms in the specimens of Anthothela grandiflora he investigated. The first form had a flat membranous plate from which robust, upright branches with little or no anastomoses arise, much like that described here for L. grandiflora n. comb. The description of the colony form of the second specimen more closely matches that of A. grandiflora . The text figure of the sclerites may easily be depicting the warty rodlets from the calyces and cortex of L. grandiflora n. comb. (his Text Fig. 1 View FIGURE 1 g–i). It is possible that the two colony forms mentioned by Stiasny actually represented the two confused species.
Considering that in every one of the six European museums visited for this study, and in collections obtained from NMNH and NHMUK, multiple, large specimens of this species were found, with most labelled as A. grandiflora , it seems highly likely there are more examples of this species still incorrectly identified as A. grandiflora in many other museums. Given that large colonies have been found in the Gulf of Mexico and noted off the coast east coast of the USA (Scott France, pers. comm.), combined with the many examples examined from Norway and the slender evidence of a presence off the east coast of Brazil, it would seem to have a widespread distribution in the Atlantic Ocean.


























| RMNH |
National Museum of Natural History, Naturalis |
| NTNU-VM |
Norwegian University of Science and Technology - University Museum |
| ZMUB |
Museum of Zoology at the University of Bergen, Vertebrate collections |
| NHM |
University of Nottingham |
| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
| NHMUK |
Natural History Museum, London |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| USGS |
U.S. Geological Survey |
| TMAG |
Tasmanian Museum and Art Gallery |
| ROV |
Museo Civico di Rovereto |
| NOAA |
National Oceanic and Atmospeheric Administration |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lateothela grandiflora ( Tixier-Durivault & d'Hondt, 1974 )
| Moore, Kirrily M., Alderslade, Philip & Miller, Karen J. 2017 |
Parerythropodium grandiflorum Tixier-Durivault & d’Hondt, 1974 : 1373
| Weinberg 1977: 148 |
| Tixier-Durivault 1974: 1373 |