
Lepidocyrtinus iegoi, Cipola & Morais & Bellini, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4898.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3F66CE1D-1036-4226-BC28-029F9961E069 |
|
DOI |
https://doi.org/10.5281/zenodo.4421088 |
|
persistent identifier |
https://treatment.plazi.org/id/7B6687A7-FFB5-FFE9-FF12-D089FC2CF8B9 |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidocyrtinus iegoi |
| status |
sp. nov. |
Lepidocyrtinus iegoi View in CoL sp. nov. Cipola & Bellini
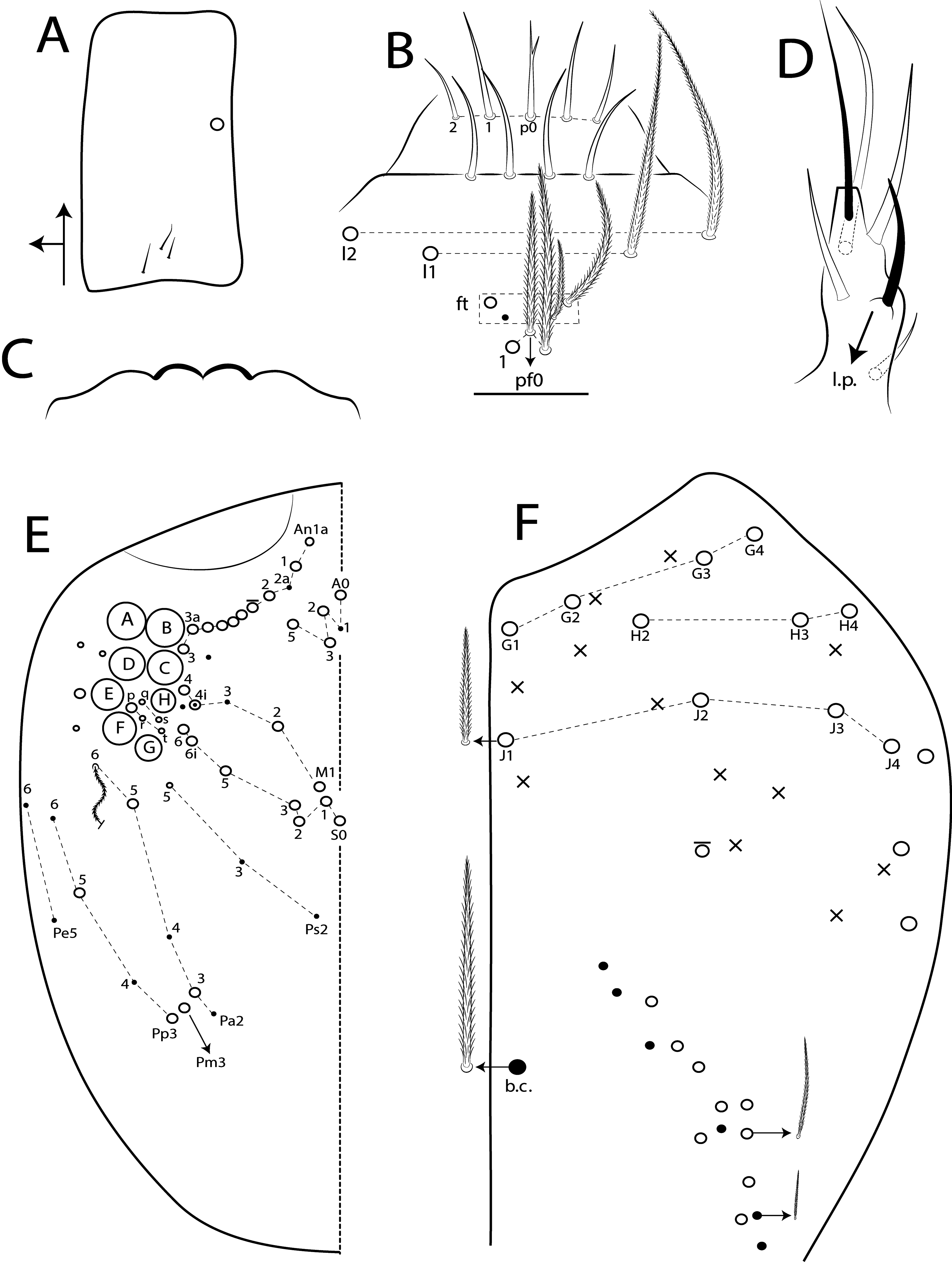
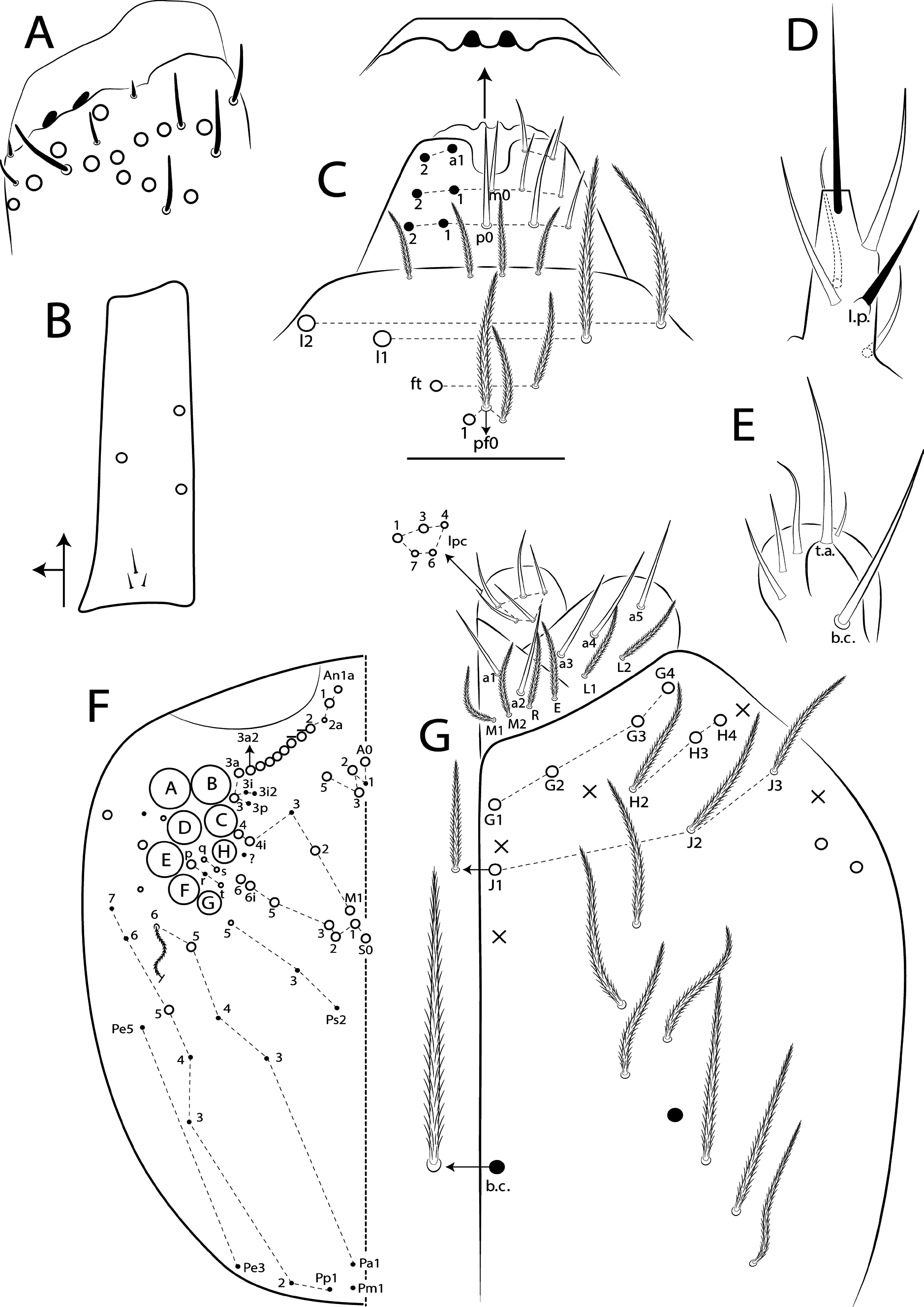
Figs 55 View FIGURE 55 , 77–80 View FIGURE 77 View FIGURE 78 View FIGURE 79 View FIGURE 80 , Tables 1 View TABLE 1 and 5 View TABLE 5
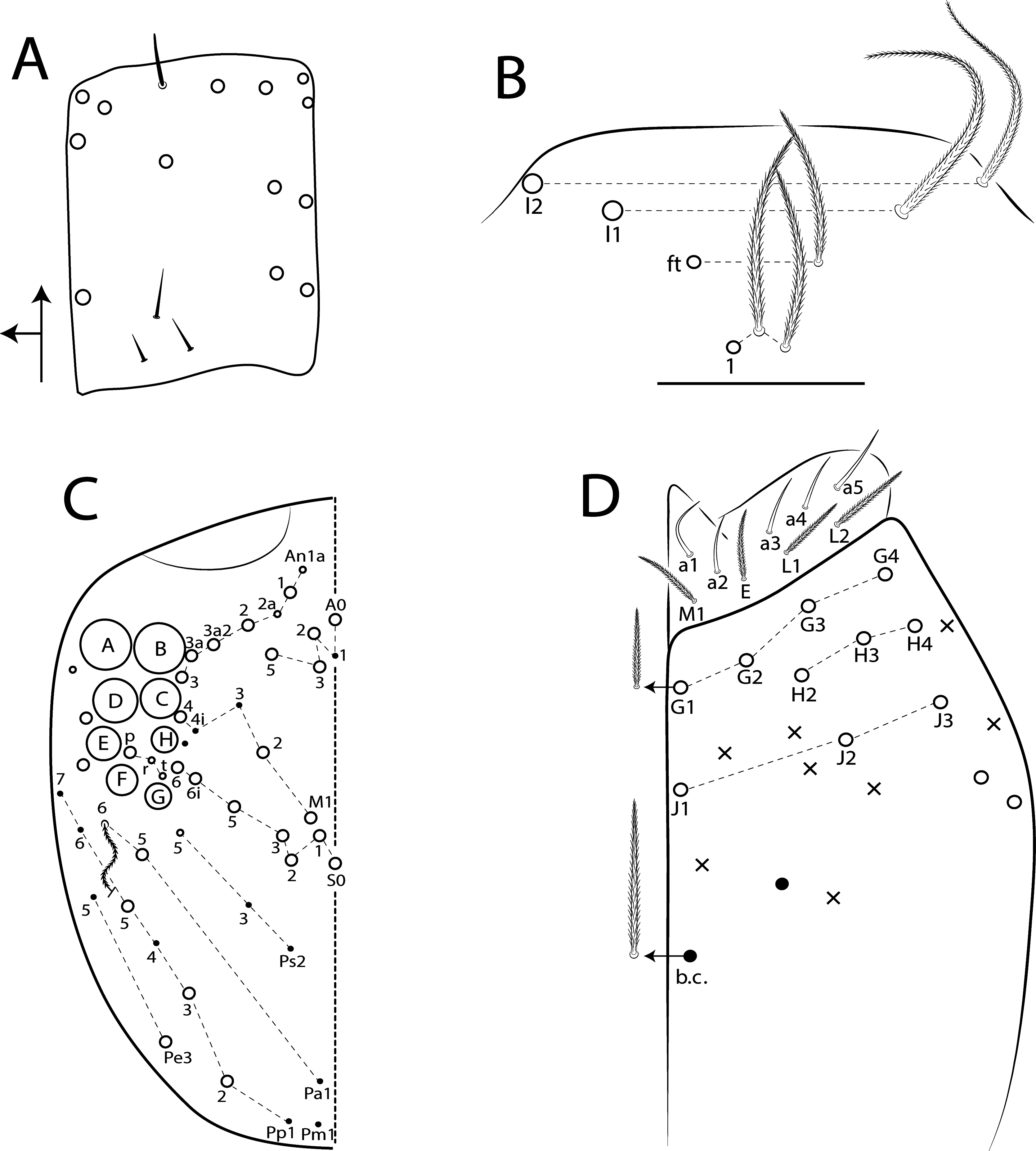
Type Material. Holotype female on slide (54107/ CAS): Madagascar, Toamasina, Moramanga municipality, Andasibe-Mantadia National Park, 18°45’39”S, 048°22’17”E ( Fig. 55 View FIGURE 55 ), Rainforest , 968 m, 23-28.xi.2012, Malaise trap, BL Fisher coll. 1 paratype female on slide (54107/ CAS), same data as holotype. GoogleMaps
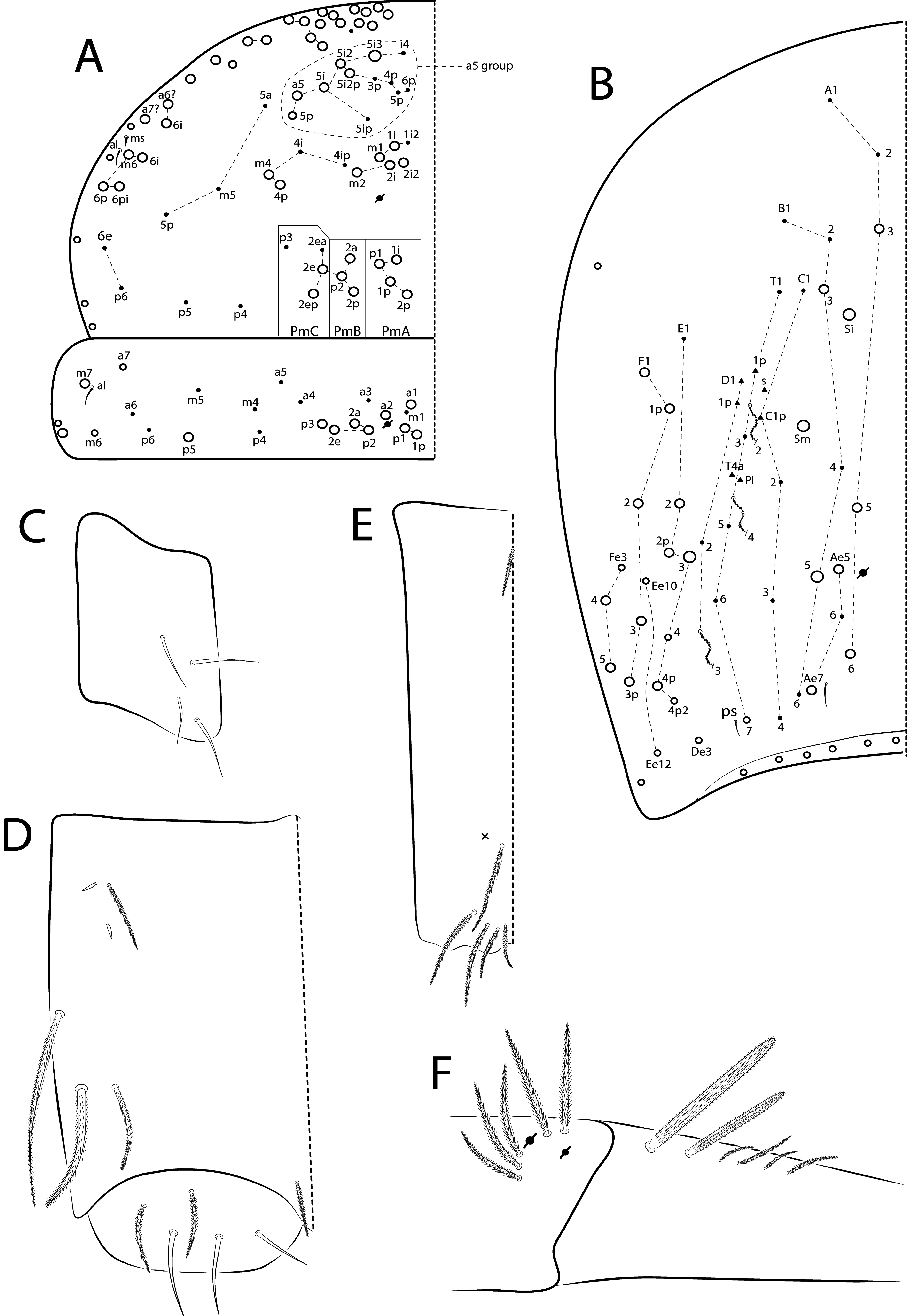
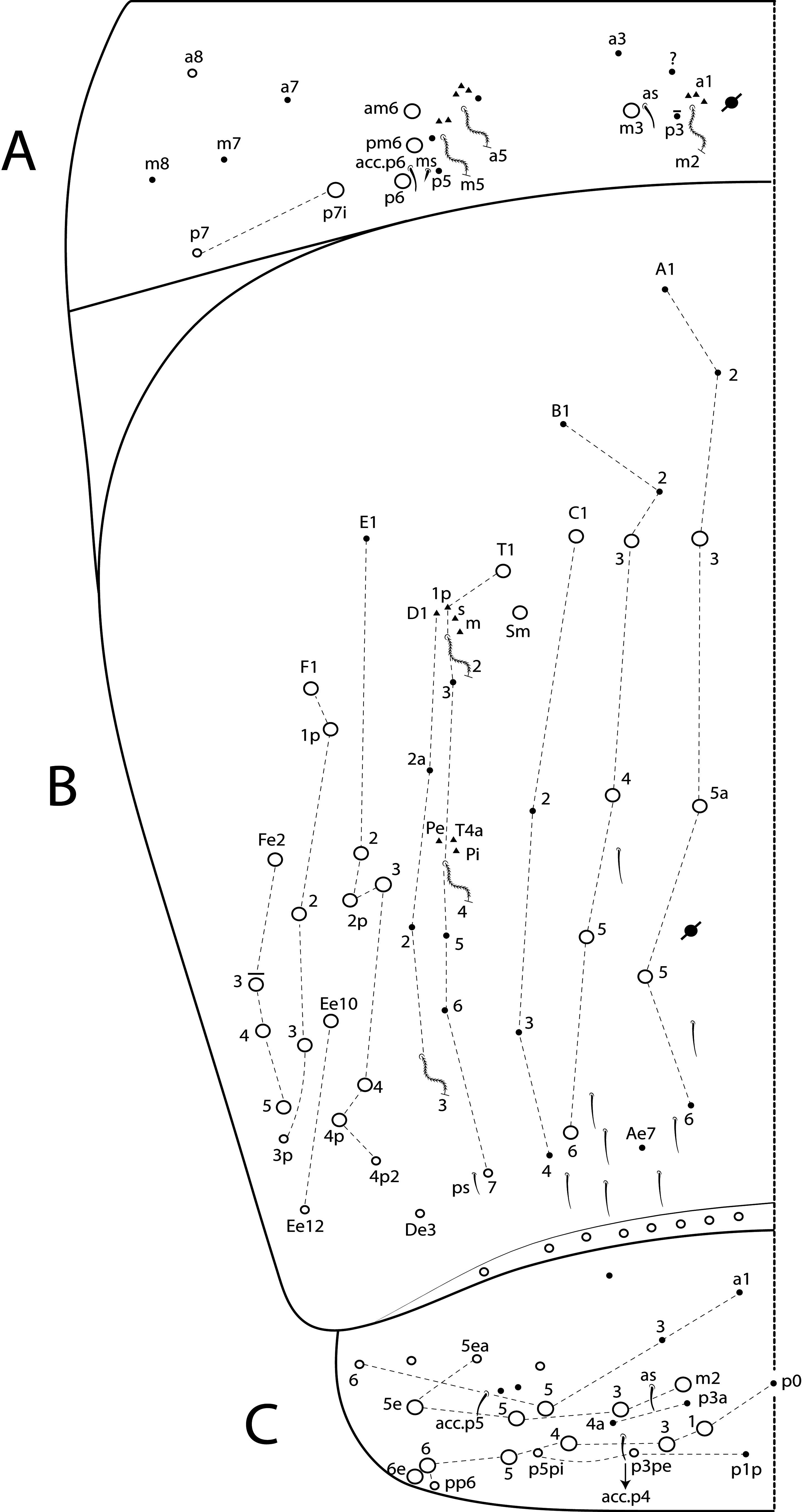
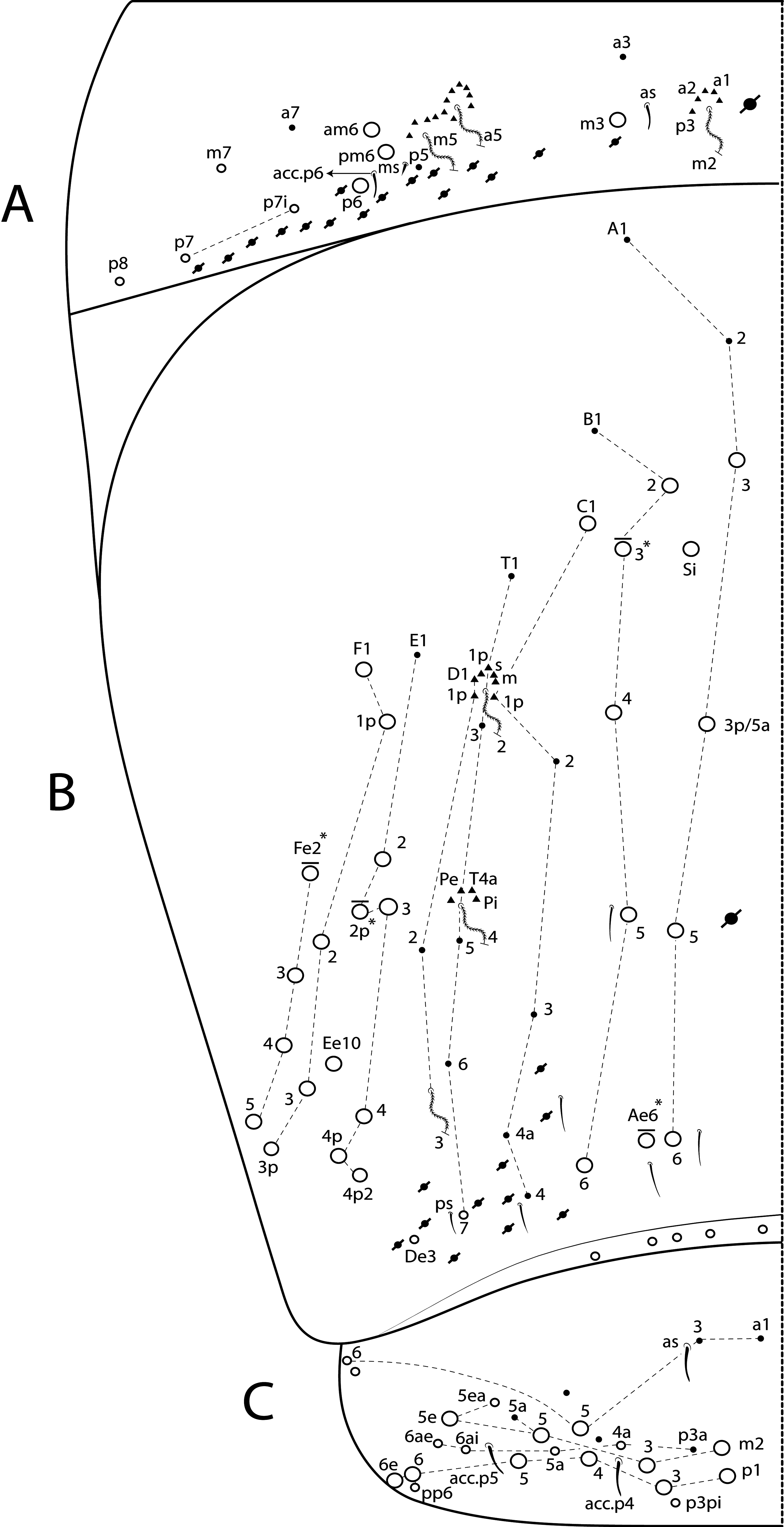
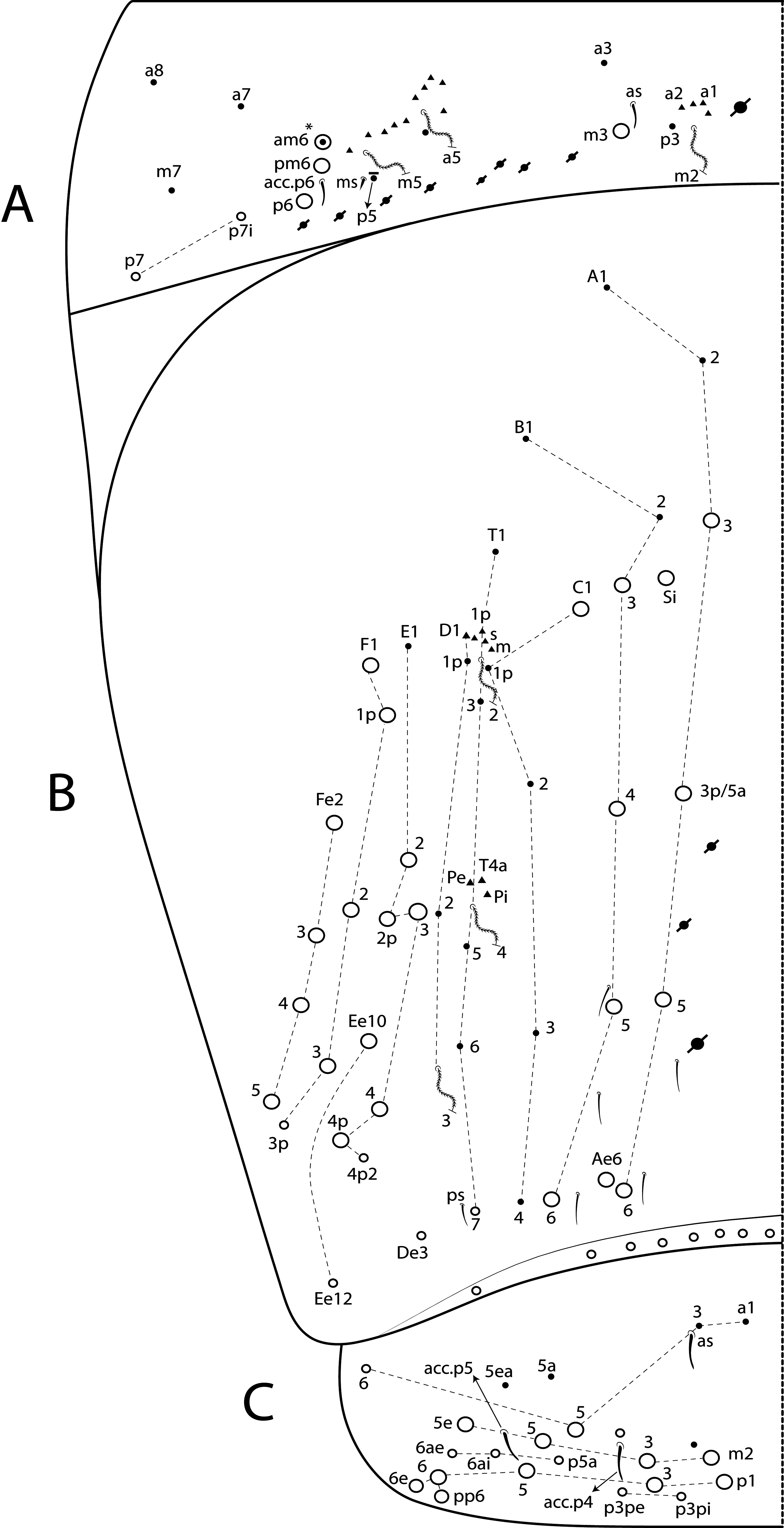
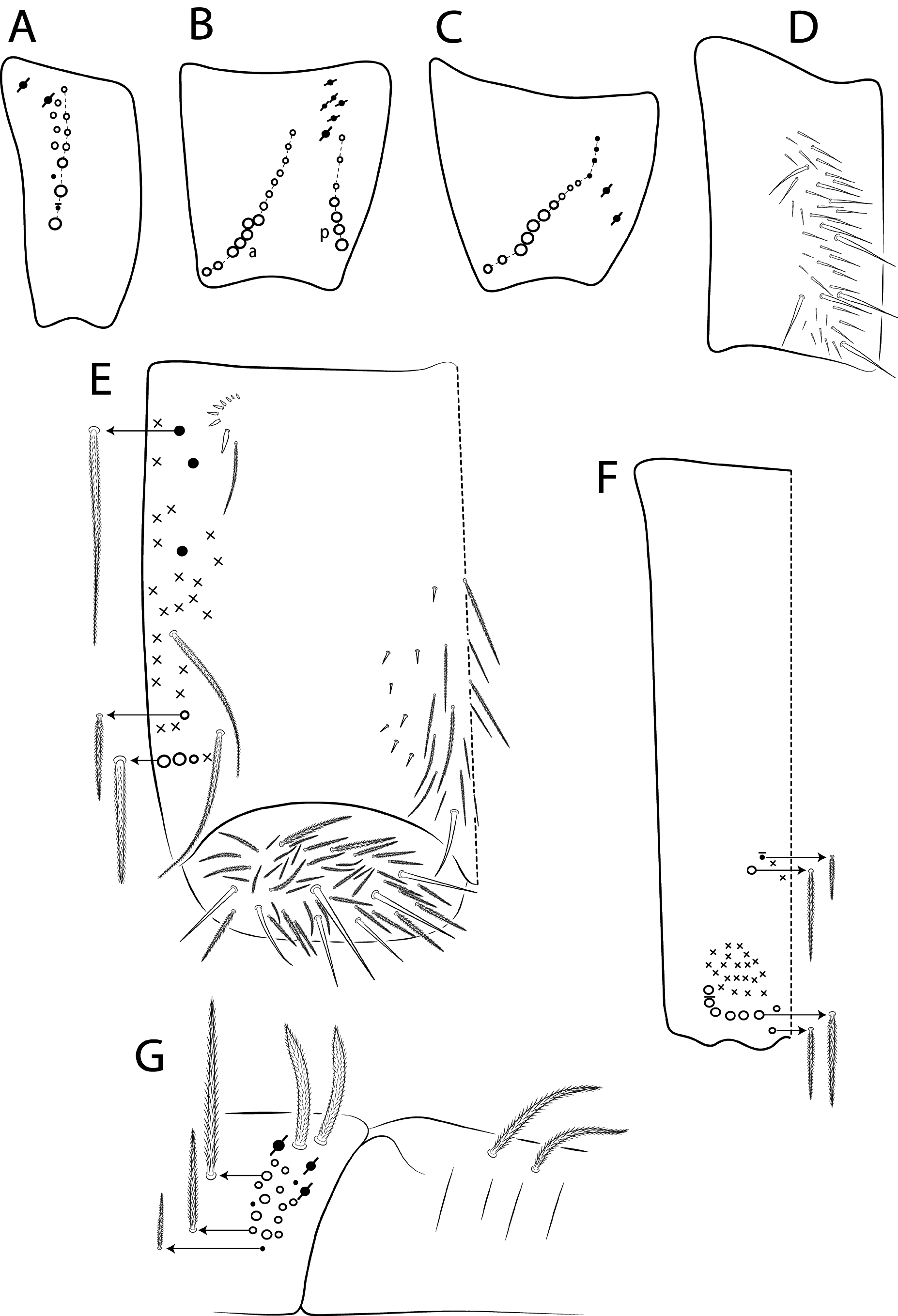
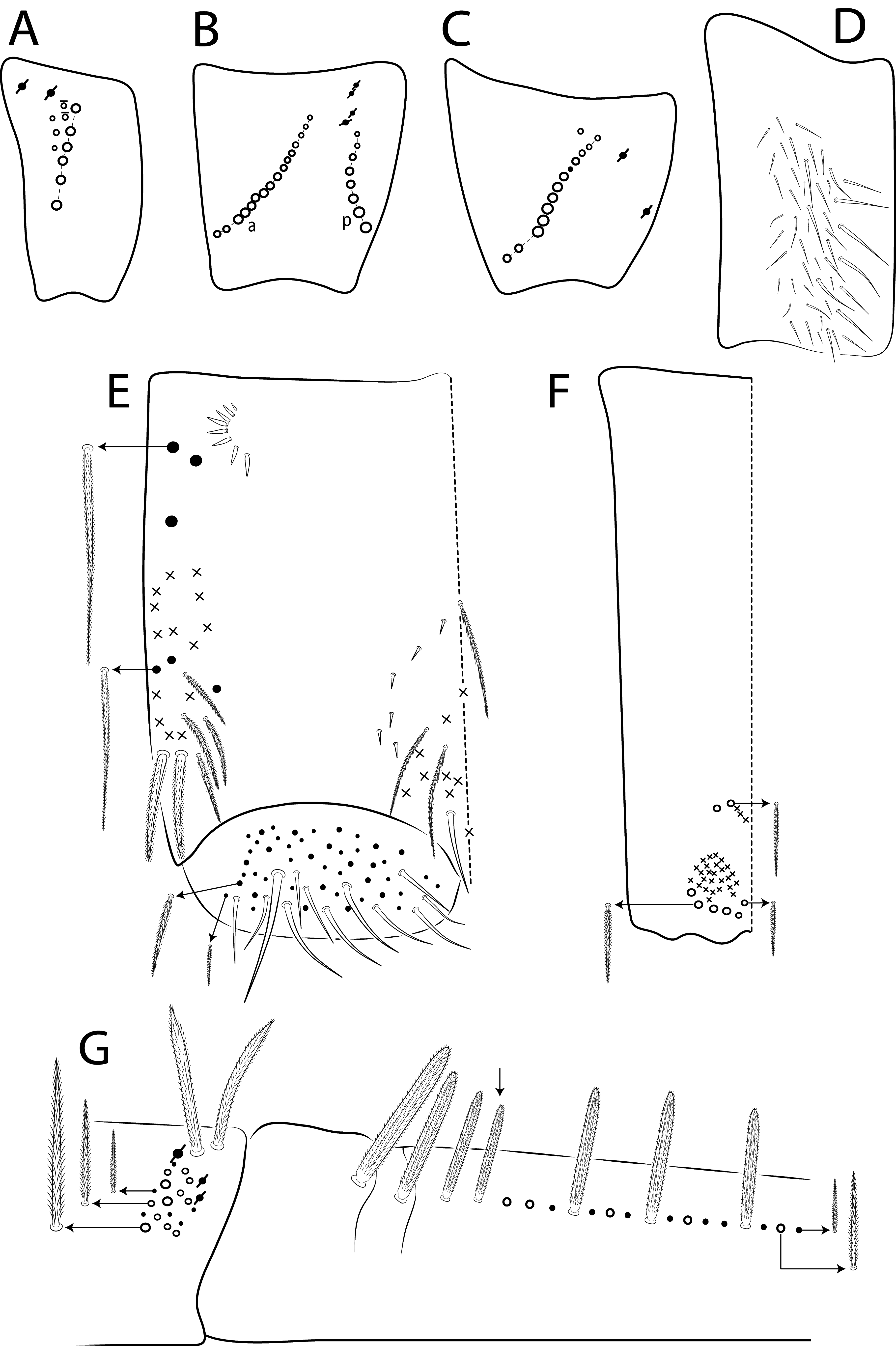
Diagnosis. Body pale ( Fig. 77 View FIGURE 77 ); head mac A0, A2–3, A5, M1–2, S0–3 and S5–6 present; prelabral chaetae smooth; labial papilla E with l.p. conical and reduced ( Figs 78 View FIGURE 78 B–C); basolateral and basomedian labial fields with chaetae R (smaller than the others) and L1–2 ciliated, e smooth; Th II projected anteriorly and with 7 anterior, 8 median (m1ip, m4–4p present, m4i mac absent) and 10–11 posterior mac (p2ep absent, p3 as mac); Th III–Abd II with 11, 5, 4 central mac respectively; Abd IV with 10–12 central mac and 14–16 lateral mac ( Figs 79 View FIGURE 79 and as in 64B); unguis a.t. absent; unguiculus pe lamella smooth and with a small proximal tooth; collophore anteriorly with 8 spine-like chaetae and 6 ciliated chaetae apically acuminate, posteriorly with 6 spines, smooth chaeta present ( Fig. 80E View FIGURE 80 ); manubrium ventrally with 0/4 subapical chaetae; manubrial plate with 2 blunt mac abruptly acuminate at the apex; dens with 4–7 blunt mac apically bullet-like ( Figs 80 View FIGURE 80 F–G).
Description. Total length (head + trunk) of specimens 2.97–3.61 (n=2), holotype 3.61 mm. Specimens pale white with vestigial dark blue pigment on distal and proximal Ant IV; eyepatches black ( Fig. 77 View FIGURE 77 ). Scales present on Ant I to proximal one fifth of Ant IV, dorsal and ventral head, dorsal thorax and abdomen, legs (except empodia), anterior and posterior collophore, ventral and dorsal manubrium and dens ventrally.
