
Krypyonesticus deelemanae, Pavlek & Ribera, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.262 |
|
publication LSID |
lsid:zoobank.org:pub:10676882-F6AC-4BC8-A0F1-9C0882EA36C2 |
|
DOI |
https://doi.org/10.5281/zenodo.3844379 |
|
persistent identifier |
https://treatment.plazi.org/id/222787B2-FFEB-FF8D-FD94-86B64038FCF1 |
|
treatment provided by |
Carolina |
|
scientific name |
Krypyonesticus deelemanae |
| status |
gen. et sp. nov. |
Krypyonesticus deelemanae gen. et sp. nov.
urn:lsid:zoobank.org:act:BAC5D31D-7A59-4FE7-BE9A-CB6167484170
Figs 1 View Fig 1 , 2A View Fig 2 , 3 View Fig 3 A–C, 4A, D, 5A, D
Diagnosis
The most prominent character of the male bulb is a large and conspicuous median apophysis which emerges from the middle part of the tegulum and reaches its apical part. The most obvious differences between the new species and the closest ones, K. fagei and K. arenstorffi , are: distal part of MA in the new species is massive and slightly thinner toward the top, while in K. arenstorffi the whole distal part is slender and the top is more narrowed; the proximal part of MA of the new species has a round process on the outer edge, while in K. fagei and K. arenstorffi it has no process. Another important feature is the tooth-like dorsomedian apophysis of the paracymbium’s dorsal process, which is not present in K. fagei and K. arenstorffi . The inner side of the distal part of MA, the curve between the proximal and distal parts of MA, the shape of the tegulum, the basal tegular apophysis and the dorsal paracymbium process are also diagnostic. The epigyne is most similar to that in K. fagei and K. arenstorffi , from which it differs in the outline of the posterior epigyne margin which is triangular in K. fagei and sinuous in other two species. The difference between the new species and K. arenstorffi is in the posterior epigyne border, lateral of the central chitinous arches, which is straight in the new species and slightly notched in K. arenstorffi . The vulva can be distinguished by the position of the spermathecae, whose apical part in the new species reaches over the vulvar chitinous structures, while it does not in K. fagei and K. arenstorffi .
Etymology
The specific name is a patronym in honor of Christa Laetitia Deeleman-Reinhold, an important Dutch arachnologist and a dear friend. Her work has vastly raised the knowledge of the cave spider fauna of Dinarides. The species name is in possessive genitive.
Material examined
Holotype CROATIA: 1 ♂, Biokovo Mt , Samogorska špilja, 43°19′5.71″ N, 17°7′29.9″ E, elevation 717 m, 23 Jan. 2016, leg. Marko Lukić ( CBSS / AR 3934-1 ). GoogleMaps
Paratypes
CROATIA: 1 ♀ ( CBSS /AR 3934-2), 1 ♂, 6 ♀♀ ( CBSS /AR 3934-5), leg. Marko Lukić; 1♂, 1♀, leg. Marko Lukić ( SMF); 1♂, 7 ♀♀, leg. Tin Rožman ( CBSS /AR 3932). All with same collection data as the holotype.
Other material examined
CROATIA: 1 ♂, Biokovo Mt, Kuna jama, 27 Aug. 1998, leg. Roman Ozimec ( CBSS /AR 1495); 1 ♂, Biokovo Mt, Crna Ledenica, 24 Oct. 2006, leg. Marko Lukić ( CBSS /AR 1277); 1 ♂, Biokovo Mt, Stonjska peć donja, 27 Oct. 2003, leg. Roman Ozimec ( CBSS /AR 1287).
Comparative material
TURKEY: K. dimensis ( López-Pancorbo, Kunt & Ribera, 2013) , 1 ♂, 1 ♀, Antalya Province, Alanya District, Kestel Town, Dim Valley, Dim Cave, 3 Dec. 2011, leg. Kadir Boğaç Kunt.
CROATIA: K. eremita (Simon, 1879) , 2 ♀♀, 2 ♂♂, 2 juv.. ( CBSS /AR 1160), Dubrovnik, Osojnik, Močiljska špilja, 21 June 2001, leg. Roman Ozimec. K. arenstorffi ( Kulczyński, 1914) and K. fagei ( Kratochvíl, 1933) : see below.
Description
Male (holotype)
COLORATION. Carapace uniform whitish, opisthosoma yellowish without marked darker patches. Sternum of the same color as carapace. Appendages slightly darker, especially the distal segments ( Fig. 1A View Fig 1 ).
