
Mortoniella (Mortoniella) spatulata, Blahnik & Holzenthal, 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.5170203 |
|
publication LSID |
lsid:zoobank.org:pub:AB1A57F0-7CB4-4830-920B-DF219740A596 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687A7-FFCD-F830-FF01-BB0641D7FC0F |
|
treatment provided by |
Felipe |
|
scientific name |
Mortoniella (Mortoniella) spatulata |
| status |
sp. nov. |
Mortoniella (Mortoniella) spatulata , new species
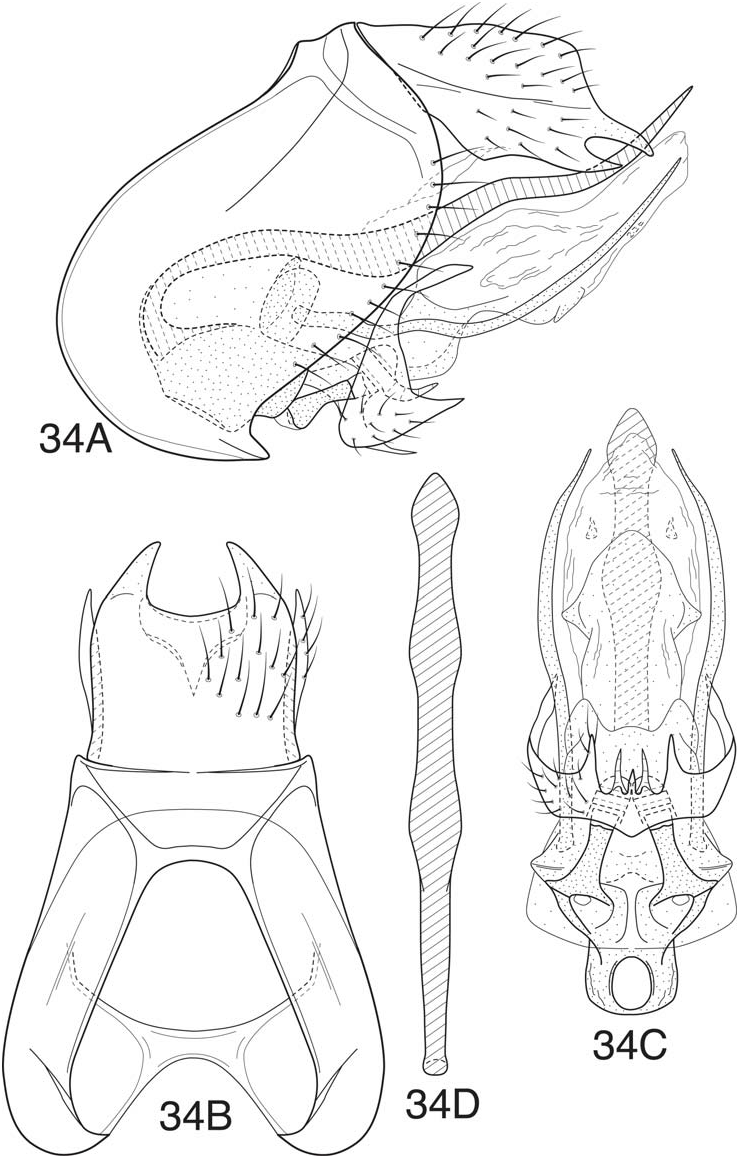
Fig. 34 View Figure 34
Mortoniella spatulata bears a general similarity to the other species of the bolivica subgroup, but the dorsolateral lobes of the inferior appendages are shorter and not as strongly flexed. Also, the dorsal phallic spine, while being widened and spatulate apically, is more nearly symmetrical, the ventrolateral lobes of tergum X are acutely developed (separated from the apicolateral lobes by a distinct notch), and the ventral spines of the endophallic membrane are very weakly developed. The holotype specimen is over cleared and only the genital capsule (segments IX-X and phallic apparatus) is present.
Adult —Length of forewing: male 3.8 mm. Forewing with forks I, II, and III present, hind wing with forks II and III. Spur formula 0:4:4. Overall color light brown, apices of tarsi and basal segments of antennae slightly paler. Tibial spurs about same color as legs, not contrasting in color. Wing bar at anastamosis relatively indistinct, interrupted, marked with whitish setae.
Male genitalia —Segment IX nearly evenly rounded anterolaterally, length greatest midlaterally, posterolateral margin convexly rounded dorsally, narrowing ventrally; segment deeply mesally excised dorsally and ventrally, forming lateral lobes, separated dorsomesally by much less than ½ width of segment. Tergum X with excision between lateral lobes slightly concave; apicolateral lobes acute, curved inward from lateral margins, mesal margins subparallel. Inferior appendages only moderately large, setose, incompletely divided ventromesally, apicolateral angles short and acute, dorsolateral lobes narrow and slightly posteriorly recurved, apices acute. Mesal pockets of inferior appendage with apical processes relatively short, dorsally curved. Paramere appendages elongate, narrow, extending about as far as dorsal phallic spine. Dorsal phallic spine, as viewed laterally, distinctly undulate in contour, nearly uniform in width, narrowing apically, base distinctly curved, apex very weakly and obtusely upturned in about apical ¼; in dorsal view, with apex slightly widened and spatulate in shape (dorsoventrally flattened). Phallicata with dorsal margin weakly sclerotized, apparently to accommodate dorsal phallic spine. Endophallic membrane moderately elongate and relatively simple, ventrally with pair of very short, weakly sclerotized spines; phallotremal spines (apparently) present, very small.
Holotype male (pinned)— VENEZUELA: Barinas: Parque Nacional Sierra Nevada, Quebrada San Juan in Santa Rosa , 8.46450° N, 70.84867° W, 1000 m, 21.iii.1997, Holzenthal ( UMSP000041337 View Materials ) ( UMSP). GoogleMaps
Etymology —This species is named M. spatulata for the dorsal phallic spine of the male, which has its apex flattened and distinctly spatulate in shape.
— florica/leroda subgroups included species (South America): Mortoniella bothrops , n. sp.; M. cressae , n. sp.; M. curtispina , n. sp.; M. draconis , n. sp.; M. elongata (Flint) ; M. furcula , n. sp.; M. grandiloba n. sp., M. ruedae , n. sp.; M. schlingeri , n. sp.; and M. simla (Flint) .
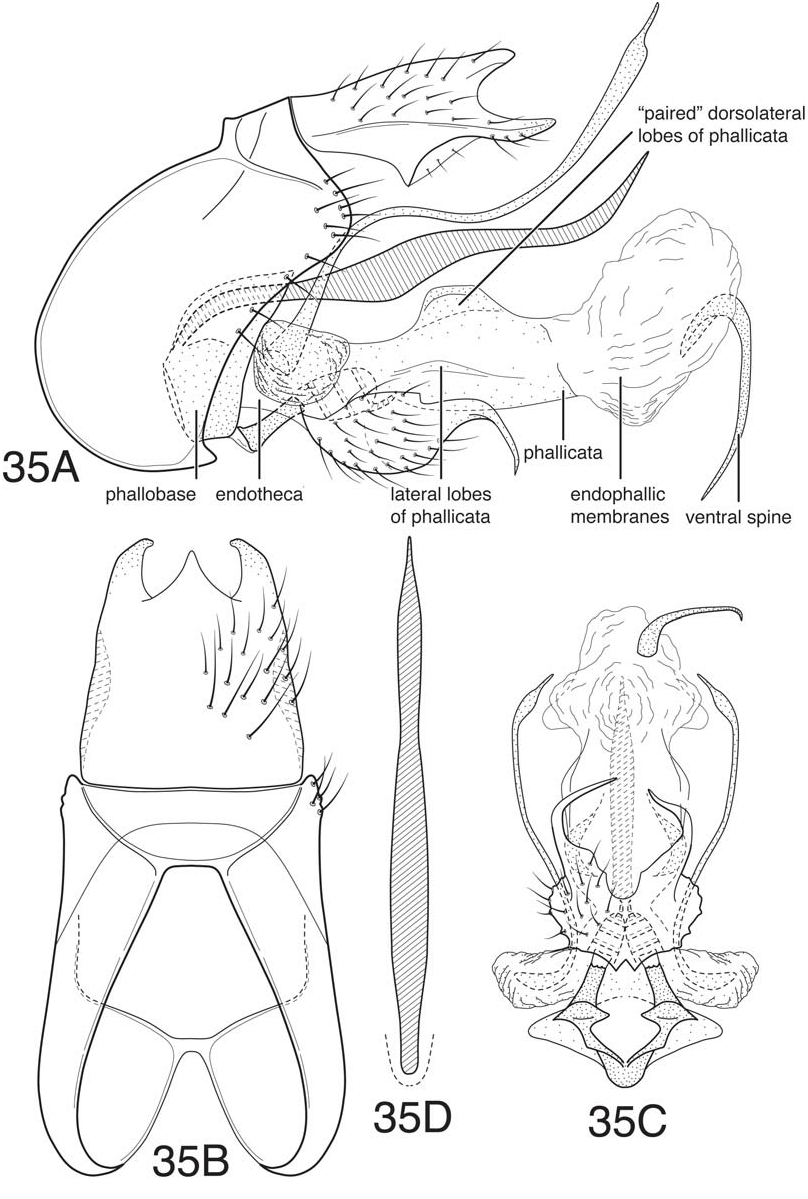
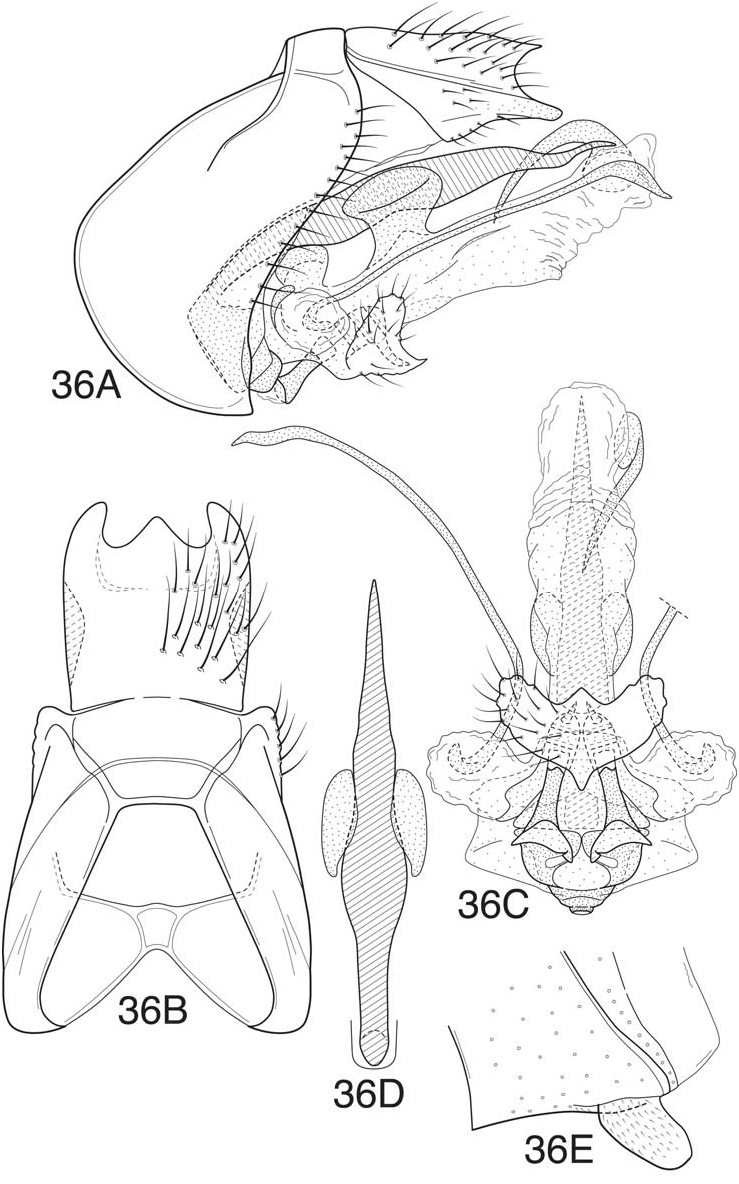
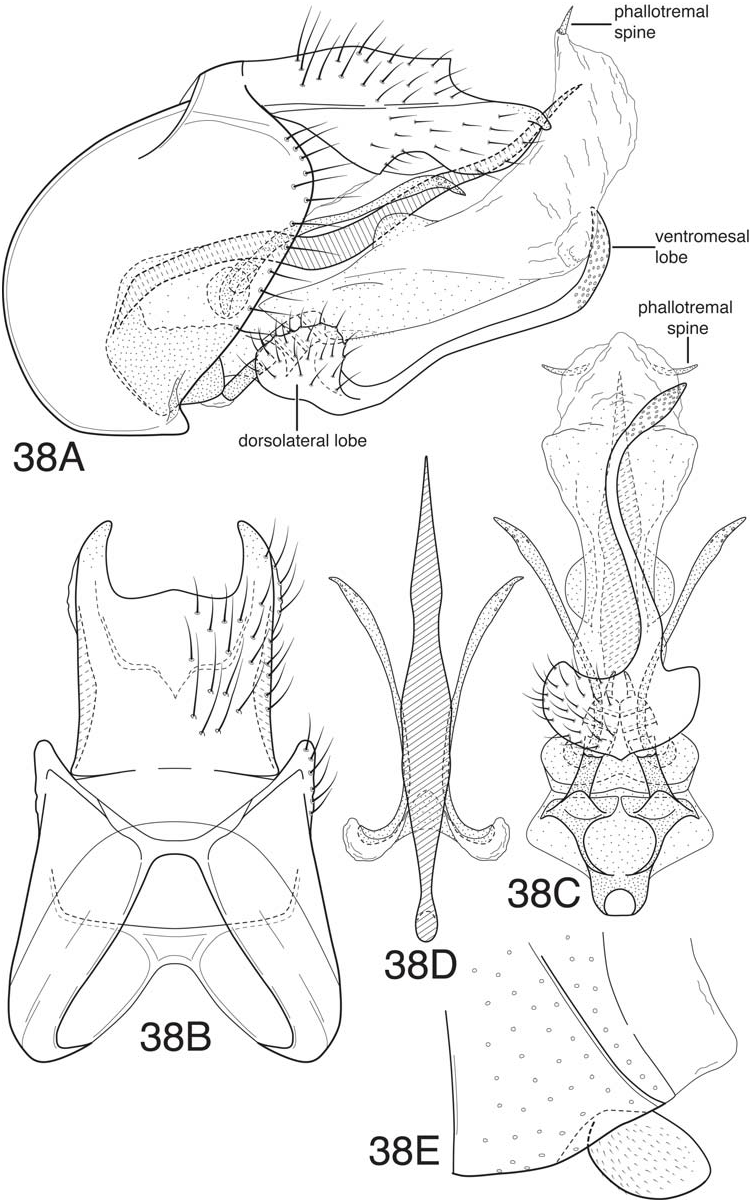
Blahnik and Holzenthal (2008) defined the leroda and florica subgroups based on species from Central America and commented that the two groups were probably related. As in the majority of the subgroups of the leroda group treated in the current work, the hind wings in both subgroups have both forks II and III present. Also, the dorsolateral projections of the inferior appendages are either absent or greatly reduced in both subgroups, usually appearing as short rounded projections. The latter is considered an apomorphy for the combined subgroups, since a projecting dorsal lobe is probably the plesiomorphic state for the subgenus Mortoniella . Many of the species, especially of the leroda subgroup, have a rather characteristically formed ventral process on segment VI, which is short, ventrally projecting, and with the apex more or less blunt or bluntly rounded. The anterobasal margin of the ventral process is characteristically somewhat retracted ( Fig. 38E View Figure 38 ). However, many species in the florica subgroup from Central America have the ventral process more angular apically, and not as evidently retracted anteriorly, though always distinctly ventrally deflected. A bluntly rounded ventral process otherwise occurs only in the atenuata subgroup, which is apomorphically distinct, and also in M. guyanensis , n. sp., in the limona subgroup. An additional, although more difficult to define, character uniting the two subgroups is the general structure of the dorsal phallic spine, which has a characteristic undulate or sinuous inflection, probably due to the tendency for the ventral margin of the spine to have a rounded deflection that articulates with a corresponding contour in the dorsal margin of the phallicata. However, this may be a plesiomorphic character for the leroda group as a whole, since a similar character state also occurs in the akantha and bolivica subgroups. Another character, which seems to characterize most of the species of the two subgroups, is that the paramere appendages are displaced ventrally on elongate membranous lobes, with the base of the appendages curved inward. This is already evident in M. leroda , which seems to be a basal, or near basal, species in the combined subgroups. The character is more developed in species with paired dorsolateral processes on the phallicata, especially those of the florica subgroup (eg. Fig. 35C View Figure 35 , 36C View Figure 36 ). An additional character similarity of the two subgroups is the structure tergum X, which has apicolateral lobes that are distinctly sclerotized and projecting and have a very reduced or nearly absent ventrolateral lobes. There is also some tendency for the lobes to be flattened and converge mesally, and for the dorsomesal margin to be projecting.
Although the majority of species from Central America seemed to fall into 2 groups, based on a combination of characters, in treating the species from South America the distinctness of the two groups is less evident. It therefore seems prudent to combine them here. The primary character difference used to define the two groups was the structure of the inferior appendages; members of florica subgroup have paired symmetrical apical processes on the inferior appendages (long or short), and those of the leroda subgroup have a single asymmetrical apicomesal process. Because the names were used previously, and the primary defining character is diagnostically useful, we have continued to group the species from South America in the two subgroups, while recognizing that they may not be mutually monophyletic. Many of the species of the leroda subgroup from South America have upright dorsal processes on the phallicata, a ventromesal spine on the endophallic membrane, and scabrous apices of the paramere appendages (characters generally found in the florica subgroup of Central America, but absent in the species assigned to the leroda subgroup). These characters occur in addition to the presence of a prominent asymmetric ventromesal process on the inferior appendages, used to characterize the leroda subgroup. Most of the South American species of the leroda subgroup also have the apex of this mesal process scabrous (or sensillate), as in M. meralda (Mosely) and M. panamensis Blahnik and Holzenthal from Central America. It seems likely that an asymmetric ventromesal process on the inferior appendages is a plesiomorphic character that has been subsequently lost in the florica subgroup. This may also explain the presence (retention) of this character in a number of species of the albolineata subgroup from Brazil, and an indication of the relationship of these subgroups. The most likely explanation for the presence of paired rounded or upright basodorsal processes on the phallicata in the majority of species of the florica subgroup is that they represent a lineage from within the leroda subgroup that derived this apomorphy, followed by the subsequent loss of the ventromesal process and gain of apicolateral projections on the inferior appendages. The new species described below as M. cressae ( Fig. 39) possibly represents a transitional species (or one that has converged on similar character modifications), with apicolateral projections on the inferior appendages present, but with the ventromesal process much reduced. Absence of a ventromesal spine on the endophallic membrane in various species of the leroda subgroup, especially those from Central America, probably represents a character loss, judging by its general presence (retention) in the florica subgroup and in a number of other subgroups. However, there seems to be no parsimonious solution for its loss, which must have occurred repeatedly. Possibly, the plesiomorphic state was for the spine to be very small, as in M. leroda , or in the species of the atenuata subgroup, making its loss more probable. If redefined in a combined sense to include both members of the leroda and florica subgroups, the composite subgroup would include all of the species from Central America, in addition to the species described below, with the exception of the three species of the akantha subgroup and M. pacuara (Flint) and M. rodmani Blahnik and Holzenthal of the subgenus Nanotrichia . This entire group constitutes a revised definition of the leroda subgroup, and will be referred to subsequently as the leroda subgroup, sensu lato.
— “florica” subgroup
| UMSP |
University of Minnesota Insect Collection |
| VI |
Mykotektet, National Veterinary Institute |
| A |
Harvard University - Arnold Arboretum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.