
Lepidophthalmus statoni, Felder, Darryl L., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3985.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:58E575F4-D7B7-4BD4-A2B2-7CDC756387C5 |
|
DOI |
https://doi.org/10.5281/zenodo.6114206 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF87C7-FF8A-FF97-FF06-E0122B36F802 |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidophthalmus statoni |
| status |
sp. nov. |
Lepidophthalmus statoni sp. nov.
( Figures 1 View FIGURE 1 a–k, 2 a–o, 3 a–f)
Lepidophthalmus spp.—Staton et al, 2000: figs. 1 (part), 2 (part), table 1 (part). Lepidophthalmus sp. “b”.—Staton et al, 2000: 161–168, fig. 3, tables 3–5. Lepidophthalmus sp. “b”.—Felder & Staton, 2000: 179.
Type material. Mexico (SW Gulf of Mexico). Holotype: male, cl 11.5 mm ( USNM 1275014), east bank of Rio San Pedro, upstream from mouth, 20 ppt salinity, seaward of Highway 180 bridge, village of Nuevo Campechito, Campeche, Mexico, 18°38.51'N; 92°27.88'W, sandy, organically rich, mud flat exposed at low tide among partially cleared red mangroves and stilted huts, 27 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman. Paratypes: 1 female, cl 12.1 mm, 1 female, cl 12.2 mm ( USNM 1275015), east bank of Rio San Pedro, upstream from mouth, 20 ppt salinity, seaward of Highway 180 bridge, village of Nuevo Campechito, Campeche, Mexico, 18°38.51'N; 92°27.88'W, sandy, organically rich, mud flat exposed at low tide among partially cleared red mangroves and stilted huts, 27 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman; 1 male, cl 10.0 mm, 1 female, cl 15.5 mm ( USNM 1275016), mud shorelines and flats in open area amid dense mangroves (9 ppt salinity), Rio Seco estuary, east of Puerto Ceiba, Tabasco, Mexico, 18°25.5'N; 93°07.0'W, tidally exposed muddy sand with detritus, 6 March 1981, D. L. Felder, W. W. Forman, and S. J. Snatic; 1 male, cl 13.3 mm, 1 male chela only, 3 females, fragmented vouchers for allozyme analyses, abdomens removed, cl 13.1–13.2 mm ( USNM 1275017), shoreline of ephemeral shallow back-beach pond (3 ppt salinity) on margin of tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44,'W, muddy sand, 29 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman; 1 male, cl 10.3 mm, 1 female, 10.7 mm ( UNAM / CNCR 30001), 1 male, cl 7.8 mm, 1 male cl 8.5 mm, 1 male, cl 9.5 mm, 1 female, cl 6.1 mm, 1 female, cl 9.5 mm ( ULLZ 16270), shallow bottom of tidal Rio Seco estuary (20 ppt salinity), along Highway 187, Puerto Ceiba, Tabasco, Mexico, 18°24.86'N; 93°10.38’W, sandy mud near mangroves, night, 27 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman; 1 male, cl 12.2 mm, 1 female, cl 8.6 mm ( UNAM / CNCR 30002), 1 male, cl 10.4 mm, 1 male, cl 10. 7 mm, 1 male, cl 11.1 mm, 1 ovigerous female, cl 12.5 mm, photo vouchers ( ULLZ 14795), shoreline of ephemeral shallow back-beach pond (2.5 – 3 ppt salinity) on margin of tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44,'W, muddy sand, 10 February 1999, D. L. Felder, S. C. Rabalais, C. D. Schubart, and C. F. Gurgel; 1 male, tissue voucher for DNA analysis, cl 13.1 mm ( ULLZ 5592), east side of Rio San Pedro, just inside river mouth (2.5 ppt salinity), seaward of Highway 180 bridge, village of Nuevo Campechito, Campeche, Mexico, 18°38.83'N; 92°27.91'W, muddy sand flat exposed at low tide, found with Leptalpheus forceps Williams, 1965 as symbiont, 7 July 2002, D. L. Felder and R. Robles; 1 male, cl 7.6 mm ( ULLZ 16271), shoreline of ephemeral shallow back-beach pond (3 ppt salinity) on margin of tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44,'W, muddy sand, 29 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman; 6 males, 1 voucher for DNA analysis, 5 females, 1 ovigerous ( ULLZ 7662), shoreline and shallow subtidal bottom <1m deep at inlet to Laguna La Mancha, Veracruz, Mexico, 19°35.36'N; 96°22.86'W, muddy sand, 2 July 2002, D. L. Felder, R. Robles, and I. T. Rodriguez; 1 male, cl 10.6 mm, tissue voucher for DNA analysis ( ULLZ 5613), shoreline, tidally exposed sand flat at inlet to Laguna La Mancha, Veracruz, Mexico, 19°35.36'N; 96°22.86'W, muddy sand, 3 July 2002, D. L. Felder, R. Robles, and I. T. Rodriguez; 1 female, cl 9.7 mm, 1 ovigerous female, cl 10.0 mm, 1 female, photo voucher, cl 10.5 mm ( ULLZ 14793), inlet to Laguna San Augustin seaward of bridge on Highway 180, Veracruz, Mexico, 19°55.27'N; 96°31.79'W, muddy sand flat at margins of shoreline mangroves and cordgrass, 9 February 1999, D. L. Felder, S. C. Rabalais, C. D. Schubart, C. F. Gurgel; 1 male, cl 11.3 mm, 1 female, cl 10.7 mm, photo vouchers ( ULLZ 15136), inlet to Laguna San Augustin seaward of bridge on Highway 180, Veracruz, Mexico, 19°55.27'N; 96°31.79'W, muddy sand flat at margins of shoreline mangroves and cordgrass, night, 7 July 2002, D. L. Felder and R. Robles.
Additional (non-type) material. Mexico, Campeche: 4 males, 8 females, east bank of Rio San Pedro, upstream from mouth (20 ppt salinity), seaward of Highway 180 bridge, village of Nuevo Campechito, Campeche, Mexico, 18°38.51'N; 92°27.88'W, sandy, organically rich, mud flat exposed at low tide among partially cleared red mangroves and stilted huts, 27 March 1991, D. L. Felder, J. L. Staton, R. B. Griffis, and T. L. Zimmerman; 2 males, vouchers for DNA analysis, ( ULLZ 5771), east side of Rio San Pedro, just inside river mouth (2.5 ppt salinity), seaward of Highway 180 bridge, village of Nuevo Campechito, Campeche, Mexico, 18°38.83'N; 92°27.91'W, muddy sand flat exposed at low tide, 7 July 2002, D. L. Felder and R. Robles. Mexico, Tabasco: 1 male, Puerto Ceiba, Tabasco, Mexico ( ULLZ 10294), 18°26.08'N; 93°09.69'W, clayey sand bottom <1 m deep off shoreline, small open area on mangrove shoreline, 5 July 2002, D. L. Felder and R. Robles; 1 male, 2 females, tissue vouchers for DNA analysis, Puerto Ceiba, Tabasco, Mexico ( ULLZ 5614), 18°25.65'N; 93°10.08'W, clayey sand bottom <1 m deep off mangrove lined shoreline, across estuary from refinery, 5 July 2002, D. L. Felder and R. Robles. Mexico, Veracruz: 1 male, major chela only ( ULLZ 10285), La Playa Escondida, Veracruz, Mexico, 18°35.63'N; 93°03.05'W, muddy sand near small stream on rocky intertidal beach, 4 July 2002, D. L. Felder, R. Robles, and I. T. Rodriguez; 4 males, 5 females, 2 ovigerous ( ULLZ 16269), shoreline of shallow back-beach pond on margin of tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44'W, muddy sand, 10 February 1999, D. L. Felder, S. C. Rabalais, C. D. Schubart, and C. F. Gurgel; 4 males, 1 female, vouchers for DNA analysis ( ULLZ 5194), shoreline of shallow back-beach tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44'W, muddy sand, 3 July 2002, D. L. Felder and R. Robles; 7 males, 18 females. 1 ovigerous, fragmented vouchers for allozyme analyses, abdomens removed ( ULLZ 16273), shoreline of shallow back-beach pond (2.5 – 3 ppt salinity) on margin of tidal stream west of Anton Lizardo, Veracruz, Mexico, 19°03.35'N; 96°00.44'W, muddy sand, 10 February 1999, D. L. Felder, S. C. Rabalais, C. D. Schubart, and C. F. Gurgel; 5 males, 6 females, fragmented vouchers for allozyme analyses ( ULLZ 16274), inlet to Laguna San Augustin seaward of bridge on Highway 180, Veracruz, Mexico, 19°55.27'N; 96°31.79'W, muddy sand flat at margins of shoreline mangroves and cordgrass, 9 February 1999, D. L. Felder, S. C. Rabalais, C. D. Schubart, and C. F. Gurgel.
Diagnosis. Rostrum acute, narrow, inclined dorsally as variably elevated arch, bearing dense ventral tuft of elongate setae extending from proximal half above eyestalks, flanked by low shoulders with apices slightly lateral to eyestalks. Dorsomesial margin of eyestalk thickened to form slight rounded swelling in distal 1/3. Inferior margin of cheliped merus with distinct single keel bearing proximal bifurcate hook and blunt denticles, superior margin with weakly developed proximal notch. Propodus of major chela with inferior margin terminating distally in sharply acute tooth, corner forming origin of sharp-edged carina extending vertically onto external face of article. Dactyl of male major chela prehensile margin with heavy subrectangular tooth in proximal half, separated by deep notch from elongate, narrow distal tooth serrated by rounded microdentition, superior margin proximally with short, strong, longitudinal carina. Third pereopod propodus inferodistal margin with small but defined median lobe between larger two. First abdominal tergite with dorsal sclerite forming broadly rounded lobe posteriorly. Second abdominal somite ventrally lacking pattern of small scattered sclerites and sclerotized median plate. Male first pleopod (gonopod) terminal article compressed, with narrow subterminal anterior margin, terminally bifurcate, distally drawn out to narrow acute tip. Appendix interna of male second pleopod large, setose, reaching beyond distal end of endopod. Second through fifth pleopods lacking distolateral spine on anterior face of basis. Telson broadly subrectangular, posterior margin weakly, if at all, trilobate. Uropodal endopod narrow, elongate subovoid, broadest proximal to midlength, length about twice breadth; triangular posteriorly directed acute tooth on protopod overreaching extended endopod by about 1/3 of endopod width.
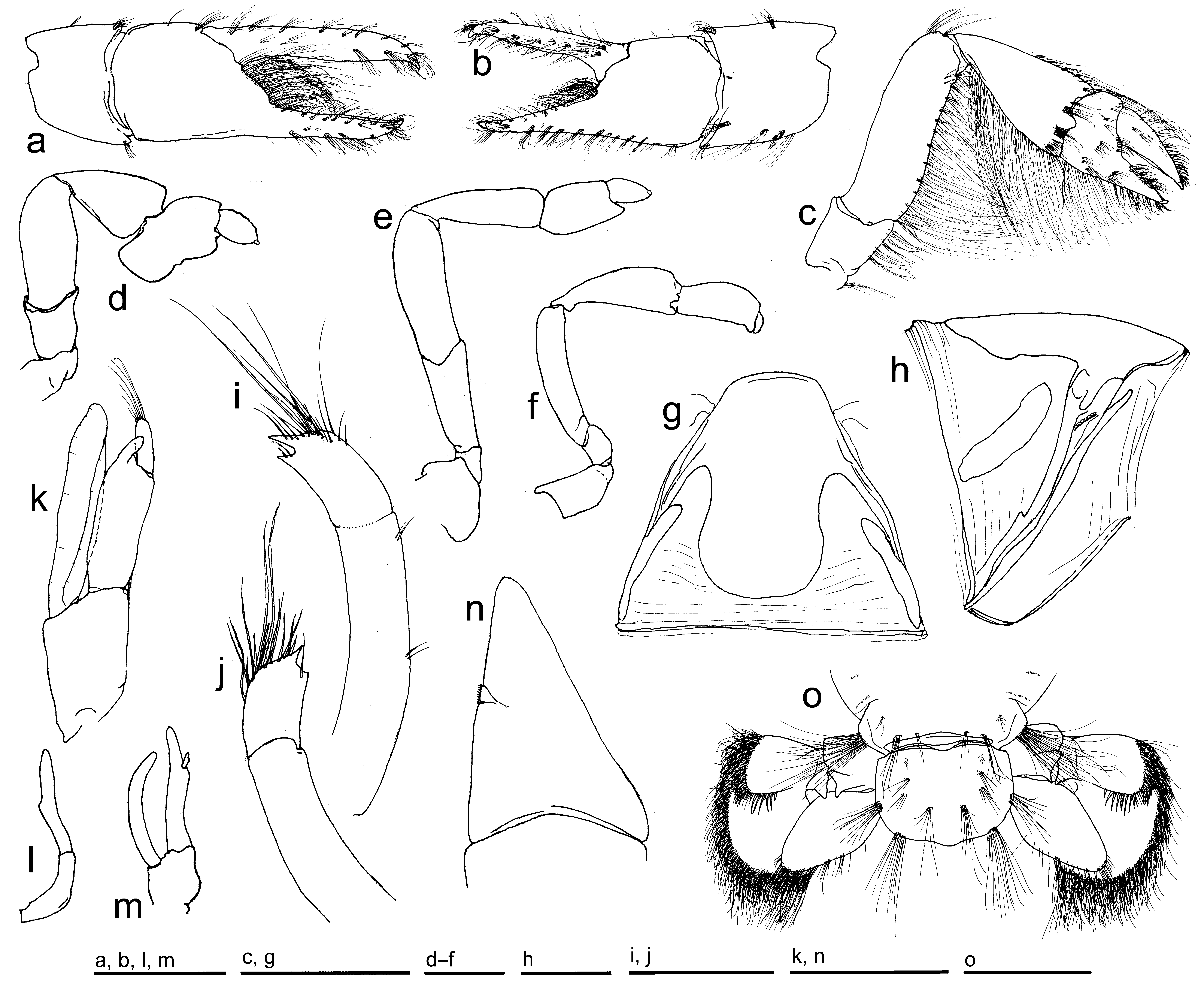
Description. Frontal margin of carapace with acute, narrow rostral spine flanked laterally by low, subangular rounded shoulders, apices of which are just lateral to eyestalks ( Fig. 1 View FIGURE 1 a); rostral spine slightly inclined dorsally, forming elevated arch above eyestalks in mature specimens ( Fig. 1 View FIGURE 1 b), extending 2/3 to 3/5 length of eyestalks in dorsal view, proximal half of rostral spine bearing ventral tuft of setae extending above and between proximal half of eyestalks. Carapace postrostrally with several pairs of short setose punctae to either side of midline; dorsal oval well defined, smooth, pair of widely separated setal punctae bearing 1 or 2 setae each near midlength, length of oval slightly less than 6/10 of postrostral carapace length; marginal suture of oval diminished at postrostral midline, stronger at posterior end with sclerotized articulation to bulbous cardiac region at posterior midline.
Eyestalks elongate subtriangular in dorsal view, reaching at least 3/4 length of basal article ( Fig. 1 View FIGURE 1 a); anterolateral margins distinctly sinuous, tapered to arcuate margin in distal half; dorsomesial margin thickened to form a slightly swollen boss in distal 1/3, ridge extending to subacute terminal protuberance of eyestalk ( Fig. 1 View FIGURE 1 b); distinctly faceted well defined cornea centered on dorsally, darkly pigmented area often exceeding faceted surface.
Antennular peduncle longer and heavier than antennal peduncle ( Fig. 1 View FIGURE 1 a); basal article dorsally invaginated to form statocyst occluded by setae, overlain by eyestalk; second article subequal to length of basal article, third article about 3 times length of second; second and third articles with dense ventromesial and ventrolateral rows of long, distoventrally directed setae. Antennular flagellum ventral ramus longer than third article of peduncle, with denser setation than dorsal ramus; dorsal ramus shorter than or subequal to third article of peduncle, heavier than ventral ramus in distal 1/3 where subterminal articles broader than those of ventral ramus, there fringed with short, dense ventral setae. Antennal peduncle reaching slightly beyond midlength of antennular peduncle third article; basal article dorsolateral carina arched to form lip above excretory pore, ventrally with setosedistal protuberance; second article with distal field of long setae on lateral boss; third article slightly shorter than combined lengths of first two, slightly shorter than fourth, ventrolaterally with broken line of long setae; fourth article narrower than others, setation limited to few long subterminal setae in single punctum. Antennal flagellum with sparse short setae, about 3 times length of antennular flagella.
Mandibular palp ( Fig. 1 View FIGURE 1 c) large, setose, 3-segmented, elongate third article robust, arched, subovoid; gnathal lobe of mandible subquadrate, with weakly angular distolateral shoulder, incisor process with well-defined corneous teeth on cutting margin, concave internal face with strong lip giving rise to molar process bearing weakly subdivided corneous tooth proximal and internal to incisor teeth; thin, rounded paragnath set against proximal convex face of molar process. First maxilla ( Fig. 1 View FIGURE 1 d) endopodal palp narrow, terminal article deflected at poorly defined articulation; proximal endite with dense, close-set setation lining sinuous mesial margin, setae spiniform distally; distal endite elongate, terminally broadened with dense long setation, several rows strongly spiniform; exopodite low, rounded. Second maxilla ( Fig. 1 View FIGURE 1 e) margins setose, endopod constricted to narrow terminus, first and second endites each longitudinally subdivided, exopod forming large, broad scaphognathite.
First maxilliped ( Fig. 1 View FIGURE 1 f) margins setose, endopod rudimentary, overlain by distal endite; blunt terminus of proximal endite coarsely setose; distal endite ovoid, narrowed distally, proximal 2/3 of external face with densely setose longitudinal elevation; exopod incompletely divided by oblique suture on external face, transverse suture complete on internal face except where abruptly deflected proximolaterally near margin to intersect lateral incision; margin lined by long setae, broadly arched proximal and distal to incision, mesial margin with comb of close-set very long setae, external face with dense field of mesially directed setae distal to oblique suture; epipod large, broad, anterior end strongly tapered. Second maxilliped ( Fig. 1 View FIGURE 1 g) margins of both rami heavily setose, endopodal merus and propodus arcuate, flexor margin of merus with comb of long setae, internal face produced distally to form rounded marginally setose lobe extending over internal proximal margin of short carpus; merus length about 4 times width; propodus length exceeding 2/3 length of merus, longest setae originating on extensor margin and distal half of external face; dactylus twice as long as broad, rounded terminally, distal half bearing short stiff cornified setae; exopod broad, bladelike, narrowing distally, distinctly overreaching endopodal merus, arcuate, terminally rounded; bilobed epipod originating from peduncle with short, truncate basal lobe, elongate narrowly tapered, distal lobe. Third maxilliped ( Fig. 1 View FIGURE 1 h) with small, terminally subacute, rudimentary exopod; endopod large, setose; endopodal ischium subrectangular, maximum diagonal length slightly less than 2 times width at midlength, internal face with rudimentary unarmed to weakly tuberculate or crenulate longitudinal carina on proximal half; merus short, subrhomboidal, broader than long, mesial margin distinctly arcuate; carpus subrectangular, longer than broad, internally with dense field of stiff setae in distal third; propodus large, subovoid to subrhomboidal, longer than broad, internally with dense field of stiff setae on proximal third, externally with low, setose, angular lobe on flexor margin immediately below articulation of dactylus, demarcated from inferior margin by slight offset incision extended onto external face; dactylus narrow, arcuate proximally, long setae of extensor and distal margins including a few long stiff bristles at terminus.
Branchial formula as reported for congeners (Felder & Staton, 2000); endopods and epipods as previously described ( Felder & Rodrigues, 1993), branchiae limited to single rudimentary arthrobranch on second maxilliped, pair of arthrobranchs on third maxilliped, and pair of arthrobranchs on each of the first through fourth pereopods. Major cheliped located on either right or left side, shape and ornamentation sexually dimorphic.
Major cheliped of male ( Fig. 1 View FIGURE 1 i, j; Fig. 3 View FIGURE 3 a–c, d) massive, fingers heavily armed; ischium slender, superior margin sinuous, inferior (flexor) marginal carina armed by very small denticles on proximal 1/2 to 2/3, usually disjunct from 2–5 much larger and longer subacute, straight teeth on distal elbow, some of which may be compound; merus with distinct, shallow depression or notch in proximal 1/4 of superior margin, inferior (flexor) marginal keel arcuate distally, evidence of bicarinate keel obsolete, at most marked by slight furrow to internal side of inferior margin, terminally bifurcate proximal hook at external base of keel, hook terminating in spiniform or dentiform tip usually paired with weak subterminal tip or lobe, 3–5 small blunt denticles originating very slightly to external side of keel margin, largest of which at or distal to most arcuate part near midlength; carpus broad, subquadrate, superior and inferior margins keeled, near parallel in distal half, terminated distally in subacutely angular superior comer, carpus with inferodistal corner terminated in single sharply acute tooth, corner forming origin of sharp-edged, sometimes crenulate carina extending short distance vertically from this corner onto external face of article (usually 1/4 to 1/3 of propodus height); propodus broad, heavy, length of fixed finger about 1/2 length of palm; internal face of palm proximally smooth, with rounded swollen boss centered near midline, distally with broadly rounded carina extending proximally from internal prehensile margin of fixed finger below smooth depression proximal to gape of fingers and above elongate concavity extending almost full length of article; external face with weak carinae defining slight depression extending proximally from gape of fingers, in which is centered large setal punctum; slightly sinuous keel of superior propodal margin restricted to proximal 2/3, keel of inferior margin distinct full length of palm, becoming obsolete on fixed finger where broken by large setose punctae; fixed finger originating below broadly subtriangular heavy tooth at proximal end of gape, separated by broad U-shaped notch from strong, distally inclining, triangular tooth originating from external prehensile margin, centered near 1/3 to 2/5 length of fixed finger; fixed finger with well-defined separation of internal and armed, elevated, external prehensile margin, internal margin unarmed but forming thick rounded carina proximally extending slightly onto palm; dactylus with hooked tip, superior margin forming short, elevated, longitudinally oriented proximal carina decreasing in elevation distally, internal face with strong rounded proximal carina, inferior shoulder forming unarmed, poorly defined internal prehensile margin, external prehensile margin usually with 3 heavy, prehensile teeth, proximal tooth a broad, low transversely bilobate molar evident only with dactylus fully extended (sometimes weakly defined), middle tooth heavy, subtruncate, slightly inclined distally, usually roughened by microtubercles on proximal or distal surfaces, originating near 1/3 length of finger, separated by Ushaped notch from narrow elongate bladelike distal tooth extending from near midlength of dactyl almost to terminal hook, elevated proximal prominence of distal tooth thickest, distally sloping prehensile edge armed by uniformly spaced series of small, heavy, rounded tubercles along immediate internal side of tooth margin.
Major cheliped of female ( Fig. 1 View FIGURE 1 k, Fig. 3 View FIGURE 3 e, f) less massive, less strongly armed and sculpted than that of male; prehensile margins on fixed finger and most of dactylus evenly armed by serration of low rounded denticles, dactylus narrow, relatively less massive proximally than fixed finger; fixed finger proximally broader vertically than in males, notch at base of fixed finger a narrow V-shaped incision proximal to small denticular serration of prehensile margin, lacking distinct development of prehensile tooth, conspicuously concave between internal and external prehensile margins; superior and inferior margins of propodus convergent distally, laterally compressed to form narrow dorsal and ventral keel; tip of dactylus usually overreaching tip of fixed finger when crossed, gape filled including incisional notch at base of fixed finger.
Minor cheliped of male ( Fig. 2 View FIGURE 2 a) sparsely armed; ischium weakly serrate by small denticles on most of flexor margin, denticles more widely separated into microtubercles distally; merus mostly unarmed, inferior margin usually with 1 small tubercle proximally; carpus with acute distal comers, rounded carina extending vertically from corner, forming subterminal edge of lower external face; fixed finger narrower than dactyl, less hooked distally, prehensile margin unarmed, sloping proximal 1/3 with broad field of dense elongate setae filling about half of gape length and extending somewhat onto internal face; dactylus prehensile margin weakly armed, limited to few weak tubercles above setae filling gape and weak evidence of subterminal tooth as broad swelling, each finger terminating in acute to subacute cornified tip. Minor cheliped of female ( Fig. 2 View FIGURE 2 b) with dense, elongate prehensile setation of fixed finger much less developed than in males, gape somewhat narrower between fingers.
Second pereopod ( Fig. 2 View FIGURE 2 c) chelate, flexor margins of merus and carpus lined with evenly spaced long setae, inferior margin of propodus with long setae proximally, grading distally to dense field of short, hooked bristles; middle 1/3 of fixed finger with patch of short stiff bristles just outside prehensile margin; superior margin of dactylus with proximal long setae grading to short stiff hooked bristles distally.
Third pereopod ( Fig. 2 View FIGURE 2 d) merus length about 2.2 times width; propodus with inferodistal margin trilobate, middle lobe small, between major distal and proximal lobes, setal tufts giving margin weakly scalloped appearance, lobes demarcated by furrows on internal face, distal margins of lobes with dense fringe of tufted setae concealing shorter corneous bristles, upper lobe with at least one heavier, weakly hooked, distally directed corneous tooth arising from margin, longest setae on inferior margin of lower lobe, patterned tufts of lighter setae on external face of article; dactylus tear-shaped, superior margin concealed by long dense setae on external face, margin internally evident as distinctly sinuous, article terminating in short, narrow, ventrolaterally directed corneous tooth, external face with fields of setae and shorter stiff bristles lining inferior margin, grading to fields and poorly defined rows of much finer setae above. Fourth pereopod ( Fig. 2 View FIGURE 2 e) weakly subchelate, inferodistal process of propodus (= fixed finger) forming subacutely angular, terminally cornified lobe extending distally no more than 1/3 length of dactylus, inferior margin of lobe with one or more very heavy, hooked, subterminal corneous spines, margin proximally with additional lighter short bristles, obscured by dense brush of setae, setae originating from inferior margin and lower internal surface strongly serrate; dactylus elongate, tear-shape, superior margin arched, terminating in short ventrolaterally directed corneous tooth. Fifth pereopod ( Fig. 2 View FIGURE 2 f) minutely chelate, opposable surfaces of fingers spooned, terminally rounded; propodus with dense field of long, close-set setae on internal face, fixed finger deflected, terminally pectinate; dactylus terminally narrower than fixed finger, hooked form of beak-like chela obscured by dense setation on distal 2/3 of propodus and superior face of dactylus.
Abdominal somites mostly smooth dorsally. First abdominal tergite ( Fig. 2 View FIGURE 2 g, h) with slightly thickened, translucent, mid-dorsal sclerite articulating anterolaterally to narrow anterolateral sclerites, forming broad medially centered rounded lobe posteriorly, separated from posteriorly diverging lateral sclerites by broad membranous area to either side; membranous area surrounding single, large, elongate posterolateral sclerite to either side, membrane lacking embedded small scattered sclerites, Second tergite posterolateral lobe below suture sclerotized like remainder of tergite, lobe crossed by well-defined sinuous anterior tranverse line of very fine setae, 2 shorter less defined oblique lines in posterior half; ventrolateral lobe of tergite centered anterior to midlength of tergite. Third to fifth tergites each encompassing a finely setose, lateral, membranous subcircular or suboval area below a weak posterolateral suture, those of third and fourth tergite more posterolaterally positioned than on fifth, that of fifth tergite smallest, most circular. Sixth tergite with 2 transverse posterolateral lines of short setae anterior to posterolateral groove from which transverse and posterior sutures originate, longest line adjacent, subparallel to transverse suture; posterolateral margin of tergite marked by typically 3 closely parallel longitudinal lines of micropunctae bearing very fine setae; transverse suture ill-defined across most of sixth tergite ( Fig. 2 View FIGURE 2 o), lacking clear branch to short longitudinal posterior suture; tufts of stiff setae mesial to each posterior suture, on posterolateral corners, and usually as 4 short lines or tufts of stiff setae on posterior margin.
Ventral surfaces of abdominal somites generally lacking extensive armor of sclerotized plates and tubercles ( Fig. 3 View FIGURE 3 c); in male, heaviest sclerites of first somite forming narrow convergent furrows extending anteromesially from base of each pleopod, furrows roughly accommodating first pleopods when fully flexed against abdomen; same sclerites in female much weaker, furrow very narrow; second somite in both sexes lacking evidence of longitudinally furrowed median plate and thicken small sclerites forming tubercles embedded in ventral abdominal membranes.
First pleopod of male and female uniramous, composed of 2 articles; in male, originating from atop external of two sclerotized ridges forming narrow trough or channel to rough accommodate flexed pleopod; appendage length slightly less than 1/2 that of second pleopod, proximal article more than 2 times length of terminal article ( Fig. 2 View FIGURE 2 i, j), terminal article compressed, subspatulate anterodistally, concave anterior edge very narrow, convex subterminal margin less so, bearing long setae, terminally bifurcate, forming distal and subdistal acutely terminated blades, weakly separated in large individuals, anterolaterally directed of distal blade usually drawn out to narrow acute tip, cornified; in female ( Fig. 2 View FIGURE 2 l), extended length subequal to that of second pleopod, proximal article bearing broad setose lobe near midlength, slightly shorter than terminal article, terminal article forming narrow spatulate blade beyond setose lobe at midlength. Second pleopod of male and female biramous, with appendix interna on endopod, both rami setose; in male ( Fig. 2 View FIGURE 2 k), appendix interna large, overreaching terminal lobe of endopod, long terminal setae subequal to appendix length, overreaching exopod; in female ( Fig. 2 View FIGURE 2 m), appendix interna small, narrow, markedly overreached by terminus of endopod, terminally subacute, terminus and mesial subterminal shoulder with field of minute, rudimentary hooked setae.
Second through fifth pleopods with, at most, a low tubercle or tooth on anterior face of basis at condylar articulation with exopod. Third to fifth pleopod pairs forming large, posteriorly cupped fans when crosslinked by hooked setae of appendices internae on opposed margins of endopods; endopod of each subtriangular ( Fig. 2 View FIGURE 2 n), stubby appendix interna demarcated by sutures where embedded in endopod, offset very slightly from mesial margin of endopod.
Telson ( Fig. 2 View FIGURE 2 o) broad, subrectangular, width about 1.5 times length, broadest near midlength, posterior margin at most very weakly trilobate, only median lobe consistently evident; dorsally with 3 pairs of strong setal tufts, anterior to which is another pair of microsetae in short longitudinal row of micropunctae, posteriormost pair of dorsal tufts set nearest midline in posterior half of telson, two anterior pairs and lines of micropunctae roughly aligned longitudinally well lateral of midline, spaced anteriorly from midlength lateral margins with pair of strong setal tufts just posterior to midlength, posterior margin well developed tufts posterolateral extreme. Uropod ( Fig. 2 View FIGURE 2 o) with strong, acutely triangular, posteriorly directed tooth on protopod and similar but usually heavier tooth on proximal article of exopod, both positioned to abut or overreach anterior margin of extended endopod, that on protopod usually sharply acute and overreaching extended endopod by about 1/3 of endopod width; endopod elongate, subovoid, about twice as long as broad, posteromesial margin with broken fringe of setae, tapering to rounded terminus bearing marginal fringe of long setae, distal end distinctly overreaching distal end of anterodorsal plate on flexed exopod; exopod anterodorsal plate falling well short of distal margin, posterodistal edge of plate with short, thick, spiniform setae grading to thinner, dense, elongate setae of exopod margin; distal margin of exopod with dense fringe of setation, longest posteriorly.
Color. In life, color of this species usually includes a small area of rose-violet to diffuse lavender brown positioned to either side (anterolateral) of the anterior dorsal oval, filling an area marked by a slight depression ( Fig. 3 View FIGURE 3 b, d-h). This pigmentation appears to be subtegumental and is darkest posteriorly. No similar area has been found in fresh specimens of regional congeners, making this a useful character for recognition of fresh specimens, though it was very faint in one specimen taken at Puerto Ceiba, Tabasco. Faint dorsal color coloration on the abdomen ( Fig. 3 View FIGURE 3 b–f) is of similar pattern to that in congeners with consistently dark yellow to yellow-orange or brown chromatophores. No examples were seen in which dorsal chromatophores of the abdomen were of the rose red to violet color often seen in fresh specimens of L. louisianensis .
Size. Maximum observed postorbital carapace length of male 14.9 mm; of female 15.5 mm. Egg length ranges from about 1.0– 1.4 mm in preserved ovigerous females, the largest being immediate pre-hatch; in life, immediate prehatch eggs measured 1.4–1.5 mm.
Habitat. These being obligate infaunal animals, all were collected with hand-operated “yabby” suction pumps from intertidal to shallow subtidal substrates including muddy quartzite sands, clayey muds, and gravelly coarse sands layered with mud, all usually with high detrital organic content. Settings ranged from back-beach ephemeral salt ponds behind sandy open Gulf shorelines to estuarine habitats that included mud banks, subtidal bars, and mangrove or cordgrass lined shorelines in euryhaline lagoons and river mouths. In most sampled locations, this species occurred together with Lepidophthalmus manningi . In those habitats near mangroves, the burrows of both species usually occurred alongside those of the axianassid Axianassa australis Rodrigues & Shimizu, 1992 .
Distribution. Western Atlantic: Southwestern Gulf of Mexico (protected shorelines, margins and mudflats of inlets, lagoons, tidal streams, and estuaries of Veracruz, Tabasco, and Campeche, Mexico).
Type locality. Sandy mud exposed at low tide among partially cleared red mangroves and stilted huts, east bank of Rio San Pedro, just upstream from mouth and seaward of Highway 180 bridge, village of Nuevo Campechito (18°34.85'N; 92°27.87'W), Campeche, Mexico,
Remarks. Lepidophthalmus statoni sp. nov. can be distinguished from the western Atlantic species L. jamaicense ( Schmitt, 1935) , L. manningi , L. richardi Felder & Manning, 1997, and L. sinuensis Felder & Rodrigues, 1993 , by its lacking a thickened ventral median sclerite and associated small sclerites on the second abdominal somite (see Felder & Manning, 1997; Felder & Staton, 2000). In this character it also differs from the eastern Pacific species L. bocourti ( A. Milne-Edwards, 1870) and L. eiseni Holmes, 1904 , both of which bear welldeveloped ventral abdominal sclerites (Felder, 2003). The strength of this ventral armor, along with its shape and augmentation by surrounding small sclerites, varies somewhat among the plated species, as well as with ontogeny, but it is consistently evident on close study. Among western Atlantic ventrally armored species, the median ventral plate of the second abdominal somite may be at times weakly defined as little more than a closed median suture in L. sinuensis , but that southwestern Caribbean species is readily separable by its striking projections on the carapace frontal margin to either side of the eyestalks ( Lemaitre & Rodrigues, 1991).
Only two other known western Atlantic species of the genus lack such median ventral armor, one being L. siriboia Felder & Rodrigues, 1993 , from Brazil and the other L. louisianensis ( Schmitt, 1935) from the northern to northwestern Gulf of Mexico. Of these two, L. statoni sp. nov. most closely resembles L. louisianaensis , sharing with it a male major cheliped in which the keel along the inferior margin is primarily single, rather than distinctly bicarinate over most of its length as in L. siriboia , and a superior margin that proximally bears a comparatively weak notch as opposed to the strong notch in the Brazilian species. Again in L. louisianaensis and L. statoni sp. nov., the male chelipeds are much more similar in proportions and shapes of the carpus and propodus, as well as finger dentition, than either is to the male cheliped of L. siriboia (see Felder & Rodrigues, 1993: fig. 1 d; 4 a, c, d),. Mature males of these two species also share a male first pleopod in which the terminal and subterminal blades are less strongly developed than in L. siriboia , the terminal blade being narrower and the incision between the two blades being shallower than in the Brazilian species (see Felder & Rodrigues, 1993: fig. 1 g, h; 3 d – f; 6 c – h). Also in mature males of the two Gulf of Mexico species, the appendix interna of the second pleopod overreaches the terminus of endopod, to varied degrees being crossed with it, as opposed to neither exceeding nor crossing it as in L. siriboia (see Felder & Rodrigues, 1993: fig. 3 h; 6 j). Finally, present examination of specimens assigned to L. siriboia (ULLZ 5611) shows the first abdominal somite to have a broad posteriorly lobate mid-dorsal sclerite with narrow posterolateral membraneous areas to either side, but with no evidence of a posterolateral sclerite within the membraneous area. The posterolateral sclerite, much as illustrated here for L. statoni sp. nov. ( Fig. 2 View FIGURE 2 g, h), is also present in L. louisianensis .
Both L. louisianensis and L. statoni sp. nov. are endemic to the Gulf of Mexico, though their ranges are allopatric. On the basis of present records, L. louisianensis ranges no further south in the western Gulf of Mexico than Laguna Tamihaua,Veracruz, Mexico, and L. statoni sp. nov. replaces it just to the south of that, ranging at least as far north as Laguna San Augustin, Veracruz, Mexico. Referred to at the time as Lepidophthalmus sp. “b”, Staton et al. (2000) showed in allozyme analyses the close genetic proximity of these two species, though this analysis did not robustly represent members of the genus. Pending sequence-based molecular phylogenetic analyses (Robles & Felder, in review) do however address the genus broadly, and continue to show these two species as paired sister species. Discussion of possible evolutionary processes that separated them is deferred to that analyses, but characters can be compared for separation of these two apparently close relatives.
In both sexes of L. statoni sp. nov., though most strongly developed in large males, the inferior margin on the major cheliped propodus terminates distally in sharply acute tooth, with this corner forming the origin of a sharpedged carina extending vertically onto the external face of the article ( Fig. 1 View FIGURE 1 i–k). While a similarly positioned carina is found in L. louisianensis , its margin is dull or rounded, less undercut, and usually not extended as far onto the external face of the propodus. Also in L. statoni sp. nov. the dactyl of the major chela typically bears a short elongate elevated ridge proximally on the superior margin, rather than a roughly rounded tubercle as in L. louisianensis , and the prehensile margin bears a distinctly subquadrate tooth in the proximal half, which is more commonly irregularly rounded to subtriangular in L. louisianensis . Among differences in other pereopods, the inferodistal margin of the third pereopod propodus is trilobate in L. statoni sp. nov., with a small median lobe being defined between the two larger, though this is not usually evident in L. louisianensis . Finally, the first and second male pleopods differ between mature individuals of the two species, the terminal blade of the first pleopod in L. statoni sp. nov. being in most cases more narrowly drawn out to a fine, narrow spiniform tip than in L. louisianensis , and the appendix interna of the second being more strongly crossed with the narrow endopod tip than in L. louisianensis .
Other than the western Atlantic species, there are at least three eastern Pacific species known to lack well developed ventral abdominal sclerites, all of which occur along the coasts of Central America. Only one of these, Lepidophthalmus rafai Felder & Manning, 1998 , has been named, the others having been long under study, as noted in numerous previous papers ( Felder & Rodrigues, 1993; Felder & Manning, 1998; Felder & Staton, 2000; Felder, 2003; Felder et al. 2003). Two of these undescribed species, both of which bear strongly trilobate posterior margins on their telsons, are treated further in a coming molecular phylogenetic analyses (Robles & Felder, in review) deemed essential to establishing their somewhat unexpected relationships among the eastern Pacific species before offering formal descriptions.
At all five of the collecting sites from which L statoni sp. nov. was taken, it occurred sympatrically with its ventrally plated congener Lepidophthalmus manningi and the axianassid Axianassa australis . On several occasions specimens of pink wormfish (Microdesmus sp.) and commensal alpheid shrimp ( Leptalpheus spp.) were also taken from burrows possibly assignable any one of these sympatric species. Ovigerous specimens of L. statoni sp. nov. occurred at most sites from which collections were made in March 1991 and February 1999, and the wormfish are now known predators on the eggs being carried by burrowed ovigerous females of Lepidophthalmus ( Felder & Rodrigues, 1993; R. W. Heard, personal observations in aquaria). The specimens of Leptalpheus appear to include Leptalpheus forceps Williams, 1965 , almost certainly from the burrow of L. statoni sp. nov., and one or two undescribed species that cannot with certainty be assocated to specific hosts.
Etymology. This species is named for Joseph L. Staton, whose work was essential to recognition of this cryptic species. In the mid-1990’s, Joe was the first doctoral student in my lab to apply genetic methods to studies of regional decapods, and much was built on what he started.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.