
Meristogenys dyscritus, Shimada, Tomohiko, Matsui, Masafumi, Yambun, Paul & Sudin, Ahmad, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.205593 |
|
DOI |
https://doi.org/10.5281/zenodo.5613136 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0B522-FFD5-4F48-FF56-90C6FF021AD4 |
|
treatment provided by |
Plazi |
|
scientific name |
Meristogenys dyscritus |
| status |
sp. nov. |
Meristogenys dyscritus View in CoL sp. nov.
Amolops sp. A: Matsui, 1979, p. 340, figs. 26, 28A; Amolops amoropalamus: Matsui, 1986 , p. 628–629 (part); Meristogenys amoropalamus (lineage 3, lineage 4): Shimada et al., 2007a, p. 173–189; Shimada et al., 2008, p. 24–34; Shimada et al., 2011, p. 158–177, Fig. 3–4 View FIGURE 3 View FIGURE 4 , Appendix 2, Meristogenys amoropalamus (larval morphotype 3-a): Shimada et al., 2007b, p. 59–63.
Diagnosis. This species is a member of the genus Meristogenys , based on the morphology of the larval specimens molecularly assigned to it (larval morphotype 3-a in Shimada et al., 2007a); these had diagnostic characteristics of the genus: abdominal sucker; divided upper jaw sheaths; and ribs on the outer surface of both jaw sheaths. Meristogenys dyscritus is a small species of the M. jerboa species group (Matsui, 1986), with male SVL 31.7–39.5 mm, female SVL 61.0– 71.2 mm; rear of thigh dark brown, dusted with small irregular light spots; web poorly developed leaving terminal two phalanges on fourth toe free of broad webbing; ova relatively small and heavily pigmented in the animal pole; upper third of iris sandy brown to yellowish brown.
Etymology. The specific name is derived from dyskritos (Gr.), doubtful or indistinguishable, referring that the new species is difficult to distinguish from M. amoropalamus in adult morphology.
Holotype. Osaka Museum of Natural History ( OMNH) Am8069, an adult male from Liwagu River, 1500 m, Headquarters of Kinabalu National Park, Sabah, Malaysia. Collected in March, 1979, by Masafumi Matsui.
Paratypes. Two males and a female from the type locality; Zoologisches Forschungsinstitut und Museum A. Koenig, Bonn ( ZFMK) 41644; Sabah Parks (SP) 26481, 21392.
Referred specimens. Forty-five males and 16 females from the type locality (Liwagu), Mahua, and Wario (See Appendix).
Description of Holotype (measurements in mm). Body moderately slender, SVL 31.7; head subtriangular, slightly longer (12.8) than wide (12.7); snout blunt, projecting slightly beyond lower jaw; eyes elevated; canthi sharp, slightly concave; lores slightly oblique, concave; nostrils lateral, just below canthal edge, distinctly closer to tip of snout (2.4) than to eye (2.7); IND (4.3) wider than IOD (3.0); latter narrower than UEW (3.4); SL 5.2; pineal spot visible, slightly behind the line connecting anterior corners of orbits; tympanum distinct, TDv (3.5) and TDh (3.4), two-thirds of EL (5.2); T-EL (0.8) one-fourth of TDv and TDh; vomerine teeth obvious, in small oblique groups separated by the width of one group, groups on line connecting rear rims of choanae; tongue deeply notched, without papilla; paired subgular vocal sacs form gular pouches at corners of throat; vocal opening just inside commissures of jaws.
Fingers slender, first and second subequal, much shorter than third; tips expanded into disks having circummarginal grooves; disks of all fingers subequal in diameter, one-third diameter of tympanum; no fringes of skin along fingers; no supernumerary metacarpal tubercles; distinct nuptial pads covering dorsal and medial surfaces of the first finger from its base to subarticular tubercle.
Forelimb length unavailable; LAL 19.1; HAL 11.0; HLL 69.7; tibia long (23.1); heels overlapping when limbs are held at right angles to body; THIGH (19.9) and FL (20.3) much shorter than TL. Toe disks similar to those of fingers in shape and size; webbing not fully extending to disk on fourth toe, leaving two phalanges free of broad sheet; excision of web between fourth and fifth toes proximal to middle subarticular tubercle of fourth toe; a narrow fringe of skin along medial edge of first toe; inner metatarsal tubercle elliptical, shorter than distance between it and subarticular tubercle of first toe; a small round, raised outer metatarsal tubercle.
Skin of dorsum finely granular on head and trunk; a weak fold from above eye to axilla; a low dorsolateral glandular fold; side of trunk coarsely granular; limbs strongly rugose above; throat smooth; chest and abdomen weakly rugose.
Colour in life (See coloured pictures of Fig. 7 View FIGURE 7 ). Dorsum light brown with small dark spots; lores with dark streak below canthus; upper lip light grey; a blackish brown band beginning behind eye bordering rear of the tympanum, diverging above the tympanum and nearly reaching the inguinal area, dorsal and ventral boundaries sharp; limbs marked dorsally with alternating light and dark brown crossbars, the darker ones narrower; a short dark streak ventrally at insertion of arm; rear of thigh light brown with scattered irregular light spots; throat and chest whitish with dots of melanophores; lower lip vaguely barred with blackish brown; ventral surface of tibia whitish, with heavy pigmentation of melanophores.
Iris colour. Although iris colour of the holotype is unavailable, other specimens from the type locality (Liwagu) had the bicoloured iris, sandy brown above and reddish brown below.
Colour in alcohol. Color pattern has not been changed even after preservation in ethanol for several years, except for iris colour which disappeared soon after the fixation in formalin solution.
Eggs: Ova bicoloured, heavy pigmentations on the animal pole, cream white on the vegetal pole; clutch size 750–1500; ovum diameter 1.6–1.8 mm.
Larvae (See coloured pictures of Fig. 8 View FIGURE 8 ): We examined 67 specimens of stage 26–41 of Gosner (1960) from Mahua. These specimens are identical to those used by Shimada et al. (2007a) to define their larval morphotype 3- a.
Head-body length (HBL) 10.0– 16.8 mm. Head-body oval, broadly rounded at snout, flat below, head-body width (HBW) maximum at level of spiracle 61–71% (median=65.8%) of HBL; depth 49–64% (median=55.4%) of HBW; eyes dorsolateral, not visible from below, pointing outward, eyeball 13–15% (median=13.7%) of HBL; interorbital 123–169% (median=147.6%) of eye diameter; eye-snout distance 36–46% (median=41.1%) of HBL; nostril open, rim not raised, closer to eye than to tip of snout; internarial 57–80% (median=69.8%) of interorbital.
Oral disk ventral, width 57–73% (median=65.7%) of HBW; upper lip separated from snout by a groove; upper lip with short marginal papillae in lateral third, inframarginal papillae near corner; lower lip with uninterrupted row of short marginal papillae; labial tooth row formulae 6(4–6)/6(1) in three specimens, 6(4–6)/7(1) in 48 specimens, and 6(4–6)/8(1) in 13 specimens; jaw sheaths heavy completely black except for outer margins covered by thin film; upper sheath film thicker than the lower; outer surface of lower jaw sheaths with several weak ribs; margin finely serrate, 3–11 and 4–7 serrae on a half of upper and lower jaw sheaths, respectively; upper jaw sheaths Mshaped, lower V-shaped; both jaw sheaths divided; a large suctorial abdominal disk following oral disk; peripheral part of disk darkened and keratinized; length 40–52% (median=46.5%) of HBL; width 78–98% (median=87.9%) of HBW.
Spiracle sinistral; tube moderately long, length subequal to length of eyeball, pointing upward and backward, free of body wall for half its length; anal tube median, free of tail; tail heavily muscled, dorsal margin strongly convex, deepest before middle, tapering to slightly pointed tip; tail length 143–179% (median=164.7%) of HBL, maximum depth 23–33% (median=26.5%) of length; caudal muscle deeper than fins in basal half; dorsal fin origin behind body, fin deeper than ventral fin except in final fourth; ventral fin origin at end of proximal third of tail; head-body with four pairs of glandular clusters; a postorbital cluster about an eye length behind eye, with 0–3 glands; a infraorbital at the base of snout, with 1–3 glands; a prespiracular cluster just anterior to spiracle, with 0–4 glands; a midlateral at the posterior end of body, with 0–8 glands; no ventral and dorsal fin glands; 0–16 ventral fin glands; head-body scattered dorsally with minute protuberances anterior to eye in developed larvae; the area occupied by spinules and their density increasing with stage of development; lateral line pores indistinct.
Head-body light brown dorsally and laterally, sometimes posterior half of lateral surface dark brown; caudal muscle light brown; translucent fins with scattered pigmentations.
Other ten larval specimens from Bundu Tuhan (st. 28), Liwagu (st. 32), Poring (st. 31), and Wario (st. 30 and 32) had characteristics similar to Mahua specimens, but all ten specimens had six rows of lower labial teeth.
Variation. Males from Mahua tended to be smaller than males from Liwagu and Wario in SVL ( Table 3 View TABLE 3 ).