Head. Antennae subequal to trunk length ( Fig. 77 View FIGURE 77 ); ratio antennae: trunk = 1: 1.07; antennal segments ratio as
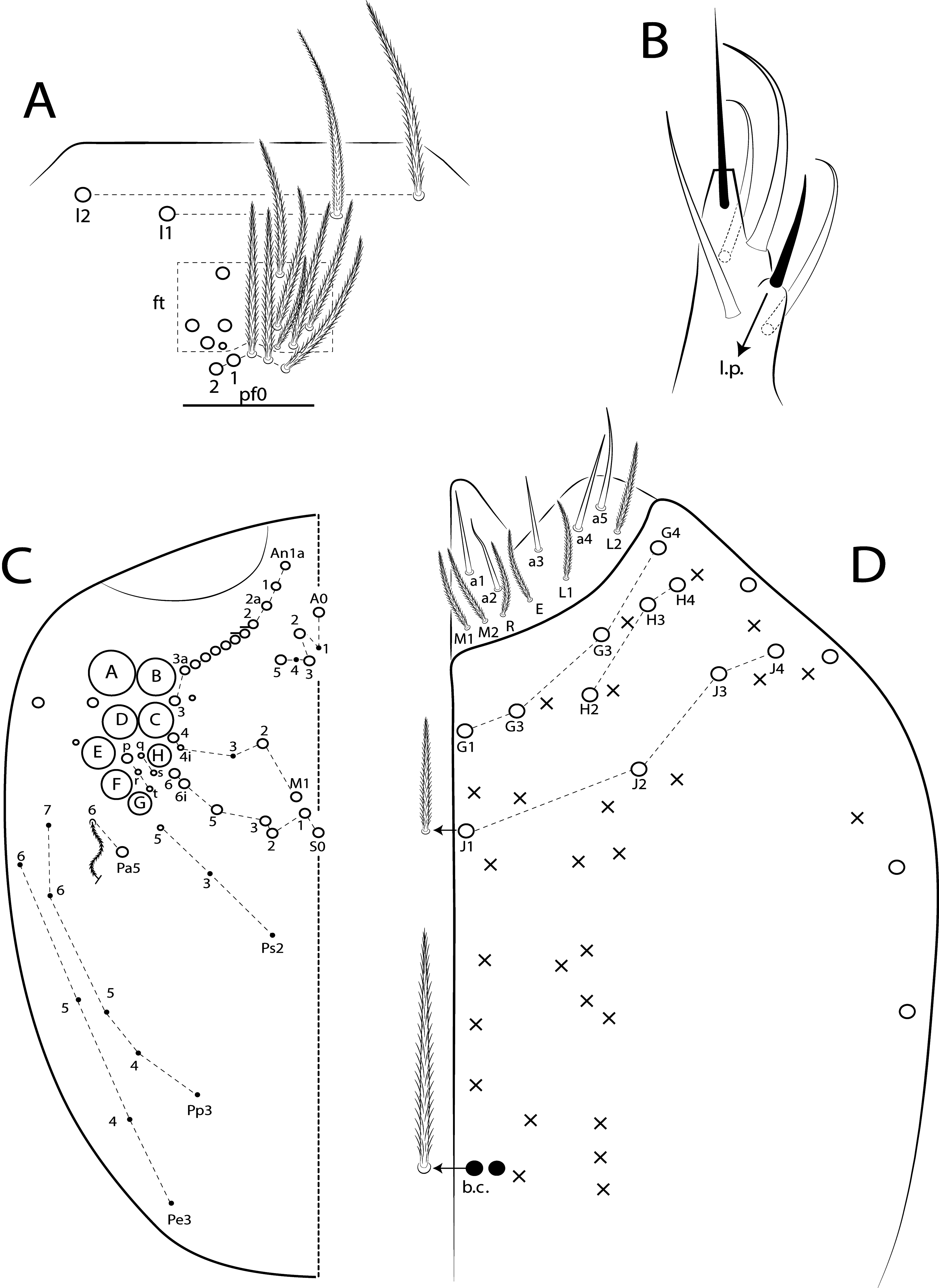
I: II: III: IV = 1: 1.38: 1.38: 2.70 in holotype (Ant III–IV missing in paratype). Ant IV annulated on distal four fifth, with apical bulb apically bilobed. Ant III not annulated, apical organ with 2 rods, 3 spiny guard sens, s-blunt sens of different sizes and ciliated chaetae.Ant I dorsally with 4 median mac surrounded by ciliated guard mic and 3 smooth mic at the base, without spines. Eyes A and B larger, G and H smaller, others subequal, and with 5 ‘IO’ chaetae; head dorsal macrochaetotaxy with 11–14 An, 4 A, 3 M and 7 S mac ( Fig. 78C View FIGURE 78 ). Clypeal formula with 6 (l1–2), 8 (ft), 5 (pf0–2) ciliated chaetae, l1–2 larger than the others, l1 apically acuminate, 2 ft smaller, others subequal ( Fig. 78A View FIGURE 78 ). Prelabral chaetae smooth and not bifurcated. Labral p0 chaeta with median filament, p1 larger than the others. Labral papillae with two inner rounded projections, outer papillae absent. Maxillary palp with smooth t.a. and b.c. weakly ciliated, thicker and 1.44 longer than the t.a. Labial papilla E with l.p. conical and reduced ( Fig. 78B View FIGURE 78 ). Basolateral and basomedian labial fields with chaetae M1–2, R (smaller than the others), L1–2 ciliated, e smooth. Ventral head with about 15 ciliated chaetae, postlabial formula with 4 (G1–4), 3 (H2–4), 4 (J1–4) chaetae, and 1 larger b.c. surrounding the cephalic groove ( Fig. 78D View FIGURE 78 ).
Thorax chaetotaxy ( Fig. 79 View FIGURE 79 A–B). Th II projected anteriorly, a, m and p series with 7, 8 and 10–11 mac, respectively, and about 5 secundary psp. Th III a, m and p series with 6, 1 and 8 mac, respectively, and about 6 secundary psp. Ratio Th II: III = 1.94–1.82: 1 (n=2), holotype 1.94: 1.
Abdomen chaetotaxy ( Figs 79 View FIGURE 79 C–D). Abd I a, m and p series with 1, 4 and 0 mac, respectively, and about 5 secundary psp. Abd II a, m and p series with 1, 4 and 0 mac, respectively, and about 5 secundary psp. Abd III a, m and p series with 0, 3 and 1 mac, respectively, and about 8 secondary psp. Abd IV with 10–12 central mac on A–T series (C1 as mac or mic) and 14–16 lateral mac on E–Fe series; about 9 secondary psp, at least 5 posterior sens (ps type I, others type II) and 3 posterior mes. Abd V a, m and p series with 1, 4 and 6 mac, respectively. Chaetotaxy of Abd III–V similar to L. andevo sp. nov. ( Figs 64 View FIGURE 64 A–C). Ratio Abd III: IV = 1: 5.15–5.44 (n= 2), holotype 1: 5.44.
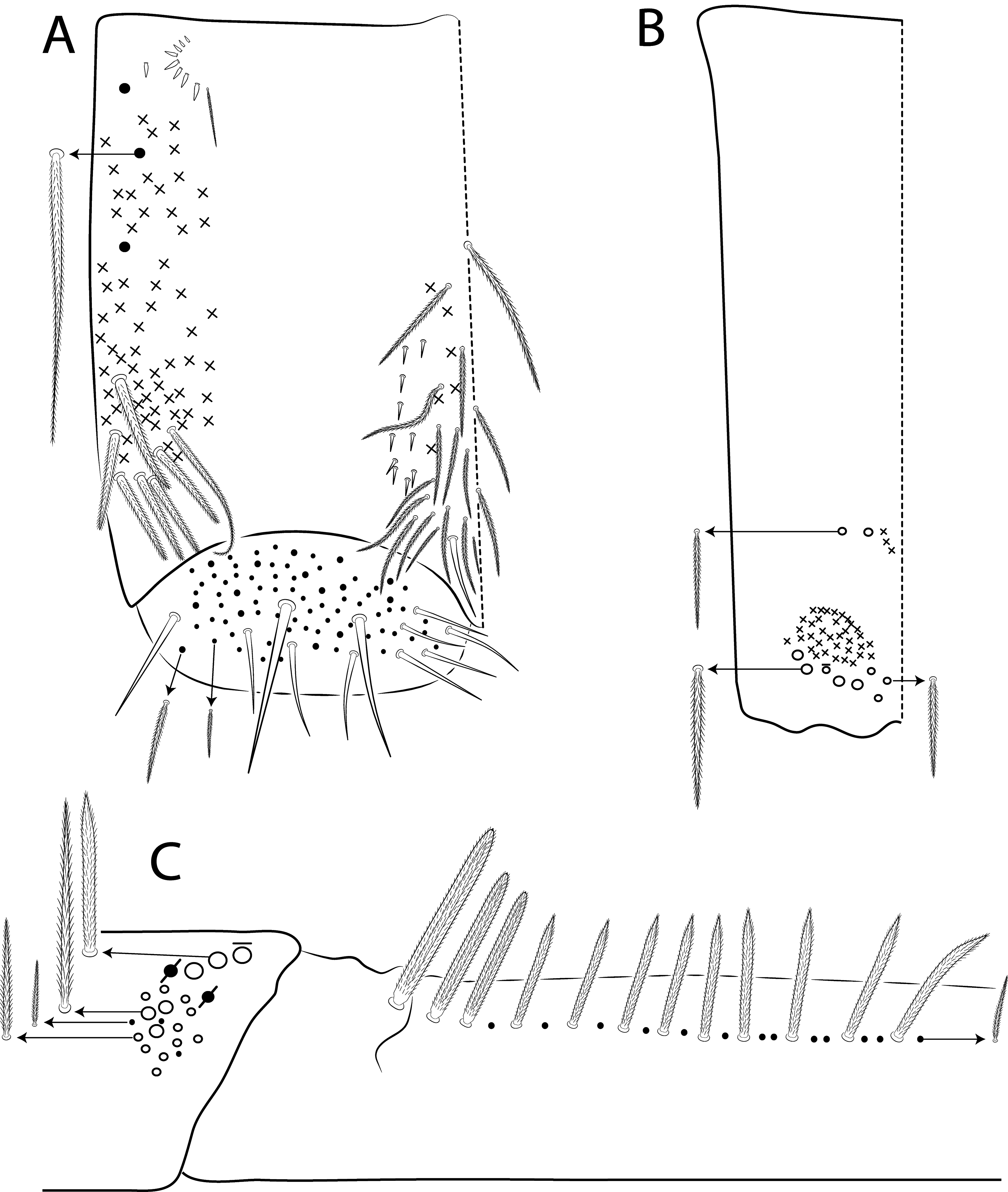
Legs. Subcoxa I with 6 chaetae on a row, 3–5 anterior chaetae and 2 psp; subcoxa II with an anterior row of 16 chaetae, posterior row of 8 chaetae and 4 psp; subcoxa III with one row of 13 chaetae, 1 anterior chaeta and 2 posterior psp ( Fig. 80 View FIGURE 80 A–C). Trochanteral organ with about 51 spine-like chaetae ( Fig. 80D View FIGURE 80 ). Tibiotarsus III normal or subdivided on slightly more than its distal half, outer side with 4 large chaetae apically acuminate, inner side with 7 mac weakly ciliated. Unguis with 3 inner teeth, b.t. smaller to m.t., a.t. absent. Unguiculus with all lamellae smooth and acuminate, except pe with a small proximal tooth; ratio unguis: unguiculus = 1: 0.69. Tibiotarsal smooth chaeta 0.82 smaller than unguiculus and tenent hair 0.73 smaller than unguis outer edge.
Collophore. Anterior side with 20 chaetae, 8 proximal spine-like chaetae, 4 ciliated chaetae, 6 ciliated chaetae apically acuminate and 2 mac distally; posterior side with 10 chaetae (1 unpaired), 6 spines, 3 thin ciliated chaetae widely distributed and 1 smooth chaeta distally; lateral flap with about 58 chaetae, 12 smooth and 46 ciliated ( Fig. 80E View FIGURE 80 ).
Furcula. Manubrium ventral formula with 0, 0, 0, 0/4 (subapical), 12 (apical) ciliated chaetae plus approximately 26 elongated apical scales per side ( Fig. 80F View FIGURE 80 ); manubrial plate with 2 blunt mac ciliated abruptly acuminate at the apex, 18 ciliated chaetae of different sizes and 3 psp. Dens dorsally with one proximal row of 4–7 blunt mac weakly ciliated and apically bullet-like, holotype with 4+6 ( Fig. 80G View FIGURE 80 ).
Etymology. The name “iegoi” (in apposition) refers to nickname of our friend and collembologist Msc. Diego Dias da Silva (INPA), who helped us providing specimens from Madagascar.