PROSOMA. Approximately circular in dorsal view. Cephalic region not differentiated from rest of prosoma. Fovea and thoracic grooves visible. Eyes normally developed. Eye size and interocular distances: AM =
0.042; AL = 0.081; PM = 0.085; PL = 0.077; AM–AL = 0.07; AM–AM = 0.067; PM–PL = 0.075; PM– PM = 0.129; PL–AL = 0.02.
OPISTHOSOMA. Sub-elliptical in dorsal view.
CHELICERA. Mesal part of promargin with three teeth of about the same size.
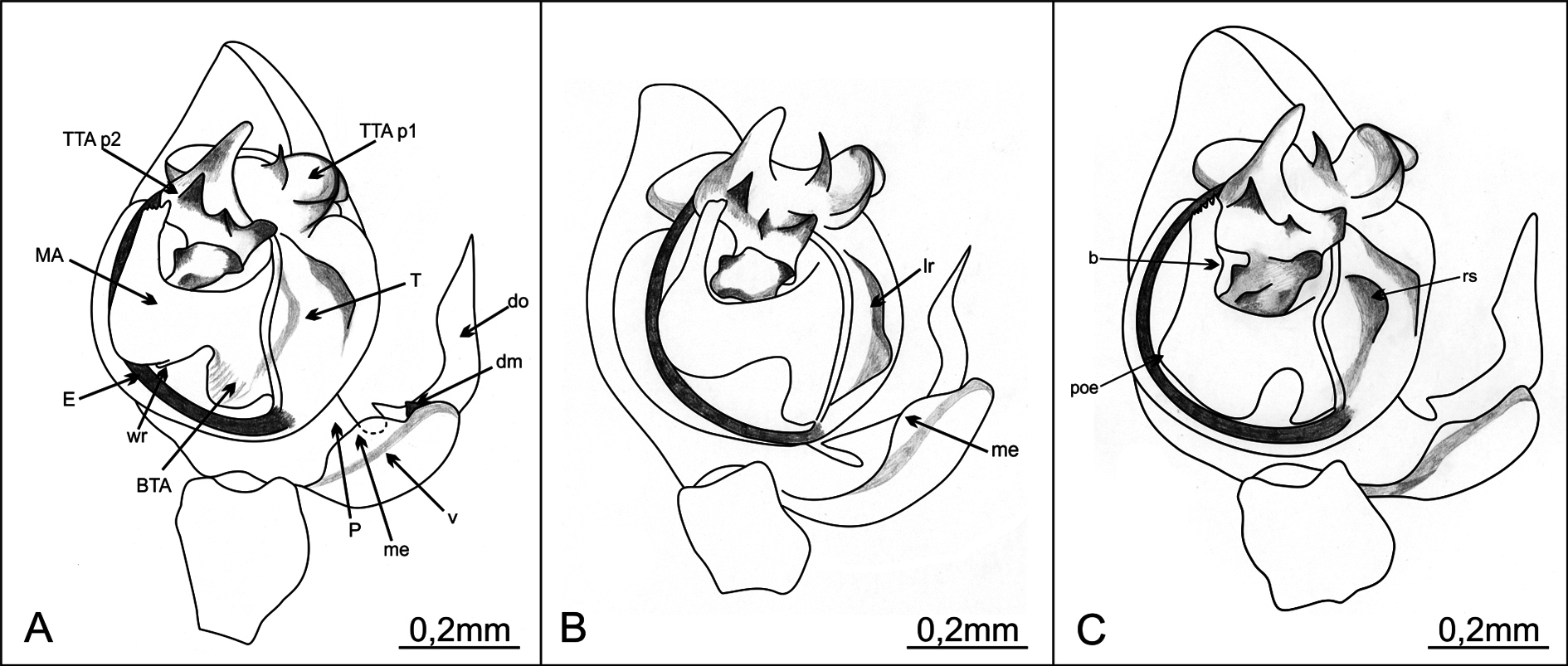
MALE PALP. Paracymbium with a well-developed dorsal and very simple ventral processes. Dorsal process with two branches, apical branch parallel to cymbium’s longer axis and shorter basal branch pointing to cymbium. Apical branch with parallel rims in proximal part, slightly dilated in middle with distal part acuminated, dagger-shaped. Basal branch with convex upper rim and with distal part sharp and tilted upward ( Figs 2A View Fig 2 , 3B View Fig 3 ). Dorsomedian apophysis tooth-like, small with broad base. Ventral process ( Figs 2A View Fig 2 , 3A View Fig 3 ) broad, rounded, with transparent middle part and a membranous extension on rim ( Figs 2A View Fig 2 , 3A View Fig 3 ). Tegulum with a shallow, rounded and broad longitudinal groove (visible in apical view, Fig. 3C View Fig 3 ) and with a broad, roughly circular basal apophysis on base (in ventral view). Median apophysis remarkably well developed, V-shaped, distal arm reaching almost to apical part of tegulum. Proximal part of MA with wrinkled outer rim which in posterior view looks like a small process. Outer edge between proximal and distal part equally rounded, distal part of MA mildly narrowing toward top, bluntly ended, with slightly serrated upper edge. Inner side of distal part of MA slightly swollen. Conductor absent. TTA with two processes: TTA p1 and TTA p2 (homologous to processes p1–p6 of the conductor complex in Huber 1993). TTA p1 saddle-shaped, longer than wide, with slim, rounded and pale process on