Although relatively weak web development is one of the diagnostic characters of this species, some females from Liwagu and Wario had developed broad webs, which reached to the toe disk. Individuals usually had a bicoloured iris, sandy brown above and reddish brown below, but some females from Liwagu had monocoloured reddish brown iris.
Larvae from Mt. Kinabalu tended to have fewer labial tooth rows in lower jaw than the specimens from Mahua (Crocker Range). In Mahua, only 4.7% of larvae had six rows of labial teeth on the lower jaw, and all other larvae had seven or eight rows, whereas all ten larvae collected around Mt. Kinabalu (Bundu Tuhan, Liwagu, Poring and Wario) had six rows.
Calls. We recorded calls of a male of M. dyscritus (SP 26472) at Liwagu at the midnight of 14 June 2007 ( Fig. 9 View FIGURE 9 ) at an air temperature of 17.5 C. The male perched on a low shrub, 50 cm above ground, and 1 m from the river. Many other males were also calling around there.
The call was emitted sporadically with several minutes intervals, and consisted of a short, unpulsed note, lasting about 0.11 sec. A call included three continuous phases, of which the first one had a marked frequency modulation; it descended quickly in frequency from 8000–10000 Hz down to about 6000 Hz. In the second phase, the frequency was kept stable around 6000 Hz. The third phase had a remarkable frequency modulation again; it ascended quickly in frequency from 6000 up to 9000 Hz. Dominant frequencies were traced at about 6000–10000 Hz ranges, and harmonic bands were often found between 12000–15000 Hz ranges.
Comparisons. Among 11 known species of Meristogenys , M. kinabaluensis (Inger, 1966) , M. poecilus (Inger and Gritis, 1983) , M. stenocephalus Shimada et al. 2011 , M. stigmachilus Shimada et al. 2011 , and M. whiteheadi (Boulenger, 1887) have a large body size (male SVL> 45 mm, female SVL> 70 mm) and are easily distinguished from M. dyscritus . From some small species, M. dyscritus is differentiated in the poorly developed toe webs [In M. jerboa (Gunther, 1872) , M. macrophthalmus (Matsui, 1986) , M. maryatiae Matsui et al., 2008 , M. orphnocnemis (Matsui, 1986) , and M. phaeomerus (Inger and Gritis, 1983) , webs reach at the toe disk], heavy pigmentations on the ventral surface of tibia ( M. jerboa , M. maryatiae , M. orphnocnemis , and M. phaeomerus have only scattered pigmentation), small irregular light spots on the rear of thigh ( M. jerboa and M. macrophthalmus have large bright blotches), and black streaks around eyes, lores and tympanum ( M. jerboa , M. maryatiae , M. orphnocnemis , and M. phaeomerus usually lack such colour patterns).
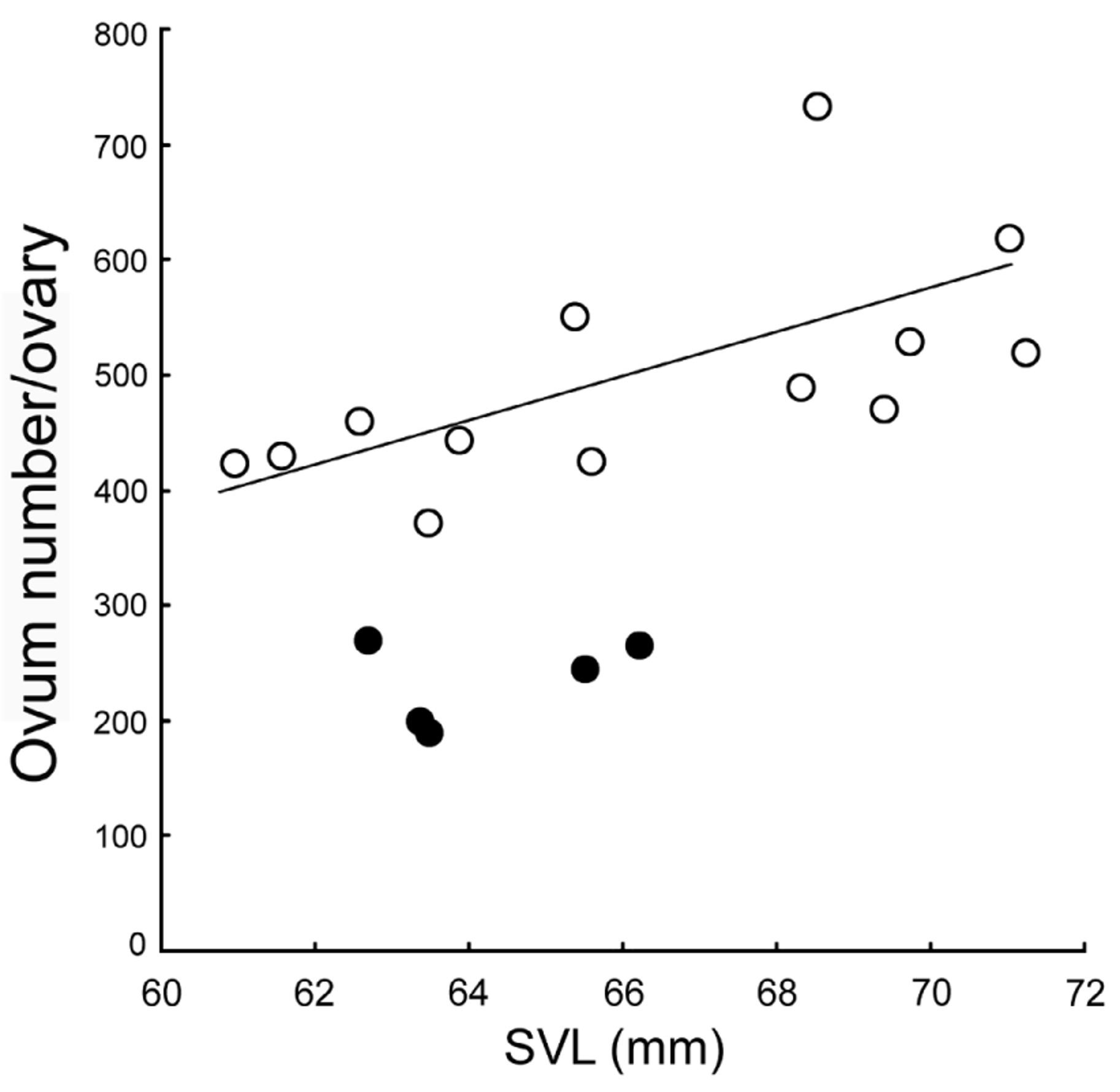
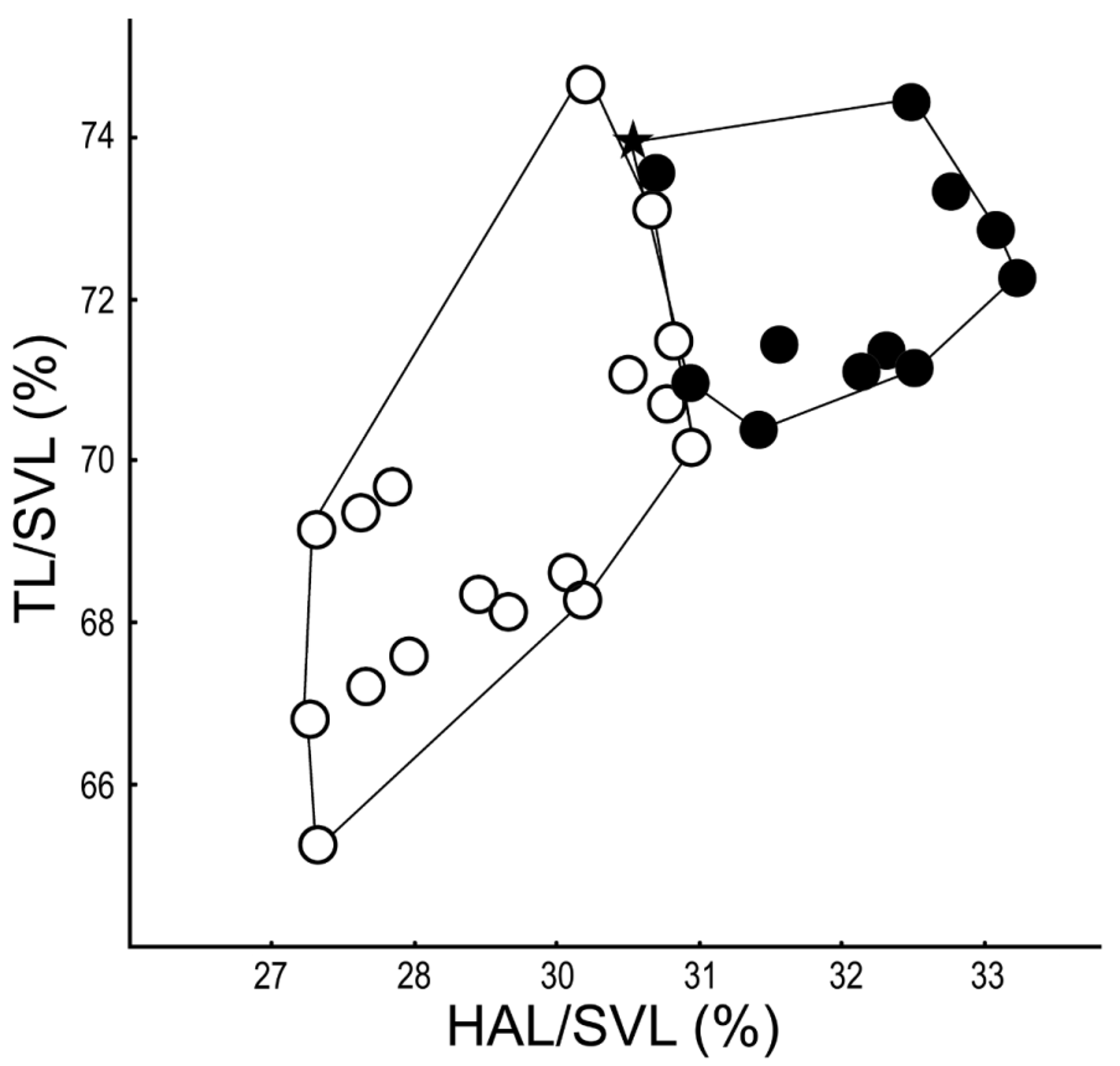
Meristogenys dyscritus resembles M. amoropalamus in adult morphology, but the typical individuals can be identified through morphological characters. The upper iris of M. dyscritus is usually sandy brown, but is yellowish green in M. amoropalamus ( Fig. 6 View FIGURE 6 ), although this character changes after anaesthesia, and there might be an intraspecific variation. Additionally, females of M. dyscritus have smaller ( Fig. 5 View FIGURE 5 ) ova and larger clutch size ( Fig. 4 View FIGURE 4 ) than M. amoropalamus (See “Reproductive characters” in Result). In females, M. dyscritus have longer hand (usually longer than 31% of SVL) than M. amoropalamus (usually less than 31% of SVL) ( Fig. 10 View FIGURE 10 ). Males of M. amoropalamus usually have larger tympanum (more than 13 mm 2) than those of M. dyscritus (less than 15 mm 2), although there is a range of overlap ( Fig 11 View FIGURE 11 ).
Contrary to the adult morphology, the larvae of M. dyscritus is quite different from those of M. amoropalamus in morphological characters. The larvae of M. dyscritus have six rows on upper jaws, while those of M. amoropalamus have seven. In the young developmental stages, the lower jaw sheaths of both larvae is divided, but only those of M. amoropalamus fuse into a large single jaw sheath before metamorphosis. Glands in a tail are limited to ventral fin in larval M. dyscritus , while they are present in both fins of larval M. amoropalamus . For the comparison between larval M. dyscritus and other Meristogenys larvae in Sabah, see the KEY TO MERISTOGENYS LAR- VAE FROM SABAH.
The larvae of M. dyscritus share divided lower jaw sheaths, six rows of labial teeth on the upper jaw, and no glands on their dorsal fin or ventral surface with larval M. orphnocnemis . The larvae of M. dyscritus , however, have rectangular body shape (body oval in M. orphnocnemis ), surface projections restricted to head (projections on head and body in advanced stage in M. orphnocnemis ), and usually seven or eight rows of lower labial teeth (usually six rows in M. orphnocnemis ; See Shimada et al., 2007a for details).
Range. Besides the type locality Liwagu, 1500 m a.s.l., adults of this species have been collected from Wario (900 m a.s.l.) in the Kinabalu National Park, and Mahua (1063 m a.s.l.) in the Crocker Range National Park, all in Sabah, Malaysia. The larvae of this species have also been collected from Bundu Tuhan (990 m a.s.l.) and Poring (500 m a.s.l.) in/around the Kinabalu National Park.
Natural history. Gravid females were collected in July (Liwagu), August (Mahua and Wario), November (Wario), and December (Mahua). Larvae collected in March (Liwagu and Mahua), August (Liwagu, Wario, Bundu Tuhan, and Mahua), and November (Poring) showed a wide range of developmental stages. Thus, there seems to be no particular reproductive seasons. Larvae were collected from rivers with a width of 5– 10 m. At night, they make a mixed cluster with larvae of M. kinabaluensis , M. orphnocnemis , and Huia cavitympanum on top of flat rocks in the river.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.