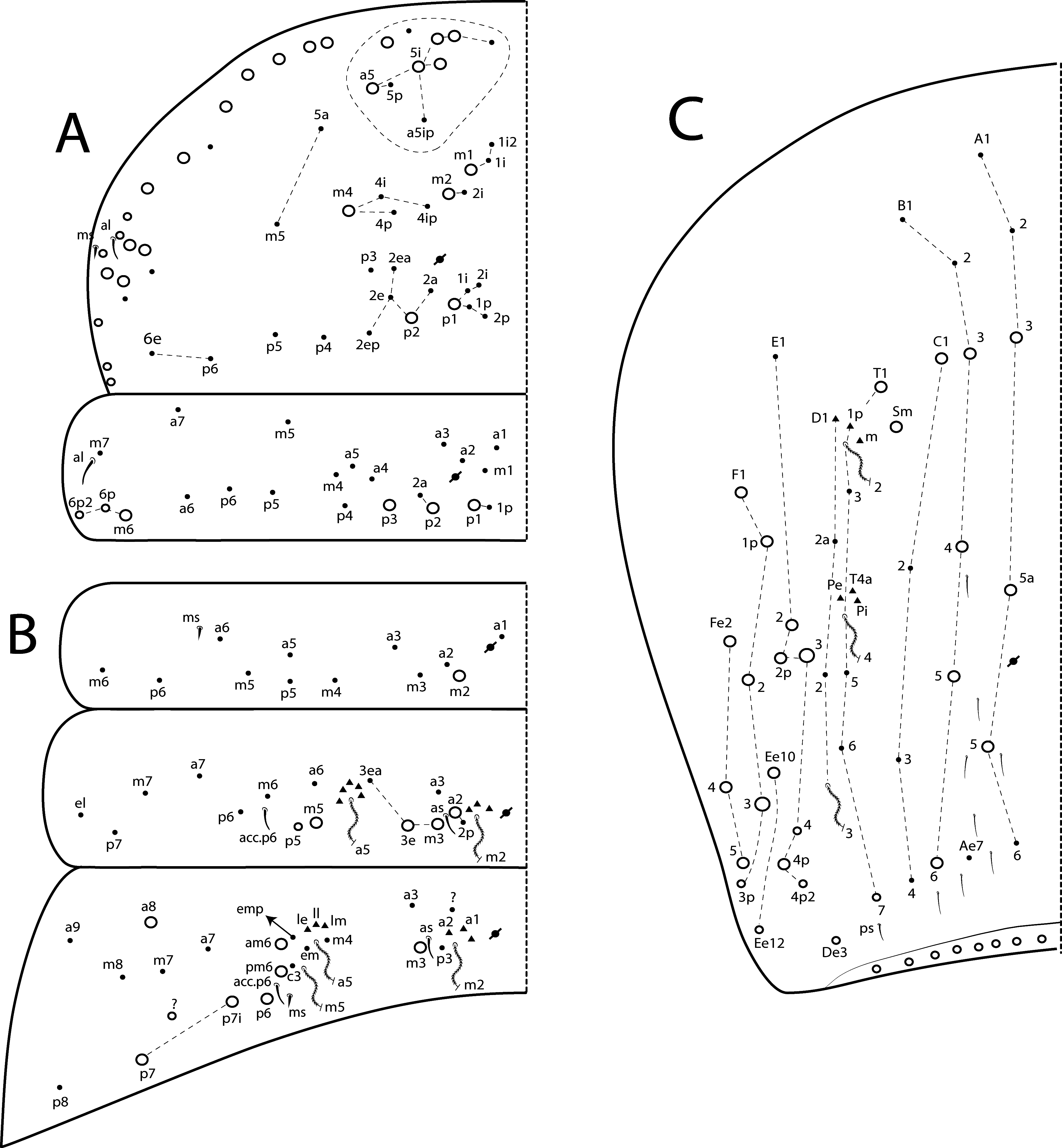
Remarks. The four new species herein described from Madagascar are somehow similar to each other in habitus, general morphology and dorsal chaetotaxy patterns ( Figs 61–80 View FIGURE 61 View FIGURE 62 View FIGURE 63 View FIGURE 64 View FIGURE 65 View FIGURE 66 View FIGURE 67 View FIGURE 68 View FIGURE 69 View FIGURE 70 View FIGURE 71 View FIGURE 72 View FIGURE 73 View FIGURE 74 View FIGURE 75 View FIGURE 76 View FIGURE 77 View FIGURE 78 View FIGURE 79 View FIGURE 80 ). However, they differ from each other by color pattern as: appendages heavily pigmented in L. andevo sp. nov., partially pigmented in L. briani sp. nov. and L. betamponensis sp. nov., and reduced in L. iegoi sp. nov. ( Fig. 77 View FIGURE 77 ). The head morphology of all new species from Madagascar lack Pp5 mac (present in L. voeltzkowi , Fig. 57D View FIGURE 57 ), labial papilla E with l.p. reduced in L. andevo , L. betamponensis and L. iegoi (reaching the base of a.a. in L. voeltzkowi and L. briani ), basomedian labial field with chaeta E ciliated in L. voeltzkowi and L. briani (smooth in L. andevo , L. betamponensis and L. iegoi ), and ventral cephalic groove with 1 b.c. in L. voeltzkowi , L. andevo and L. iegoi (2 in L. betamponensis and L. briani ).
The dorsal chaetotaxy of the Madagascar species differs in: L. voeltzkowi Th II with more than 12 anterior mac in a5 group (7 or less on the four new species) and posteriorly with p3 as mic and p2ep as mac (the opposite in the four new species). In Th III there are 2 lateral mac in L. voeltzkowi and L. briani , while in the other new species there are 4 mac. Other differences in Th III are: a5 mac only present in L. iegoi sp. nov., a1 mac only absent in L. briani , p1i mac absent in L. andevo and L. briani , and a4 mac only absent in L. voeltzkowi ( Figs 58 View FIGURE 58 A–B, 63A–B, 69A–B, 74A–B, 79A–B).
Concerning the body appendages, the Madagascar species differ in number of spine-like chaetae on trochanteral organ and unguis a.t. present in L. briani and L. voeltzkowi ( Fig. 60E View FIGURE 60 ). The collophore anteriorly differs by 6 distal mac present in L. andevo (3 in L. voeltzkowi and 2 in the other three new species) and about 16 spine-like chaetae in L. voeltzkowi (less in the four new species). On the posterior face of the collophore, the species differ by 10 spines present in L. andevo (8 in L. briani and L. betamponensis , 6 in L. iegoi , 2 in L. voeltzkowi ) and smooth chaeta present in the four new species (absent in L. voeltzkowi ). Finally, in dens they differ by blunt mac: apically nipple-like in L. voeltzkowi , bullet-like and/or abruptly acuminate at the apex in L. andevo , L. betamponensis and L. iegoi (only bullet-like in this species), while such chaetae are completely absent in L. briani . More comparisons are listed in Table 5 View TABLE 5 .
Key to Lepidocyrtinus species from Madagascar
1 Head without Pp5 mac ( Fig. 62F View FIGURE 62 ); Th II with 7 or less anterior mac (a5 group), posteriorly with p3 mac, and p2ep absent ( Fig. 63A View FIGURE 63 )............................................................................................... 2
- Head with Pp5 mac ( Fig. 57D View FIGURE 57 ); Th II with more than 12 anterior mac (a5 group), posteriorly with p3 as mic and p2ep as mac ( Fig. 58A View FIGURE 58 )..................................................................... L. voeltzkowi (B̂rner, 1907)
2 Basomedian labial field with chaeta e smooth ( Fig. 73E View FIGURE 73 ); Th III with 4 lateral mac ( Fig. 63B View FIGURE 63 ); Abd II with m3e mac ( Fig. 63D View FIGURE 63 ); unguis a.t. absent ( Fig. 65E View FIGURE 65 )............................................................................. 3
- Basomedian labial field with chaeta E ciliated ( Fig. 68D View FIGURE 68 ); Th III with 2 lateral mac ( Fig. 69B View FIGURE 69 ); Abd II m3e mac absent ( Fig. 69D View FIGURE 69 ); unguis a.t. present (as Fig. 60E View FIGURE 60 )........................................................ L. briani sp. nov.
3 Body with pigments ( Figs 61 View FIGURE 61 , 72 View FIGURE 72 ); Th III a5 always as mic ( Fig. 74B View FIGURE 74 ), Abd I with 4 or less central mac ( Fig. 63C View FIGURE 63 , 74C View FIGURE 74 ).... 4
- Body pale ( Fig. 77 View FIGURE 77 ); Th III with a5 mac ( Fig. 79B View FIGURE 79 ), Abd I with 5 central mac Fig. 79C View FIGURE 79 ).................. L. iegoi sp. nov.
4 Femur I–III completely pigmented ( Fig. 61 View FIGURE 61 ); Abd I often with 3 central mac, rarely with 4 mac (a2 often mic) ( Fig. 63C View FIGURE 63 ); collophore anteriorly with 6 distal mac ( Fig. 66A View FIGURE 66 )................................................ L. andevo sp. nov.
- Femur I–III distally pigmented ( Fig. 72 View FIGURE 72 ); Abd I often with 2 central mac, rarely with 1 mac (m3 often mac) ( Fig. 74C View FIGURE 74 ); collophore anteriorly with 2 distal mac ( Fig. 76E View FIGURE 76 ).......................................... L. betamponensis sp. nov.
Descriptions of intermediary stages of Lepidocyrtinus
The juvenile’s description of Lepidocyrtinus is based on L. africanus sp. nov. and L. fabioi sp. nov., both probably in 3 rd instar due to their similar dorsal chaetotaxy compared to the same instar of Seira (Soto-Adames 2008; Zhang et al. 2019). In L. africanus it was studied the development of the colour pattern, antennal segments, clypeus, all head and dorsal trunk chaetotaxy, trochanteral organ, collophore and furcula. For L. fabioi only the dorsal chaetotaxy was studied, since other characteristics are similar to the adult. The morphology of both species is compared to 3 rd instar Seira dowlingi ( Wray, 1953) and adult, and to L. barnardi adult (Soto-Adames 2008; Zhang & Deharveng 2015a, 2015b; Zhang et al. 2019).
Colour pattern. Specimen with weak dark pigments on Ant II to IV, and Abd III with 1 lateral spot similar to adult (in coxa-shape) ( Fig. 23 View FIGURE 23 ). In Lepidocyrtinus this gain of pigments in postembryonic development must be common, as observed in other Entomobryidae ( Pan et al. 2011, 2015, 2019; Cipola et al. 2017, 2018a).
Scales. Present on all head, dorsal and lateral thorax and abdomen; few scales on coxa and femur II–III, and manubrium and dens ventrally; antennae and collophore unscaled. Such displacement was observed in 3 rd instar S. dowlingi (Soto-Adames 2008) .
Antennal segments. Antennae smaller than trunk length ( Fig. 23B View FIGURE 23 ); antennal segments ratio as I: II: III: IV = 1: 1.89: 2.56: 3.79. Ant I–IV chaetotaxy reduced. Ant IV weakly annulated with apical bulb apically unilobed. Ant III not annulated. Ant II with 2 bothriotricha and 1 median mac, sensilar chaetotaxy unclear. Ant I dorsally with 13 chaetae (2 lateral mac), 1 distal sens (not seen in adults), and typically with 3 smooth mic at the base, bothriotricha absent ( Fig. 30A View FIGURE 30 ). This same bothriotricha pattern on Ant I–II was observed in 3 rd instar S. dowlingi (Soto-Adames 2008) , and so there are no clear generic differences regarding this topic.
Clypeus ( Fig. 30B View FIGURE 30 ). Clypeal formula as in the adult, with 4 (l1–2), 2 (ft) and 3 (pf0–1) ciliated chaetae, but l1–2 apically acuminate. It is most likely all these chaetae are primary and homologous to other Entomobryoidea ( Zhang et al. 2016). Consequently, extra chaetae present in adults of some species of Lepidocyrtinus ( Figs 14C View FIGURE 14 , 19B View FIGURE 19 , 62C View FIGURE 62 , 68A View FIGURE 68 , 73A View FIGURE 73 , 78A View FIGURE 78 ), as well as in other Entomobryoidea in general, are secondary chaetae of difficult homology to determine. The emergency of extra secondary clypeal chaetae during the development was also reported to Symphypleona ( Betsch & Waller 1994).
Interocellar chaetae ‘IO’ ( Fig. 30C View FIGURE 30 ). Interocellar field with 3 chaetae (p, r and t, p as mac), all primary according to Soto-Adames (2008). Therefore, the secondary chaetae (at least q and s) should appear in Lepidocyrtinus probably from the 4 th instar, as observed in Seira .
Head dorsal chaetotaxy ( Fig. 30C View FIGURE 30 ). Differs from adult by 5 An, 3 M (m4i as mic), 3 Pp (Pp2 and Pp3), and 1 Pe mac (Pe3). In adult or any instar after the 3 rd, the An series has additions of mac, m4i turns into mac, and there is also the reduction of 3 mac (Pa2, Pa3 and Pe3), at least in L. africanus sp. nov. ( Fig. 24F View FIGURE 24 ). In S. dowlingi , only occurs transformations of mic to mac during the development (Soto-Adames 2008), and therefore, such reductions of postoccipital mac in Lepidocyrtinus can be an exclusive feature in the Seirinae . In Entomobryinae genera as Sinhomidia Zhang, 2009 (in Zhang et al. 2009) it was also observed the reduction in some postoccipital mac of 1 st to adult, while in Homidia B̂rner, 1906 the secondary mac emerges during other instars’ development ( Pan et al. 2011, 2019). In this sense, a study with the complete development (1 st to adult) needs to be done contemplating more Seira and Lepidocyrtinus species to make sure if there is a generic pattern.
Basomedian and basolateral labial fields ( Fig. 30D View FIGURE 30 ). Similar to the adult ( Fig. 24G View FIGURE 24 ), except by M2 and R chaetae absent. Such condition of the 3 rd instar is shared with Seira (Soto-Adames 2008) . Chaetae a1–5, M1 (as M), E and L1–2 are primary of Entomobryoidea, and it is common the emergence of extra chaetae (at least M2, R) during the 3 rd or 4 th instar (Soto-Adames 2008; Pan et al. 2011, 2019; Zhang & Pan 2020).
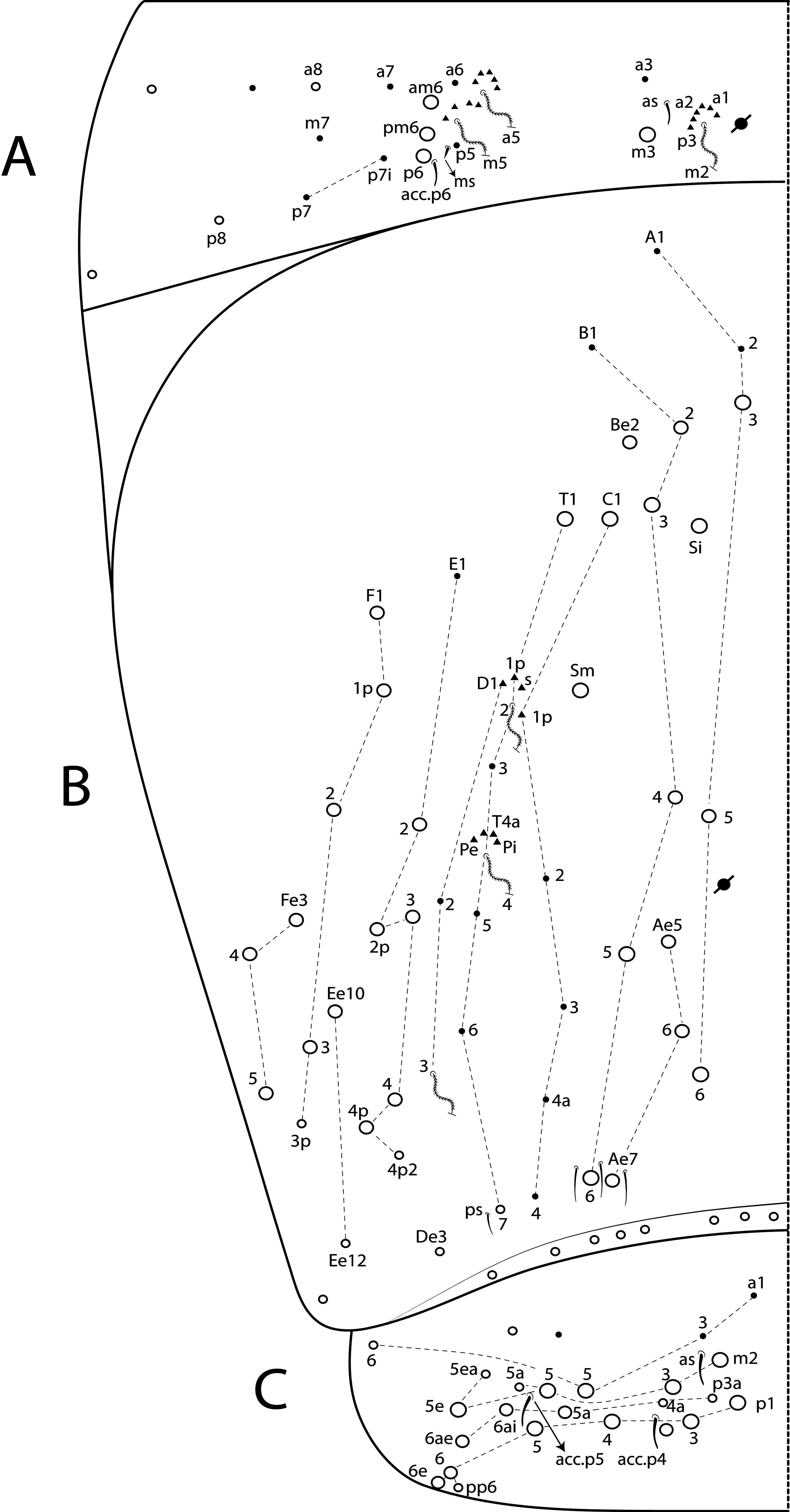
Postlabial field ( Fig. 30D View FIGURE 30 ). Formula of the main chaetae as in the adult ( Fig. 24G View FIGURE 24 ), with 4 (G1–4), 3 (H2–4), 3 (J1–3) chaetae, except by a sole larger chaeta posteriorly, between J1 and J2. Therefore, the secondary chaetae in this region appear at least after the 3 rd instar. Next to J3 and J4 chaetae, postero-laterally, there are two unnamed chaetae both in the juvenile and the adult of L. africanus sp. nov., as well as in other species like L. botswanensis sp. nov., L. voeltzkowi , L. betamponensis sp. nov. and L. iegoi sp. nov., and even Seira and Tyrannoseira taxa ( Cipola et al. 2018b, 2018c; 2019). Of such chaetae, the GHJ series are, quite probably, primary and present in all Entomobryoidea, since they were also observed in the 1 st instar of Homidia and Sinhomidia ( Pan et al. 2011, 2019). However, so far little is known about the development of the other postlabial setae, since the development of this region has never been properly studied.
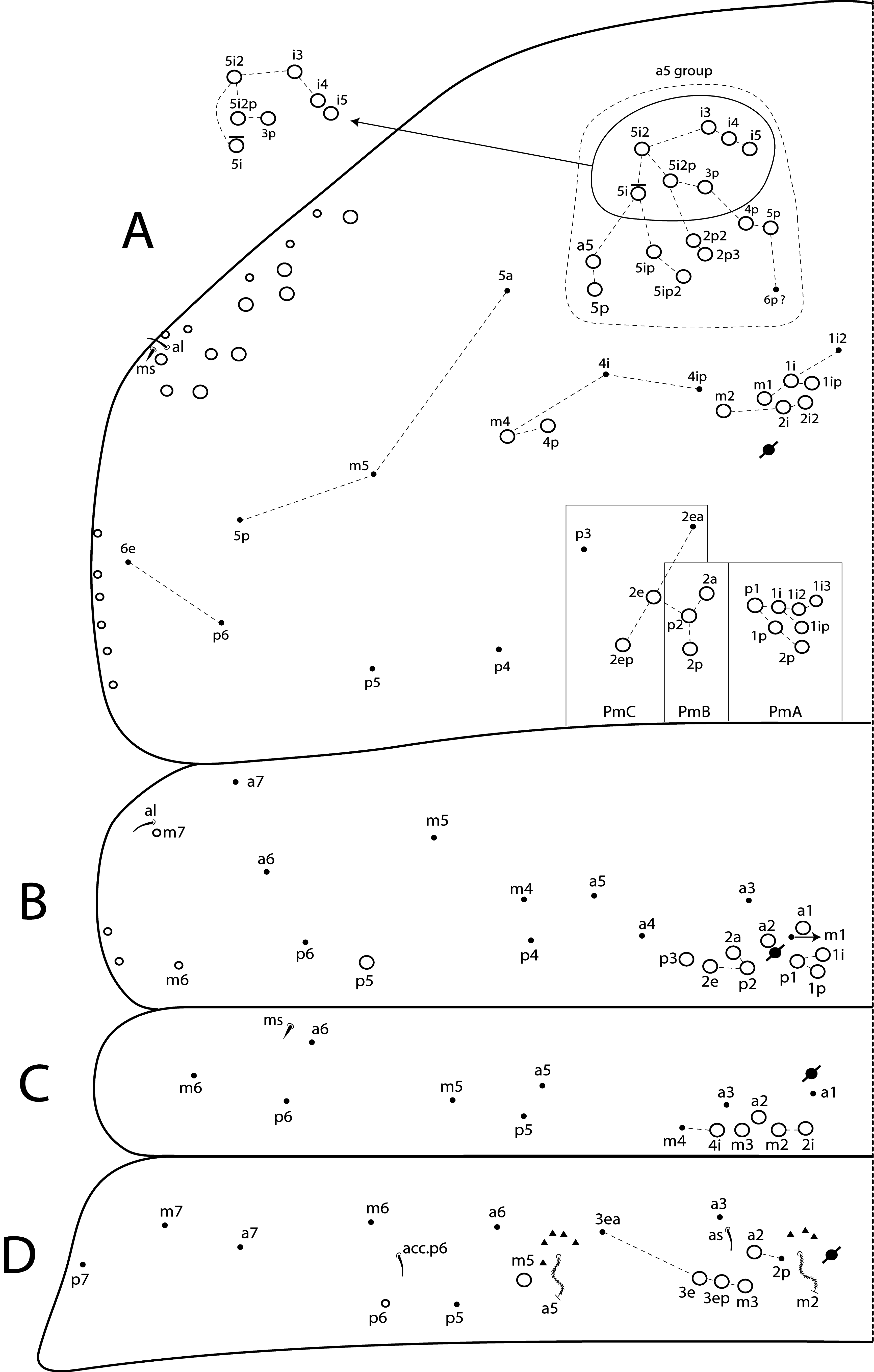
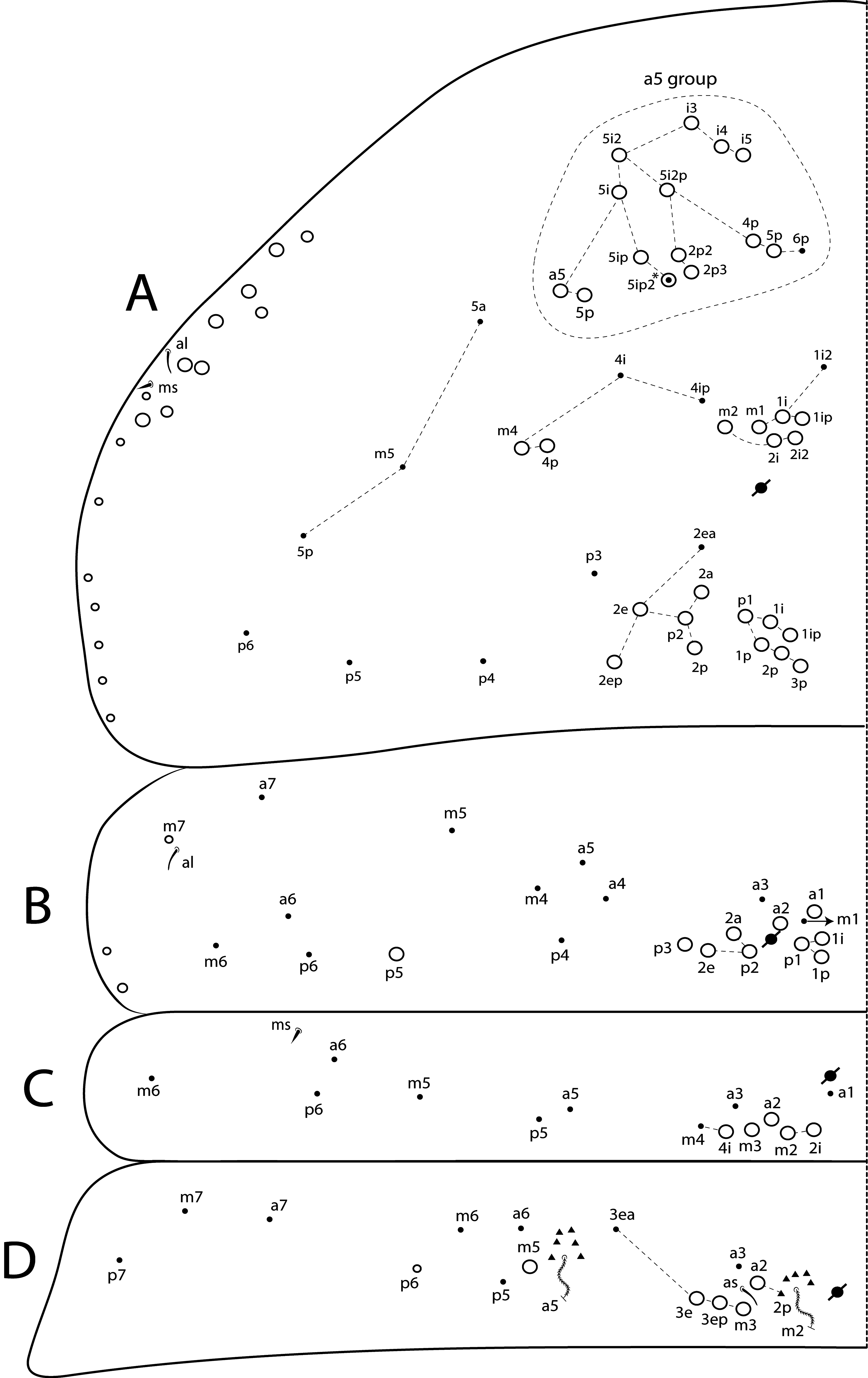
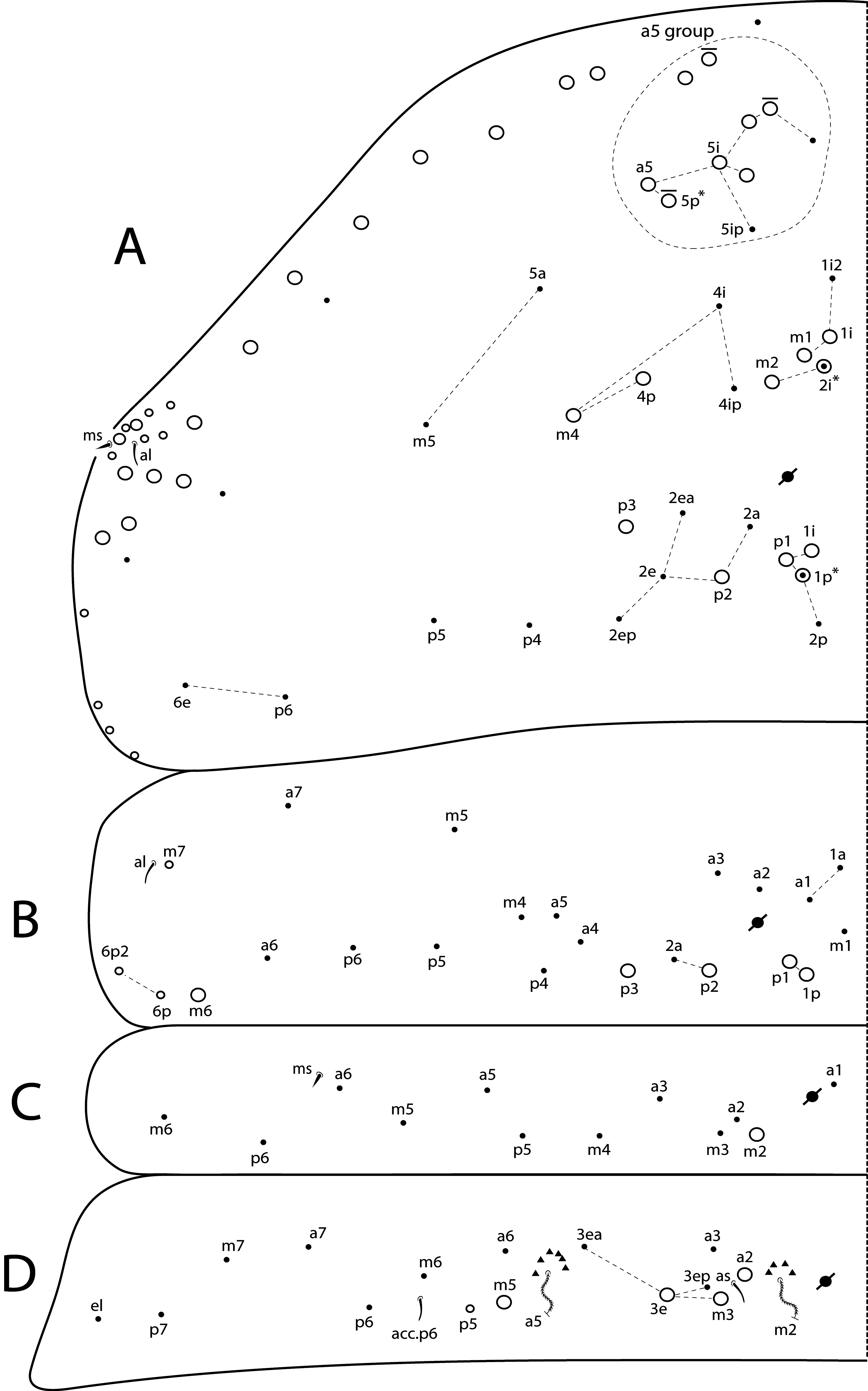
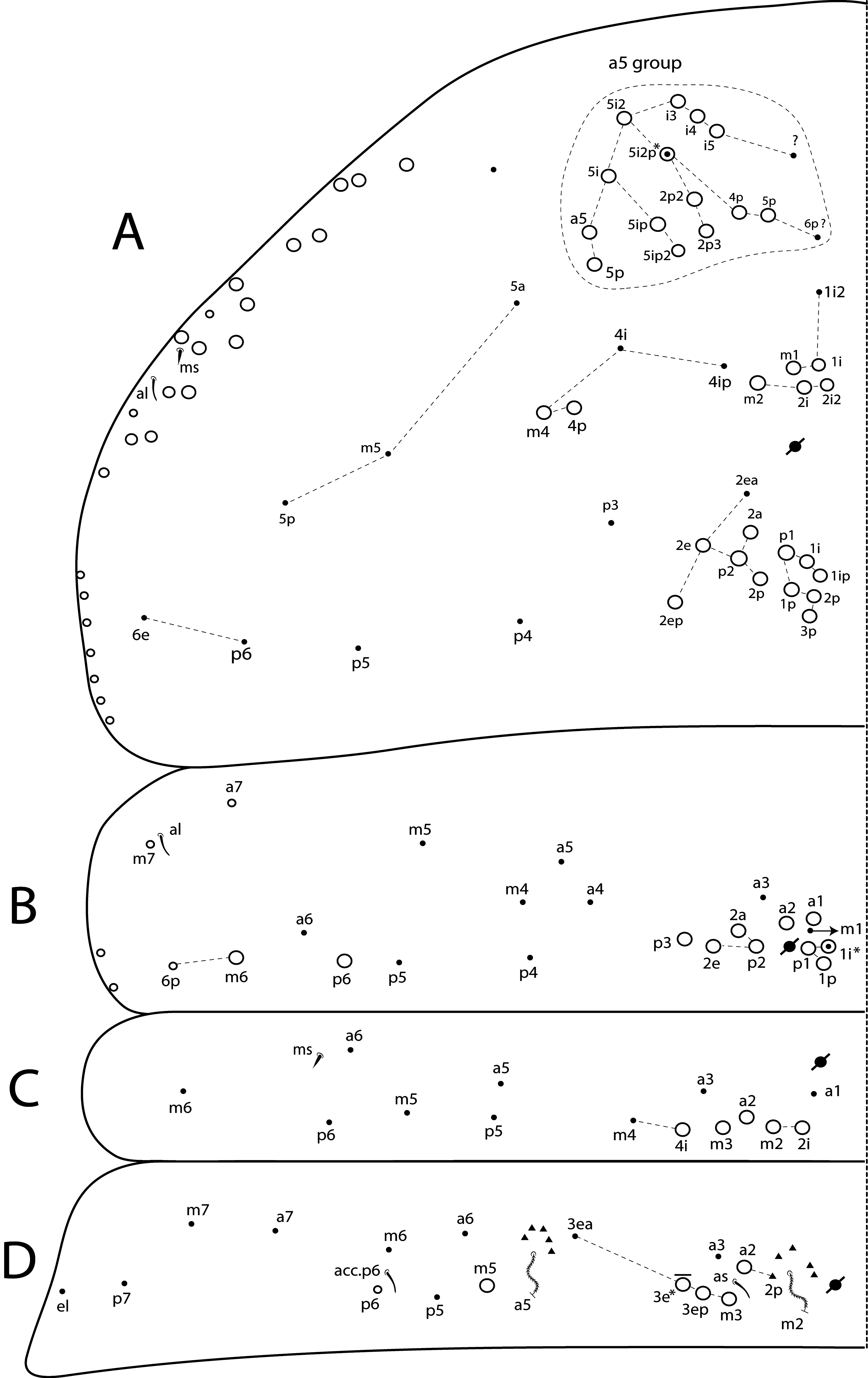
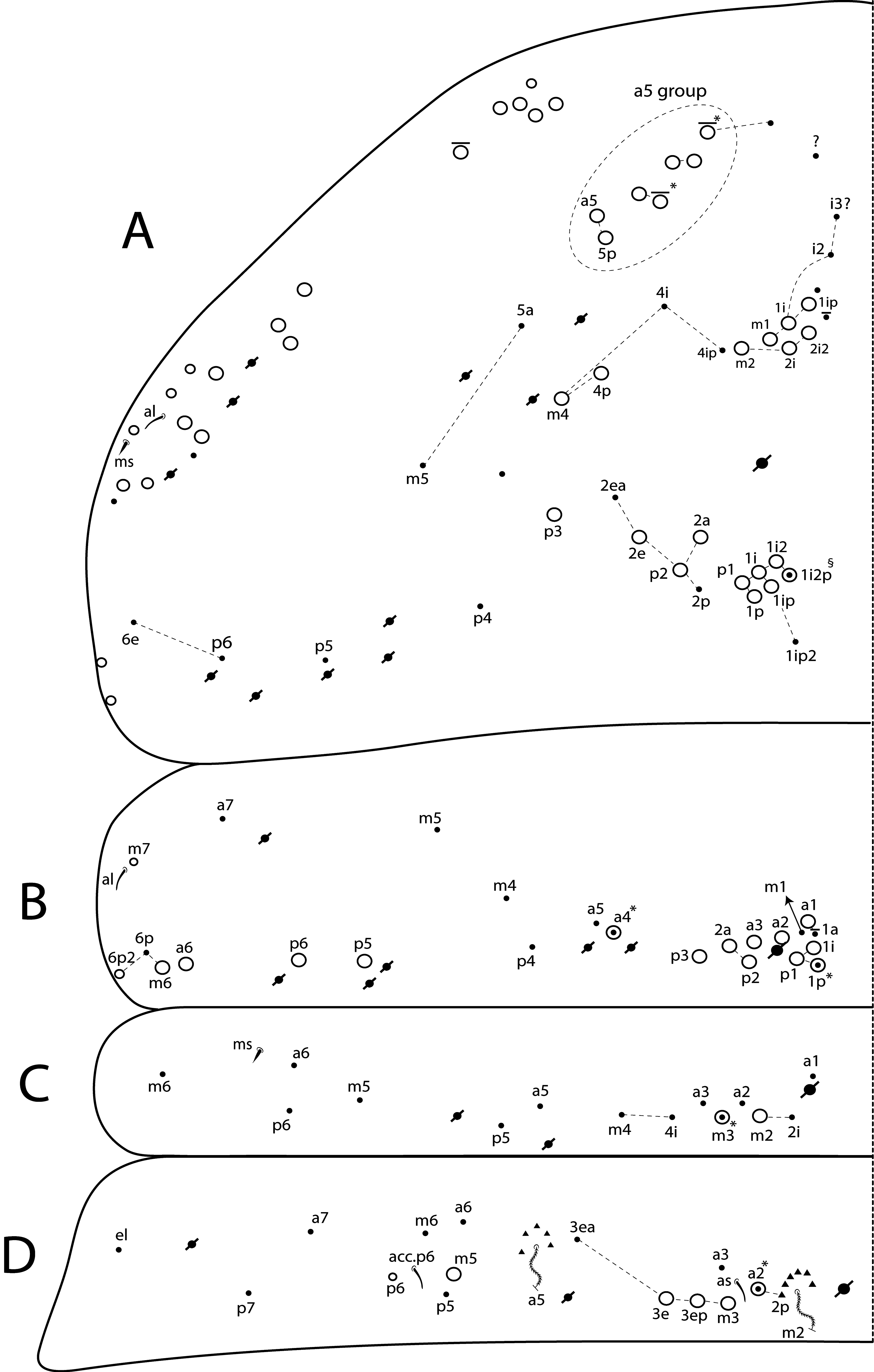
Th II dorsal chaetotaxy ( Figs 31A View FIGURE 31 , 44A View FIGURE 44 ). In L. africanus sp. nov., the juvenile’s Th II a series has 6 mac (a5p–a5i3, a5i2p), and 6 mic on a5 group (a5i4, a5i3p– 6p, a5ip), the latter probably gradually turning into mac until the adult, as well as the appearance of extra chaetae (a5ip2, a5i2p2–p3, a5i5) ( Fig. 25A View FIGURE 25 ). These chaetae are extra rows of a5i, which are shared among Lepidocyrtinus species ( Figs 35A View FIGURE 35 , 41A View FIGURE 41 , 47A View FIGURE 47 , 52A View FIGURE 52 , 58A View FIGURE 58 ). So, they are probably homologous in most species, except for L. barnardi and L. dayi ( Figs 15A View FIGURE 15 , 20A View FIGURE 20 ) which are atypical concerning such multiplets ( Figs 15A View FIGURE 15 , 20A View FIGURE 20 ). Regardless, in all African Lepidocyrtinus species there are commonly 8 mac (except for L. fabioi sp. nov. with 6 mac) in two transvesal rows posteriorly on a5 group (e.g. Fig. 35A View FIGURE 35 ), which are absent in Seira and Tyrannoseira (Soto-Adames 2008; Cipola et al. 2014a, 2014b, 2018b, 2018 c, 2019). These transvesal rows are reduced and some chaetae do not show clear homologies in L. fabioi ( Fig. 41A View FIGURE 41 ), so it is not clear if they represent the same seen in other species or emerged independently in the latter species. Juveniles of L. fabioi have almost the same pattern as the adults in a5 group, except for two chaetae as mic (a5p and another anteriorly of uncertain homology), that become mac in adults ( Figs 41A View FIGURE 41 , 44A View FIGURE 44 ).
The juvenile’s m and p series of L. africanus are quite similar to the adult, except by m1ip mac absent and only 4 mac (p1i–p2p) present on PmA group ( Figs 31A View FIGURE 31 , 25A View FIGURE 25 ). However, the juvenile of L. fabioi only hold primary chaetae as mac in m and p series, and p3 emerges first as a mic and becomes a mac in the adult together with five other secondary chaetae (m1i, m2i, m4p, p1i and p1p) ( Figs 41A View FIGURE 41 , 44A View FIGURE 44 ). In p series, p5 is always mic and sometimes p3 too, in Lepidocyrtinus adults (e.g. Figs 25A View FIGURE 25 , 35A View FIGURE 35 , 47A View FIGURE 47 ). Both chaetae are also mic in the 3 rd instar, at leats in L. africanus and L. fabioi ( Figs 31A View FIGURE 31 , 41A View FIGURE 41 ). In the 1 st instar of S. dowlingi and L. barnardi , such chaetae are mac (Soto-Adames 2008; Zhang et al. 2019), but p5 becomes a mic in adult L. barnardi ( Fig. 15A View FIGURE 15 ). Further juveniles of different taxa of Lepidocyrtinus need to be investigated to see if p3 is always a mic in the 1 st instar or if changes its state (from mac to mic) at any instar in different species. Thus, notably the chaetotaxy final pattern of occurs only in adults of both genera.
Th III dorsal chaetotaxy ( Figs 31A View FIGURE 31 , 44A View FIGURE 44 ). The juveniles of the 3 rd instar are similar to the adults in L. africanus sp. nov. ( Figs 24B View FIGURE 24 , 31A View FIGURE 31 ) and L. fabioi sp. nov. ( Figs 41B View FIGURE 41 , 44A View FIGURE 44 ), except by p1i mac absent and m7 as mac (as mes in adult) in L. africanus , and a1a absent and p1p as mic in L. fabioi . Such data support that in the 3 rd instar of Lepidocyrtinus the diagnostic chaetotaxy pattern of each species is not yet fully defined, while in Seira such pattern is well established at the 2 nd instar, at least in S. dowlingi (Soto-Adames 2008) .
Abd I–III dorsal chaetotaxy ( Fig. 44B View FIGURE 44 ). The 3 rd instar Abd I–II chaetotaxy are the same as the adult in L. africanus sp. nov. and L. fabioi sp. nov. ( Figs 41 View FIGURE 41 C–D, 44B), and such similarity is also shared in the development of S. dowlingi (Soto-Adames 2008) .
The Abd III of the juvenile of L. africanus is the same as in the adult, but in L. fabioi some modifications occur, as two mac are reduced in mes (a8 and p7), a9 and p8 mic and one mes of uncertain homology disappear, and em and emp change into accessory chaetae ( Figs 42A View FIGURE 42 , 44B View FIGURE 44 ). In S. dowlingi , the am6 mac change only in the 5 th instar (Soto-Adames 2008), while in Lepidocyrtinus such change occurs in the 3 rd instar, at least in L. africanus and L. fabioi .
Abd IV dorsal chaetotaxy ( Figs 31B View FIGURE 31 , 44C View FIGURE 44 ). The 3 rd instar of L. africanus sp. nov. has 6 central mic (Ae6, B2, B4, B6, C1, T1) which become mac in the adult (or any stage before) ( Figs 26B View FIGURE 26 , 31B View FIGURE 31 ). In addition, in lateral series 3 mes are modified into mac during the development (E4, Ee10, Fe3), while Fe3 changes from mac to mes. Also, two accessory chaetae arise surrouding T4 bothriotrichum (Pe and other unnamed) and at least 2 sens next B6 mac (as sens not observed). On the other hand, in L. fabioi sp. nov., the macrochaetotaxy pattern of the 3 rd instar is most similar to the adult, at least in the central region ( Figs 42B View FIGURE 42 , 44C View FIGURE 44 ). On lateral series they differ only by the alteration of E4 mes into mac, and the emergence of Fe3 mac and s accessory chaeta surrouding T2 bothriotrichum, during the development. On Abd IV posterior region, more two mes occur only on the adult L. africanus , but in L. fabioi the number of 8 posterior mes seen in the adult is already established at the 3 rd instar. A similar development of the posterior chaetae was recorded to S. dowlingi , but the adult’s number is already reached during the 2 nd instar (SotoAdames 2008).
In the 3 rd instar as well as in the adult of all Lepidocyrtinus species, there may be one or two secondary mac on central series (Si and Sm), which most likely appear during the 2 nd instar, as in most Entomobryoidea, including Seira (Soto-Adames 2008; Zhang et al. 2019). Consequently, these mac are not useful to separate Lepidocyrtinus from other Seirinae , but to support interspecific diagnoses, as other secondary chaetae (e.g. A3a/5, Ae4–7, Be2) which can be present or absent in different species ( Figs 36B View FIGURE 36 , 48B View FIGURE 48 , 53B View FIGURE 53 , 59B View FIGURE 59 , 64B View FIGURE 64 ). Compared to the 3 rd instar of S. dowlingi , at least 8 chaetae (A5, B3, T7, E2, E4, E4p2, Ee10, F1) change into mac in the 4 th instar, plus 4 more mac on lateral series (F2p, F3p, Fe2, Fe3) arise, and in the 5 th instar 4 more accessory chaetae (s, m, Pi, Pe,) surrouding T2 and T4 bothriotricha emerge (Soto-Adames 2008). In this sense, Lepidocyrtinus and Seira development support that the final chaetotaxy pattern of Abd IV is only reached at the final instars, or even in the adult.
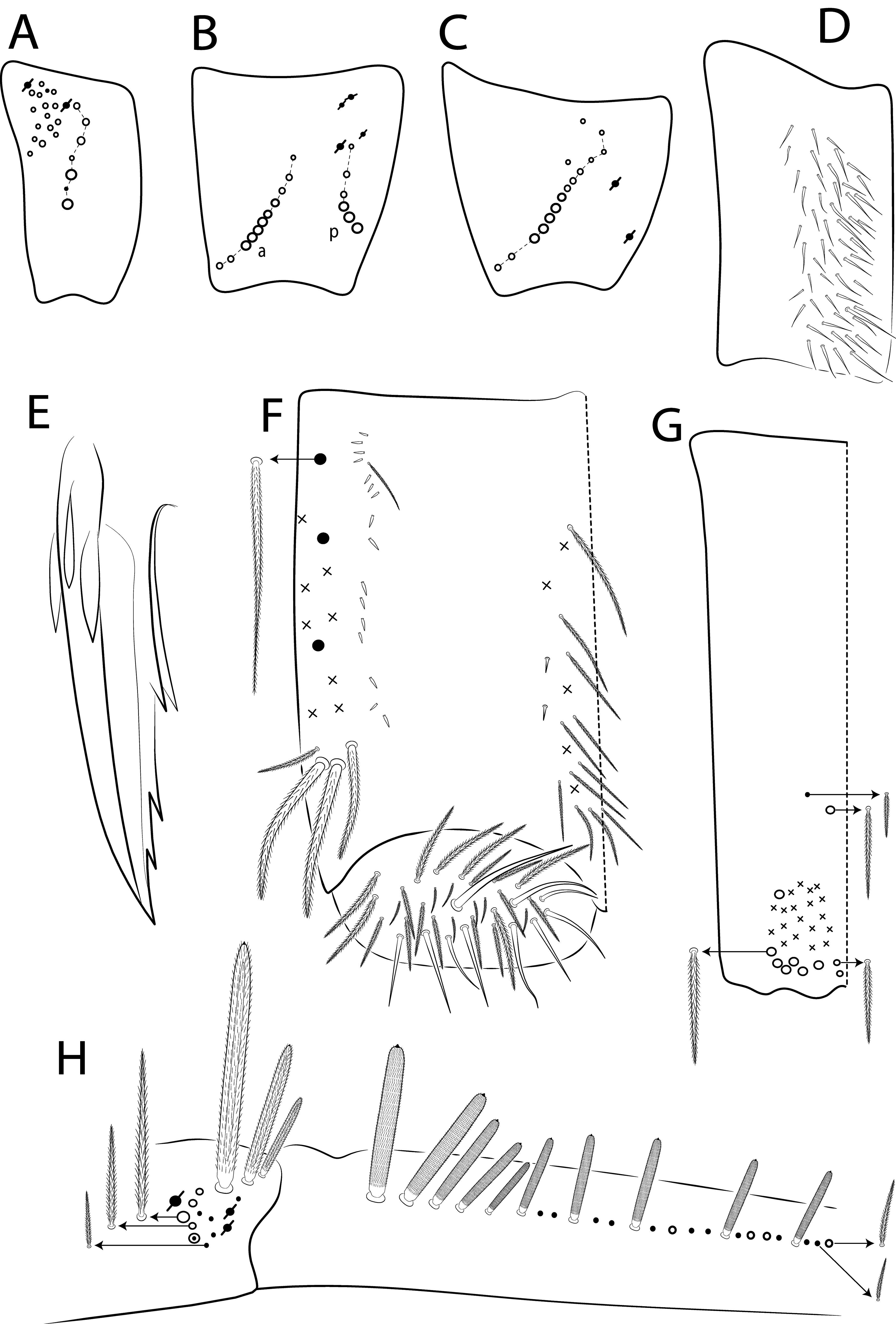
Legs ( Fig. 31E View FIGURE 31 ). Subcoxa and tibiotarsus chaetotaxy reduced in the juveniles. Tibiotarsal outer chaetae not clearly elongated and with few chaetae, similar to the 1 st instar of L. barnardi ( Zhang & Deharveng 2015a) . Thus, the elongation of these chaetae must occur at the final instars. The trochanteral organ has 4 spine-like chaetae ( Fig. 31C View FIGURE 31 ). Legs subsegments chaetotaxy show a gradual increase in complexity (number and shape of chaetae) until the adult in Lepidocyrtinus , and this probably occurs in all Entomobryoidea as well ( Pan et al. 2011, 2019). Unguis and unguiculus morphology of juveniles are as in the adults; ratio unguis: unguiculus = 1: 0.65. The tibiotarsal smooth chaeta is 1.49 larger than unguiculus and the tenent hair is 1.28 larger than unguis outer edge.
Collophore ( Fig. 31D View FIGURE 31 ). Anterior side with 6 chaetae, 2 spine-like chaetae and 1 proximal ciliated chaeta, 1 median ciliated chaeta apically acuminate and 1 mac and 1 mes distally; posterior side with 1 ciliated chaeta distally; lateral flap with 5 chaetae, 3 smooth and 2 ciliated. From these chaetae, at least 1+1 distal on posterior side and 2 smooth on lateral flap are probably primary, since they were also observed in the 1 st instar of Homidia and Sinhomidia ( Pan et al. 2011, 2019).
Furcula ( Figs 31 View FIGURE 31 E–F). Manubrium ventral formula as in the adult, except by 8 apical chaetae plus only 1 subapical scale per side ( Fig. 31E View FIGURE 31 ); manubrial plate with 5 ciliated chaetae (2 inner larger but not blunt) and 2 psp apparently ( Fig. 31F View FIGURE 31 ). Dens dorsally with 2 proximal blunt mac weakly ciliated and apically bullet-like ( Fig. 31F View FIGURE 31 ). It is still necessary to investigate further juveniles of Lepidocyrtinus in in the 1 st instar to clearly understand the origin and pattern of increase of furcal modified mac up to the adult. Our data suggests there is also an increase in the number of furcal chaetae during the development, both in the manubrium and dens, as well the change of states of at least part of them, similarly to what is seen in the legs subsegments.

























| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |