
Merostenus (Reikosiella) melinus (Yoshimoto) Yoshimoto, 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.556479 |
|
publication LSID |
lsid:zoobank.org:pub:3EF97DA3-EF37-4D91-8689-2AB9151D1440 |
|
DOI |
https://doi.org/10.5281/zenodo.6049216 |
|
persistent identifier |
https://treatment.plazi.org/id/03C187CB-225F-CF12-FF78-FC11FAF749E2 |
|
treatment provided by |
Plazi |
|
scientific name |
Merostenus (Reikosiella) melinus (Yoshimoto) |
| status |
comb. nov. |
Merostenus (Reikosiella) melinus (Yoshimoto) n. comb.
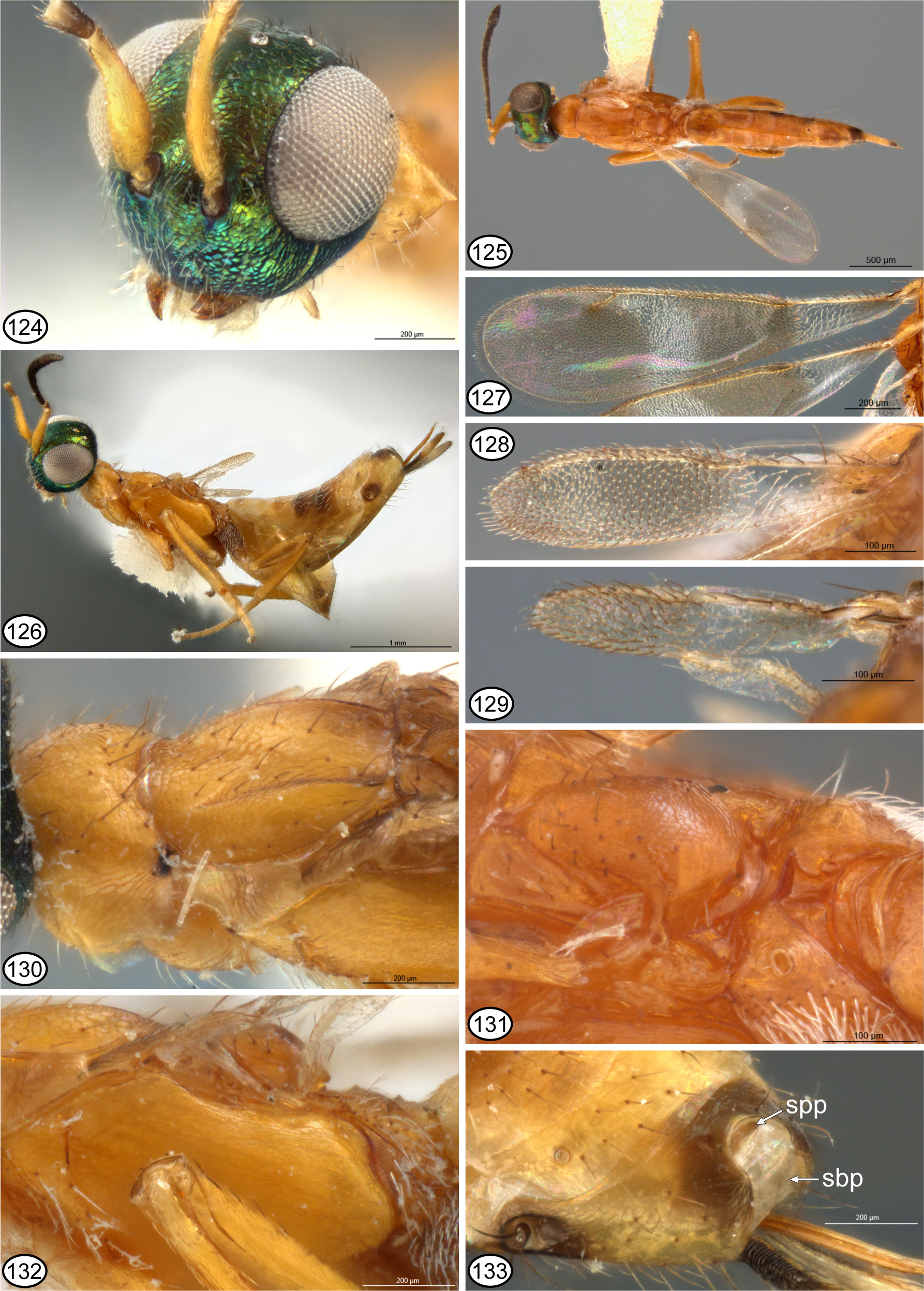
Figs 124–133 View FIGURES 124 – 133
Reikosiella melina Yoshimoto, 1969: 629 View in CoL . Holotype ( BPBM, macropterous ♀, Bishop 8021, examined). Type data: Mt. Kaala , Oahu, VI.1967, C. M. Yoshimoto.
Reikosiella melina View in CoL ; Bouček, 1988: 557 [species groups, biology]; Fusu, 2013: 32 [biology]. Reikosiella (Reikosiella) melina View in CoL ; Gibson, 1995: 265.
Holotype. The holotype is point-mounted by its right acropleuron and is entire except for its front legs, though it is contorted with the wings over the propodeum and gaster. The label data is as given in the original description except the collection date is stated as “ 20.VI. '67”.
Additional material examined (all BPBM unless stated otherwise). AUSTRALASIAN: Hawaii. Kauai, Kokee, VIII.1955 (1 ma PT ♀), 13.IX.1965 (1 br PT ♀), 16.IX.1965 (1 ma PT ♀, CNC Photo 2016-109), J.W. Beardsley. Lanai, Lanaihale , 920m, 25.III.1966, J.W. Beardsley (1 ma ♀) . Maui , Kula Agric. Park, I–VI.1988 (1 ma ♀, CNC Photo 2016-110), 28.VIII.1988, on ground with Argentine ants (1 ma ♀), J.W. Beardsley. Molokai, nr. Halawa Val., 7.IX.1956, J.W. Beardsley, reared ex. Plectronia fruit with Orneodes (5 PT ♀: 2 ma and 2 br; 1 br ♀ CNC Photo 2016-112); Kalaupapa Lookout, 24.III.1966, J.W. Beardsley (1 ma ♀) ; Mapulehu , X.1954, J.W. Beardsley (1 br PT ♀) . Oahu, Ewa , 1.III.1960 (1 br PT ♀), 2.III.1960 (1 br PT ♀), 4.VIII.1965 (1 br PT ♀), 23.IX.1965 (1 PT ♀), J.W. Beardsley; Ewa, Coral Plain, 27.X.1955, J.W. Beardsley (1 br PT ♀) ; Honolulu, P.H., 18.X.1950, K.S. Hagen (1 ma♀ UCRC 00407593); Honolulu, HSOA Exp. Sta., 20.VII.1955, J.B. Beardsley (1 ma PT♀); Kolekole Pass , 1725 ft, 27.II.1967, J.R. Vockeroth (1 ma ♀ CNC) ; Mt. Kaala , 17.VI.1964, H.L. Carson (1 ma ♀) ; Peacock Flats , 1800 ft, 18.VIII.1988, J. Straznanac & C.J. Pritzler (1 br ♀) ; Waianae Mts. , 14.VIII.1963, D. Perkins (2 br ♀), 14.VIII.1963 (1 br ♀), D. Perkins, ex. Lantana berries containing seed fly; Wailupe Val., 13.XII.1959, J.W. Beardsley (1 br ♀) ; Waipio, Peninsula , 24.XI.1965, J.B. Beardsley (1 br PT ♀, CNC Photo 2016- 111). NEOTROPICAL: Argentina. Missiones, Parque Nat. Iguazy, Cantera, 200 m, 8.XII.90 – 6.I.1991, S. & J. Peck (1 br♀ CNC, CNC Photo 2016-108). Brazil. Jatai , Goias, I.1977, F.M. Oliveira (1 ma ♀ and 1 br ♀ CNC).
Yoshimoto (1969) stated that in addition to BPBM paratypes were deposited in BMNH, CNC and USNM, but none were located in the latter three collections.
Description. FEMALE (habitus: Figs 125, 126 View FIGURES 124 – 133 ). Length = 1.9–3.3 mm. Head ( Figs 124–126 View FIGURES 124 – 133 ) bright green with limited coppery to reddish-violaceous lusters under some angles of light; in frontal view subquadrate, about 1.1–1.2× as wide as high, and in lateral view about 1.4–1.5× as high as long with frontal surface highly convex; frons sometimes with variably distinct though shallow mediolongitudinal furrow between scrobal depression and median ocellus, but frontovertex otherwise mostly meshlike coriaceous, scrobal depression more coriaceousimbricate to alutaceous-reticulate with smaller-sized cells, and lower face more coriaceous-granular to distinctly granular with convex cells ( Fig. 124 View FIGURES 124 – 133 ); with longer, more conspicuous white setae on lower face and interantennal prominence comparted to darker and much sparser setae on upper face excluding bare scrobal depression; scrobal depression inconspicuously, transversely Π-shaped, consisting primarily of lateral margin directed dorsally at level of lateral margin of torulus so as to differentiate distinct but short parascrobal region, and shallow scrobe above each torulus, with region above interantennal prominence smoothly merged into frons such that dorsal limit of scrobal depression indistinct but at least widely separated from anterior ocellus ( Fig. 124 View FIGURES 124 – 133 ); ratio of OOL: POL: LOL: MPOD = 1.2–1.4: 2.4–3.2: 1.6–1.9: 1.0. Antenna ( Fig. 125 View FIGURES 124 – 133 ) with scape yellow ( Fig. 126 View FIGURES 124 – 133 ), pedicel yellow to brown dorsally ( Fig. 124 View FIGURES 124 – 133 ), and flagellum dark brown; scape spindle-shaped, widest subbasally, with length about 4.8–5.0× as long as wide; pedicel about 2.0–2.3× as long as wide; fl1 slightly longer than wide, fl2–fl4 all distinctly longer than wide, and fl5–fl8 decreasing in length such that apical funicular about 1.3–1.4× as long as wide, and clava about 2.4–3.2× as long as wide ( Yoshimoto 1969, fig. 1h). Labiomaxillary complex yellow. Mandibles yellowish basally and reddish apically.
Mesosoma ( Figs 130–132 View FIGURES 124 – 133 ) yellowish to orange except pronotal collar dark posterolaterally anterior to spiracle ( Fig. 130 View FIGURES 124 – 133 ), mesoscutal lateral lobes sometimes variably extensively dark longitudinally along angulation ( Fig. 130 View FIGURES 124 – 133 ), and scutellar-axillar complex sometimes variably extensively yellowish-brown to light brown. Pronotum ( Figs 125, 130 View FIGURES 124 – 133 ) completely sclerotized; collar shiny, at most only obscurely meshlike coriaceous, and at least along margins with longer dark setae than on the head though often bare or mostly bare medially. Mesoscutum ( Fig. 130 View FIGURES 124 – 133 ) similarly sculptured as pronotum; with anterior margin convex, not reflexed behind pronotum, and with comparatively lowly convex lateral lobes relative to broad and shallowly concave median region, the lateral lobes not carinately margined except sometimes posteriorly near transscutal articulation; dorsally with scattered dark setae, including along inclined outer surface of lateral lobe. Scutellar axillar complex ( Fig. 131 View FIGURES 124 – 133 ) with scutellum uniformly convex and elongate-ovate with sides converging anteriorly from frenum to form acute angle only slightly separated from transscutal articulation, quite highly convex above and conspicuously larger than axillae, meshlike coriaceous to coriaceous-imbricate laterally, and with dark setae laterally anterior to frenum; axillae elongate-triangular, almost flat, meshlike coriaceous, with dark setae, and anterior to scutellum separated by only punctiform depression. Fore wing ( Figs 127–129 View FIGURES 124 – 133 ) polymorphic in length, color pattern and venation, but entirely setose except for costal cell dorsally. Macropterous female with fore wing extending to about apex of syntergum ( Fig. 125 View FIGURES 124 – 133 ), about 3.5–3.8× as long as wide with mv about 1.0–1.3× as long as wing width, and with complete venation including long stigmal vein ( Fig. 127 View FIGURES 124 – 133 ; Yoshimoto 1969, fig. 1a, b); basal cell basally very slightly infuscate with at least a few darker setae, but mostly hyaline with white setae extending to level of base of parastigma, and disc hyaline to very slightly infuscate with dark setae except for hyaline cross-band with white setae starting at about mid-length of marginal vein and extending to stigmal vein anteriorly, though both basal and apical margins of band often slightly curved ( Fig. 127 View FIGURES 124 – 133 ); cc: mv: stv: pmv = 3.3–5.0: 3.6–5.0: 1.0: 2.0–2.5. Brachypterous female with fore wing variably strongly shortened and narrowed but at most with punctiform stigmal vein and usually venation consisting only of submarginal vein and vein along leading margin extending to near wing apex ( Figs 128, 129 View FIGURES 124 – 133 ; Yoshimoto 1969, fig. 1c–e); usually with more or less similar infuscation and setal color patterns as for macropterous female ( Fig. 128 View FIGURES 124 – 133 ), but when wing most strongly reduced sometimes with entirely dark setae or with only basal hyaline region with white setae ( Fig. 129 View FIGURES 124 – 133 ). Mesopectus with white setae ventrally and anterior to acropleural sulcus dorsolaterally; acropleuron ( Fig. 132 View FIGURES 124 – 133 ) posteriorly angulate with posterodorsal margin sinuate anterior to metapleural flange and posteroventral angulation, and only very finely meshlike coriaceous. Metanotum with dorsellum transverse, horizontal ( Fig. 131 View FIGURES 124 – 133 ) under protruding scutellar apex ( Figs 131, 132 View FIGURES 124 – 133 ). Metapleuron ( Fig. 132 View FIGURES 124 – 133 ) with about ventral half setose and dorsal half bare, smooth and shiny, and over about middle third sinuate anterior margin appressed to depression along posterodorsal margin of acropleuron, with ventral margin only slightly reflexed over base of metacoxa, and with linear region between acropleuron and base of metacoxa bare. Legs ( Fig. 126 View FIGURES 124 – 133 ) same color as mesosoma except outer surface of metacoxa variably dark brown and outer surface of metafemur sometimes brownish longitudinally; mesotibia with stronger, spine-like setae apically but not pegs; metacoxa with outer surface entirely setose ( Fig. 126 View FIGURES 124 – 133 ). Propodeum ( Fig. 131 View FIGURES 124 – 133 ) with transverse, carinate ridge medially within about anterior third to half differentiating anterior, medially concave portion under scutellar apex from lunate, concave to inclined posterior surface having variably long median carina and usually longitudinal to oblique carina on either side of median carina; callus with spiracle about in middle of convex region, separated from lateral margin by distance similar to own maximum diameter ( Fig. 131 View FIGURES 124 – 133 ), with transverse line of setae anterior to spiracle, laterally, and over almost entire surface posterior of spiracle to posterolateral margin of propodeal foramen and posterior margin of callus.
Metasoma with petiole transverse, linear ( Fig. 131 View FIGURES 124 – 133 ). Gaster ( Figs 125, 126 View FIGURES 124 – 133 ) similarly yellowish to orange as mesosoma except one or more of Gt3–Gt5 with darker brown regions at least laterally and syntergum similarly dark brown dorsally and anterolaterally in subcircular region encompassing cercus; very finely meshlike coriaceous with pale setae on Gt1–Gt3 and laterally on Gt4–Gt5, but dark setae dorsally on Gt4–Gt6 and syntergum; syntergum ( Fig. 133 View FIGURES 124 – 133 ) deeply emarginate and dorsally bare except for a couple of long setae along emargination laterally, and with brown supra-anal and subanal plates evident in uncollapsed specimens; ovipositor sheaths ( Figs 125, 126 View FIGURES 124 – 133 ) rigid, projecting beyond syntergum by distance equal to about half length of metatibia; banded with thinner basal portion dark ( Fig. 126 View FIGURES 124 – 133 ) though region sometimes concealed under syntergum ( Fig. 125 View FIGURES 124 – 133 ), and thicker apical portion variably extensively pale basally and variably dark brown apically ( Figs 125, 126 View FIGURES 124 – 133 ).
MALE. Unknown.
Distribution. AUSTRALASIAN: Hawaii. NEOTROPICAL: Argentina, Brazil.
Hosts. Uncertain, but reared from fruits of Psydrax odorata (Förster) A. C. Sm. & S. P. Darwin (Rubiaceae) , purportedly from the larvae or pupae of Alucita objurgatella (Walsingham) ( Lepidoptera : Alucitidae ) ( Yoshimoto 1969). Label data also indicates specimens were reared from “ Lantana berries” ( Verbenaceae ) containing a “seed fly”, and one female is labelled as found on the ground with Argentine ants, Linepithema humile (Mayr) (Formicidae) .
Although Yoshimoto (1969, p. 631) “surmised” that M. melinus was a larval or pupal parasitoid of A. objurgatella , it has been reared only once from this host and then only 3 females were mass reared from fruits of P. odorata . Reared from the same fruits were 135 specimens of the purported host as well as four other parasitoid species, including 8 specimens of Euderus sp. ( Chalcidoidea : Eulophidae ), 20 specimens of Eupelmus sp., 9 specimens of Fidiobia sp. (Platygastroidea: Scelionidae ) and 3 specimens of Pristomerus sp. (Ichneumonoidea: Ichneumonidae ), plus mites living within cavities from which moths had emerged. It was also noted that many of the fruits contained one or two entrance holes that probably were made by the moth larvae chewing their way in after hatching from eggs laid externally on the fruit. Species of Fidiobia are all egg parasitoids and, according to L. Masner (AAFC, pers. comm.), of eggs of weevils ( Coleoptera : Curculionidae ). The known biology of M. excavatus as a curculionid egg predator that subsequently pupates within the plant associate raises the possibility that this might also be so for M. melinus . Even if not, rearing of the other parasitoid species raises the possibility that it might be a hyperparasitoid through one of the other primary parasitoids. Another rearing from Lantana berries, purportedly from some seed fly, further suggests a broader host range than just A. objurgatella . The association with Argentine ants is definitely unusual, but might simply be an accidental association.
Remarks. Other than M. melinus , species of M. ( Reikosiella ) are restricted to North, Central, and South America. It is therefore anomalous that the species was described from Hawaii, as the only known species of the genus from the Hawaiian Islands. However, even though A. objurgatella is known only from Hawaii, Zimmerman (1901) stated that it is not native and someday its true native range would be discovered elsewhere. The discovery of a brachypterous female of M. melinus from Argentina and a macropterous and brachypterous female from the same collecting event in Brazil suggests that R. melinus was introduced to Hawaii from South America. Of the Hawaiian eupelmid fauna, only Eupelmus is speciose with over 60 described species ( Noyes 2016). This suggests a relatively ancient introduction of Eupelmus into Hawaii and subsequent explosive diversification within the islands. The presence of M. melinus as the only species of Merostenus in Hawaii indicates a much more recent introduction. The CNC also has macropterous females of other species of M. ( Reikosiella ) from Central and South America that are quite similar in body color pattern to M. melinus , which suggests closely related species. However, females of M. melinus uniquely have the metapleuron partly setose ( Fig. 132 View FIGURES 124 – 133 ) in combination with rigid ovipositor sheaths that are only about half the length of the metatibia and have a dark-pale-dark colour pattern ( Fig. 126 View FIGURES 124 – 133 ), though the basal dark region sometimes is hidden under the syntergum ( Fig. 125 View FIGURES 124 – 133 ). Further, other macropterous females seen from the Neotropical region with similar body colour patterns have completely hyaline fore wings with dark setae.
Yoshimoto (1969) recognized macropterous females, four intergradations of brachypterous females, and micropterous females. I did not see any females he described and illustrated as micropterous, i.e. with the wing having only a ‘rudimentary’ vein, apparently restricted to the basal half of the wing ( Yoshimoto 1969, fig. 1f). Females listed above under material examined as macropterous have fore wings with complete venation including a distinct stigmal vein angled away from the postmarginal vein ( Fig. 127 View FIGURES 124 – 133 ) (= macropterous sensu Yoshimoto 1969 , fig. 1a, and brachypterous intergradation 1 sensu Yoshimoto 1969 , fig. 1b). Females with variably strongly shortened fore wings ( Figs 128, 129 View FIGURES 124 – 133 ) that either lack a distinct stigmal vein ( Yoshimoto 1969, figs 1c–e) or have a punctiform to short stigmal vein that is appressed to the postmarginal vein are considered as brachypterous.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Merostenus (Reikosiella) melinus (Yoshimoto)
| Gibson, Gary A. P. 2017 |
Reikosiella melina
| Fusu 2013: 32 |
| Gibson 1995: 265 |
| Boucek 1988: 557 |
Reikosiella melina
| Yoshimoto 1969: 629 |