top and in central area with three small dark teeth in a row, middle one smallest and proximal one faintly rounded. TTA p2 located in an apical position, with one sharp and pointed process, the rest forming groove where tip of embolus lodges and which serves as a conductor. Filamentous embolus partially borders tegulum and reaches its apex in semicircular course ( Figs 2A View Fig 2 , 3A View Fig 3 ). Measurements: PL: 1.552; PW: 1.407; OL: 2.075; OW: 1.412; TL = 3.627. Leg formula: I> IV> II> III.
Female (paratype CBSS/AR 3934-2)
SOMATIC CHARACTERS ( Fig. 1B View Fig 1 ) As in male, except that eye size and interocular distances are a bit different and body length a little larger: AM = 0.052; AL = 0.095; PM = 0.085; PL = 0.076; AM–AL = 0.061;
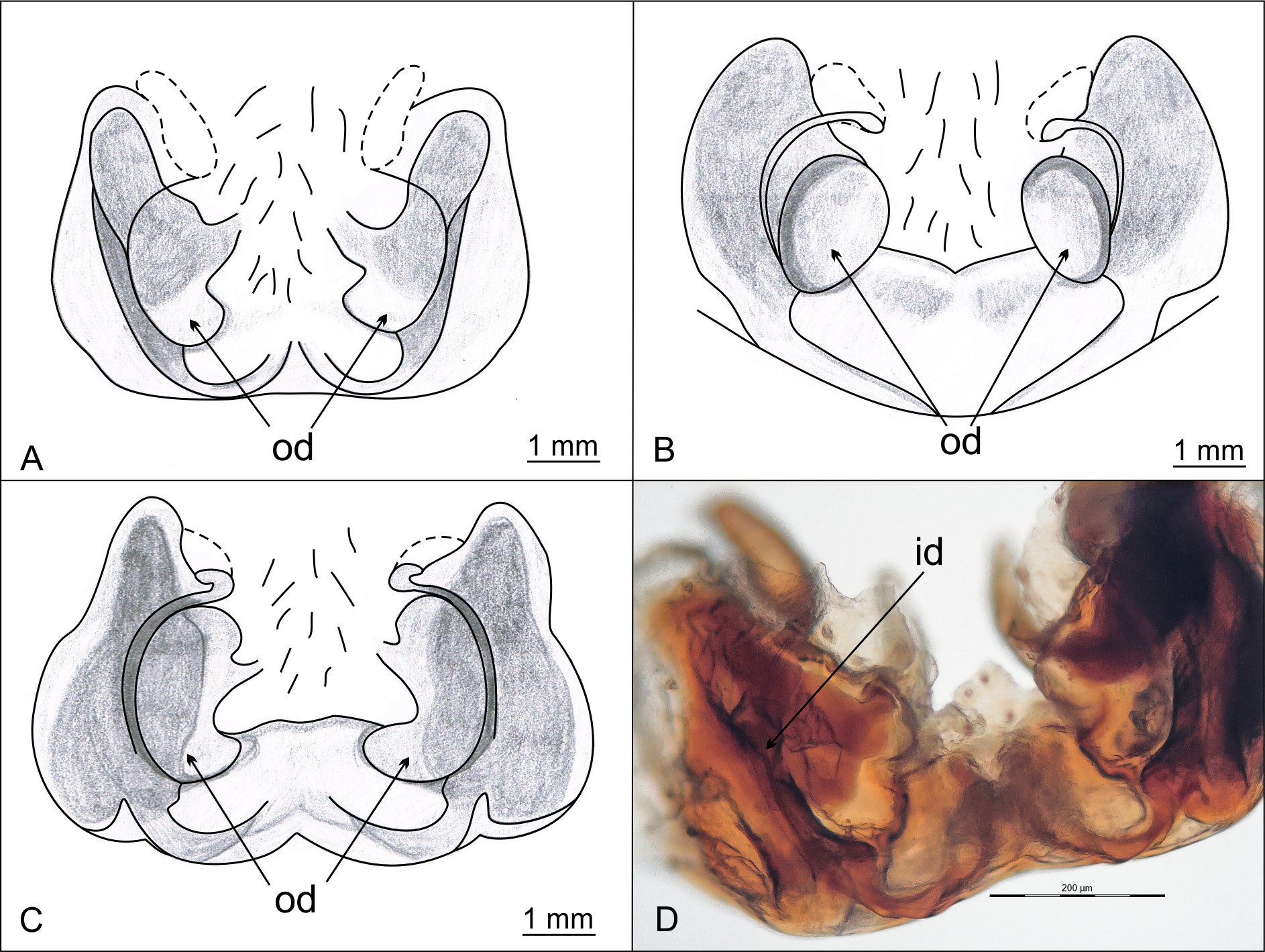
AM–AM = 0.070; PM–PL = 0.128; PM–PM = 0.1112; PL–AL = 0.015. Epigyne slightly wider than long, in lateral view not much protruding ( Fig. 5D View Fig 5 ). Lateral lobes with oval depressions in posterior part, reaching middle of epigyne. Posterior epigyne border made of two semicircular chitinous arches, flanking depressions, coming together in middle and forming a small, pale, inconspicuous convex tubercule. A ridge apically bordering tubercle and depressions faintly visible. Epigyne outline, laterally from semicircular chitinous arches, straight, with no notches ( Figs 4A View Fig 4 , 5 View Fig 5 A–B).
VULVA. Lateral sides of vulva made of compact dark chitinous structures enclosed in a firm transparent membrane (lateral pouches). Oval shaped spermathecae, with a shallow constriction in middle, located just above chitinous vulval structures (dorsal view). Narrower, upper part of spermathecae protrudes above chitinous vulval structures ( Fig. 5C View Fig 5 ). Insemination ducts depart from spermathecae at bottom ( Fig. 5C View Fig 5 ), make a sharp turn outwards and continue ventro-laterally in semicircular course to reach copulatory orifices ( Fig. 4D View Fig 4 ). In ventral view, spermathecae slightly visible above lateral dark structures, while chitinous curves, through which insemination ducts run visible on both sides of epigyne ( Fig. 4A View Fig 4 ). Two chitinous narrow, horn-like apophyses protrude from inner sides of dark chitinuos vulval structures, in apical direction, to empty middle part of vulva ( Fig. 5C View Fig 5 ). Measurements: PL: 1,666; PW: 1,414; OL: 2,609; OW: 1,872; TL = 4,275. Leg formula: I> IV> II> III.
Distribution
The new species is endemic to Croatia; it is distributed on Biokovo Mt in central Dalmatia, a coastal region in Croatia. So far it has been recorded in 20 caves scattered through the whole mountain, from the south-west sea side to the north-east continental side, from the 310 to 1640 asl ( Fig. 6A View Fig 6 ). Data on all records of K. deelemanae gen. et sp. nov. are given in Appendix 2 View Appendix 2 . The distribution area of K. deelemanae gen. et sp. nov. is more than 80 km away from that of K. fagei and more than 100 km from that of K. arenstorffi ( Fig. 6B View Fig 6 ).
Natural history
The type locality, Samogorska špilja, is a small cave with two entrances ( Fig. 7 View Fig 7 ). On the date of the last collection, 23 Jan. 2016, the air temperature in the cave was 0.5°C with cold air streaming through the cave, mostly near the cave floor. Spiders were found freely walking on the ceiling of the chamber (probably avoiding the cold air flow at the bottom) and on the side walls of the cave. The temperature in other caves where K. deelemanae gen. et sp. nov. is found ranges from 0 to 15°C. Some of those caves are very small and are greatly influenced by outside conditions (like the type locality), while the others are quite big, have a true cave microclimate and harbor diverse types of cave habitats (for example Pretnerova jama, a 254-meter deep pit). No other nesticid species are found in caves on Biokovo Mt.






| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |