
Gromphas aeruginosa (Perty, 1830)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3722.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:20D125E7-8CC0-4942-8AF9-75EA31EBBD53 |
|
DOI |
https://doi.org/10.5281/zenodo.5668589 |
|
persistent identifier |
https://treatment.plazi.org/id/707787C6-5973-624B-FF0A-250036379D89 |
|
treatment provided by |
Plazi |
|
scientific name |
Gromphas aeruginosa (Perty, 1830) |
| status |
|
1. Gromphas aeruginosa (Perty, 1830) View in CoL
Figs. 1–6, 16, 22, 24, 29, 34, 36–37, 39, 41, 46, 48, 52–53, 57, 62–63, 66
Onitis aeruginosus Perty, 1830: 39 ; Lacordaire 1856: 105; Burmeister 1874: 130; Scherer 1983: 298; Cupello 2013: 15–17.
Gromphas aeruginosa: Harold 1859: 199 ; Harold 1869a: 1016, 1869b: 62; Waterhouse 1891: 60; Heyne & Taschenberg 1908: 64, plate 9 Fig. 42 View FIGURES 34 – 48. 34 ; Ohaus 1909: 135; Gillet 1911: 80; d’Olsoufieff 1924: 20, 58, 138, plate II Fig. 3, plate IX; Pessoa & Lane 1941: 478; Blackwelder 1944: 208; Barattini & Saenz 1960: 23, 1964: 177; Halffter & Matthews 1966: 131; Vulcano & Pereira 1967: 566; Halffter & Edmonds 1982: 86; Scherer 1983: 298; Kirk 1992: 49; Amézquita et al. 1999: 119; Escobar 2000: 206; Medina & Lopera-Toro 2000: 303 Fig. 3c, 305 Fig. 4c; Vaz-de-Mello 2000: 193; Medina et al. 2001: 138; Noriega 2002: 72, 2004: 40; Philips et al. 2004b: 59 Fig. 33 View FIGURES 29 – 33 , 62 View FIGURES 57 – 62 ; Quinteros et al. 2006: 78; Monaghan et al. 2007: 681; Hamel-Leigue et al. 2009: 49, 2013: 281; Medina & Pulido 2009: 61; González-Hernández & Navarrete-Heredia 2011: 479; Figueroa et al. 2012: 1; Korasaki et al. 2012: 427; Cupello 2013: 15.
Gromphas lacordairei Blanchard, 1846: 181 ; Harold 1859: 199; Harold, 1869a: 1016, 1869b: 62; Gillet, 1911: 80; d’Olsoufieff, 1924: 138; Blackwelder, 1944: 208; Barattini & Saenz 1960: 23, 1964: 177; Hamel-Leigue et al. 2009: 61.
Type specimen:— Onitis aeruginosus Perty, 1830 : Lectotype: designated by Scherer (1983), undetermined sex, (“ Brasilien ”, “3-8 typi”, “ Type von Gromphas aeruginosa Perty ”, “alte Sammlung”, “ Holotypus Onitis aeruginosus Perty. det. Dr. G. Scherer, 1981”, “ Gromphas aeruginosus (Perty) det. G. Scherer, 1981”), ZSMC (examined by photos [ Figs. 6 View FIGURE 6 a–e]). Paralectotypes: 7 specimens, ZSMC (not examined).— Gromphas lacordairei Blanchard, 1846 : Not located at MNHN; unknown to us.
Type locality: — Onitis aeruginosus : “Habitat in mediterraneis Prov. S. Pauli et Minarum” (Perty 1830; but see comments below).— Gromphas lacordairei Blanchard : “Province de Santa-Cruz de la Sierra ( Bolivia)” (Blanchard 1846).
Redescription: Color: Anterior region of head black, remainder of dorsum with bright and metallic reflections of green, yellow, copper, and burgundy; some of these colors predominating or, more commonly, a mixture of all. Pygidium colored as elytra. Metasternum bright and of uniform color.
Head: Clypeus with two lobes in the apex (in worn specimens lobes absent and apex round or truncate); margin between these lobes and genae round ( Fig. 24 View FIGURES 24 – 28 ). Margin of genae extended beyond clypeal margin ( Fig. 24 View FIGURES 24 – 28 ). Clypeus with smooth tegument adjacent to eyes; granulation gradually denser and squamous toward outer margin ( Fig. 24 View FIGURES 24 – 28 ). Cephalic projection ranging from a low and transverse elevation arched upward to a horn flattened antero-posteriorly with apex distinctly bifid (apex truncate or rounded in worn specimens); horn granulate on both faces ( Figs. 29 View FIGURES 29 – 33 a–e). Frons smooth adjacent to eyes and granulate in center ( Fig. 24 View FIGURES 24 – 28 ).
Thorax: Lateral region of pronotum with dense squamous granulation (Fig. 16); granulation gradually sparser toward posteromedian region (Figs. 1–5). Longitudinal midline with smooth and glossy tegument present at center of pronotum (clearly in Fig. 3). Posterior pronotal fossae always present; granulation absent in fossae. Posterior margin of pronotum rounded.
Anterior region of pronotum with ornamentation ranging from two shallow impressions in anterior margin (Fig. 5) to prominence with two tapered projections apically obtuse or slightly acute; usually, projections largely divergent and directed towards sides of body; lateral margin of prominence curved (Fig. 1) or, less usually, straight (Fig. 2); separation between projections always very wide; anterior to prominence, a pair of concavities with smooth tegument (Fig. 16).
Mesosternum with dense pilosity ( Fig. 52 View FIGURES 49 – 52 ). Metasternum with fine and sparse punctation in the center, almost imperceptible in worn specimens. Anteromedian angle of metasternum flat and with rounded apex ( Fig. 52 View FIGURES 49 – 52 ), area in front of angle with evident setae (Figs. 16, 52).
Legs: Protibiae very broad in both sexes; in ventral view, longitudinal carina simple in both sexes ( Fig. 22 View FIGURES 21 – 23 ). Protibial spur with apex slightly bent laterally and downward ( Figs. 22 View FIGURES 21 – 23 , 46 View FIGURES 34 – 48. 34 ); inner apical protibial angle with a tuft of long setae in both sexes ( Fig. 22 View FIGURES 21 – 23 ). Apical protarsomere tapered and only slightly elongate at apex ( Figs. 22 View FIGURES 21 – 23 , 46 View FIGURES 34 – 48. 34 ). Metatibiae very broad and robust ( Fig. 41 View FIGURES 34 – 48. 34 ). Mesotarsi and metatarsi not particularly enlarged and with apical tarsomeres slightly curved apically ( Fig. 48 View FIGURES 34 – 48. 34 ). Metatibial spur with apex distinctly curved ( Fig. 39 View FIGURES 34 – 48. 34 ).
Elytra: Striae very fine and, especially those more medial, carinulate from base to no more than midlength of of elytra, where they become simple ( Fig. 37 View FIGURES 34 – 48. 34 ); interstriae flat. Sutural margin only sparsely punctate (Figs. 1–5).
Abdomen: Pygidium flat and not margined basally ( Fig. 36 View FIGURES 34 – 48. 34 ), sometimes with a pair of weak and almost imperceptible impressions on sides. Groove of propygidium extending to base of pygidium or penetrating to its basal third. Abdominal sternites sparsely punctate.
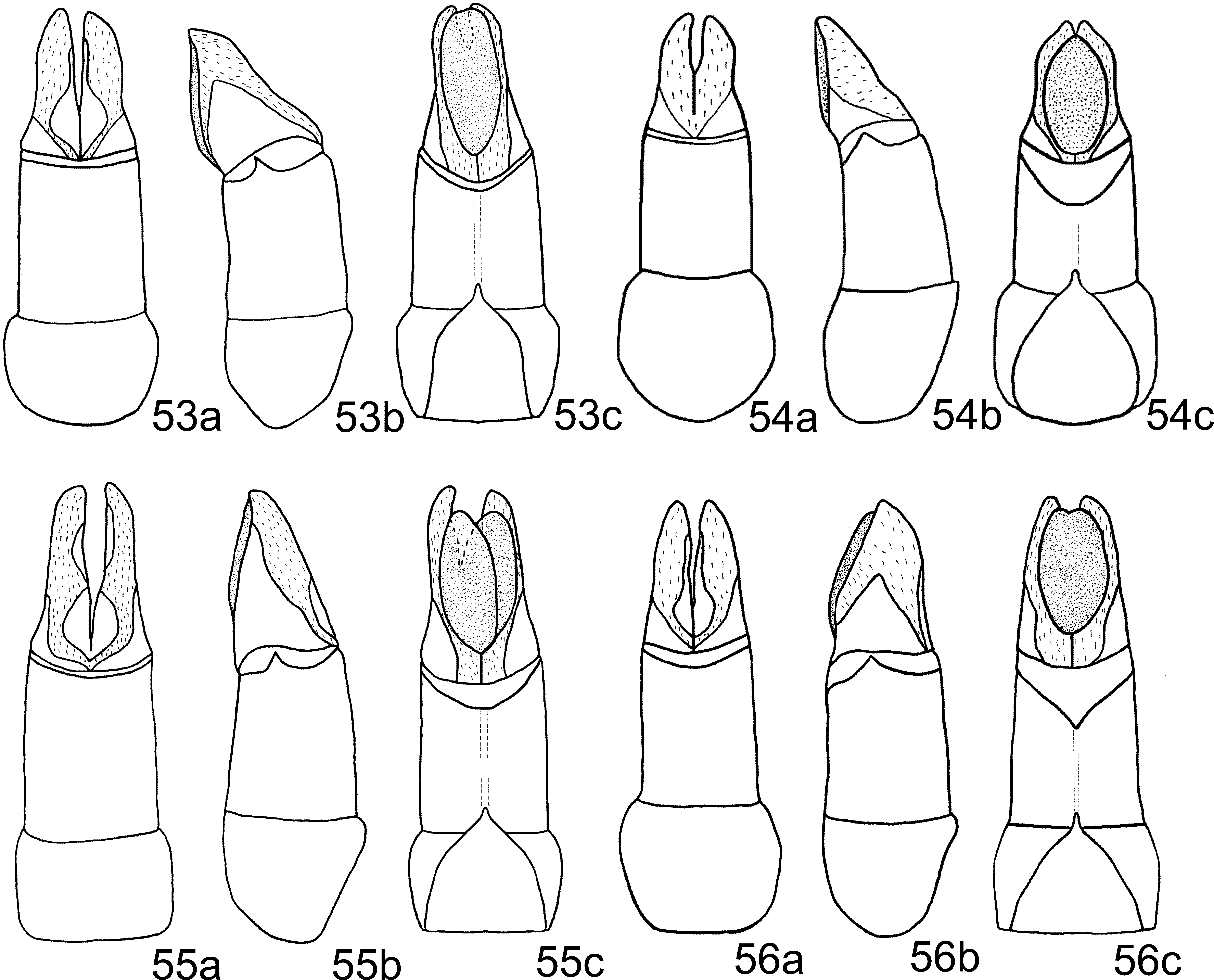
Aedeagus: Phallobase, in ventral view, with a very narrow apical membranous area ( Fig. 53 View FIGURES 53 – 56 c). Medial sclerite strongly curved downward ( Fig. 57 View FIGURES 57 – 62 ).
Measurements: Males (65 specimens): TL: AV: 15.44; MX: 20.3; MN: 11.6; SD: 1.87. PL: AV: 12.42; MX: 15.9; MN: 9.6; SD: 1.32. PW: AV: 9.13; MX: 11.7; MN: 6.8; SD: 1.17. PP: AV: 2.79; MX: 4.7; MN: 0.9; SD: 0.84. Females (50 specimens): TL: AV: 15.71; MX: 20.3; MN: 11.9; SD: 1.87. PL: AV: 12.59; MX: 15.3; MN: 9.7; SD: 1.36. PW: AV: 9.29; MX: 12.4; MN: 6.9; SD: 1.2. PP: AV: 2.89; MX: 5; M: 1; SD: 0.95. Total (115 specimens): TL: AV: 15.56; SD: 1.87. PL: AV: 12.49; SD: 1.34. PW: AV: 9.19; SD: 1.18. PP: AV: 2.83; SD: 0.89.
Intraspecific variation and taxonomic discussion: A wide variation in the degree of development of the ornamentation is observed in this species. Small specimens have the cephalic projection and especially the pronotal prominence weakly developed (in smaller specimens the pronotal prominence is completely absent and the two anterior concavities are reduced to a pair of shallow impressions [Fig. 5]), while large specimens have them well developed, with the width of pronotal prominence up to 5 mm (Figs. 1–2). Between these two extremes, an entire gradual variation is present, with most specimens in an intermediate position (Figs. 3–4).
Previous authors mistakenly believed that this difference in development was related to sexual dimorphism. Perty (1830) stated that males had an armed head and thorax, while females had the head transversely carinate and thorax unarmed; and Blanchard (1846) thought that the cephalic horns of females of G. lacordairei Blanchard would be brief and rudimentary. d’Olsoufieff (1924) was the first to recognize correctly that the females of G. aeruginosa could have the ornamentation as developed as males; even so, in some recent works, such as the cladistic analysis of Philips et al. (2004b), the ornamentation is still incorrectly regarded as sexually dimorphic. We observed that the average width of the pronotal prominence does not differ significantly between the sexes, and the same is true with the cephalic projection. Having already recognized this, Barattini & Saenz (1960, 1964) pointed out the presence of a wide allometry in this species. As demonstrated for a wide variety of other dung beetles, the ornamentation of G. aeruginosa is positively allometric in relation to body length and pronotal width, but lacks the dimorphic condition found in some other genera, such as Onthophagus and Oxysternon , and in other beetles (e.g., Eberhard & Gutiérrez 1991; Edmonds & Zidek 2004; Emlen 1997a, b; Kawano 2006).
Sexual dimorphism of G. aeruginosa thus appears to be restricted to that observed for the genus in general (i.e., protarsus present only in females and sixth visible abdominal sternite narrowed in the middle in males and of even width in females). Barattini & Saenz (1960, 1964) observed a sexual difference in the apex of clypeus; according to them, only the females have the clypeus upturned. It is true that many well-preserved specimens of both sexes have the margin of the clypeus (and of genae, to a lesser degree) slightly folded, but we did not observe any sexual difference.
In addition to the degree of development, the shape of the ornamentation also varies. Specimens of G. aeruginosa may possess the projections of pronotal prominence with apices acuminate or rounded. Most specimens have the projections widely divergent, but in a few specimens the projections are subparallel and the pronotal prominence is “U” shaped (Fig. 2). Still, all this variation occurs within a very limited range, and a cohesive unit is clearly visible. The species closest to G. aeruginosa , G. lemoinei , has a different shape of this prominence (see discussion of G. lemoinei ).
Unrelated to the development, the carinulae of the elytral striae also vary widely. In most specimens only the two most medial striae have evident carinulae, and they never extend beyond the apical half of the elytra ( Fig. 37 View FIGURES 34 – 48. 34 ). However, many specimens have the second stria entirely simple and the first very weakly carinulate, while in others the carinulae are evident even in the third and fourth striae.
Harold (1869b), after observing the type specimens of G. lacordairei Blanchard then deposited at the Jardin des Plantes (current MNHN), confirmed the synonymy between G. aeruginosa and G. lacordairei Blanchard that he proposed in 1859. Although we have not examined the type specimens of G. lacordairei Blanchard (see comments below), this seems to be a correct position, since the description of Blanchard is very clear and consistent with G. aeruginosa . Moreover, we examined more than 20 Bolivian specimens of G. aeruginosa and did not find any notable difference from the rest of the species. The other species present in Bolivia, G. inermis , clearly does not fit the description given by Blanchard (e.g., “ Le prothorax est trés-épais, élevé antérieurement et muni dans cette partie de deux pointes divergentes ”). The only other species that would be consistent with this description, G. lemoinei , does not occur in Bolivia or in the rest of the Amazon subregion. Also, the name Onitis aeruginosus Perty was at no time cited by Blanchard, suggesting that he was unaware of the description of Perty’s species.
Comments: The type series of G. aeruginosa was collected during the expedition of Johann Baptist von Spix and Karl Friedrich Philip von Martius through Brazil (1817–1820), who, from Rio de Janeiro, toured the southeast, northeast and north of that country (Papavero 1971). Despite being cited as the type locality by Perty, the former provinces and now states of São Paulo and Minas Gerais in southeastern Brazil are certainly not part of the distribution of this strictly Amazonian species. None of paralectotypes or the lectotype of G. aeruginosa possess labels indicating something more precise than Brazil ( Fig. 6 View FIGURE 6 e). Although the material collected in southeastern Brazil and in the Amazon were sent to Bavaria at different times (Papavero 1971), it is possible that they have been mixed up, which would have led to Perty’s mistake (also revealing, Phanaeus chalcomelas , another Amazon species collected by Spix and Martius and originally described in Onitis by Perty (1830), also has the wrong type locality of São Paulo and Minas Gerais).
Many type specimens described by Perty were lost during World War I and II, but most of the type specimens of Scarabaeidae survived, including eight syntypes of G. aeruginosa (Scherer 1983) . According to Scherer (1983), the original labels written in pencil by Perty were removed from these specimens and replaced by other labels by subsequent curators. Designated by Scherer (1983), the lectotype of G. aeruginosa ( Fig. 6 View FIGURE 6 ) has seven labels, none written in pencil ( Fig. 6 View FIGURE 6 e). Based on information indicated by him, the orange label written “ type von Gromphas aeruginosa Perty ” was prepared by Hans Kulzer, former preparator of the Zoologische Staatssammlung München; and the circular green label indicates that the material came from South America. The red-border label written by Scherer states “ holotypus ”, although the specimen has been designated by Scherer himself (Scherer 1983) as a “lectoholotypus”, equivalent to lectotype (Evenhuis 2008). It is clear that Perty (1830) based his description on more than one specimen because he made comparisons between what he considered to be males and females.
We examined only photographs of the lectotype, and therefore could not to confirm its sex. Scherer said it was a male, but it is likely he believed that the differences in the development of ornamentation were sexually dimorphic. The confusion in sex determination probably occurred also with the paralectotypes, of which, according to Scherer, there were three males and four females.
We initially doubted the commonly cited date of publication of Onitis aeruginosus , but 1830 seems to be correct. Blackwelder (1957) and Scherer (1983) cited this year as the publication of pages 1–60 of the “ Insecta Brasiliensis ” part of “ Delectus Animalium Articulatorum ”; Onitis aeruginosus was described on page 39. Also published in 1830, the eighth plate has the figure “8. Onitis aeruginosus ”, the first published illustration of a specimen of Gromphas known to us. (The date of publication of G. lacordairei Blanchard is also discussed in the comments on G. dichroa ).
The type of G. lacordairei Blanchard was not found by us. Based on what Blanchard (1846) wrote, we assume that is a series of syntypes. This assumption is based on the length provided for G. lacordairei Blanchard ; he did not mention an exact measurement, but a range (“ long. 15 à 30 millim ”).
Two junior primary homonyms of Onitis aeruginosus Perty were proposed in the 19th century: O. aeruginosus Gistel, 1831 and O. aeruginosus Klug, 1855 . The first name was proposed for a Brazilian species, but has not been cited (as valid or invalid) by any other author since; as its description is too vague and no type specimens are known to exist, Cupello (2013) regarded it as a nomen dubium and asked the International Commission on Zoological Nomenclature (ICZN) to suppress the name O. aeruginosus Gistel for the purposes of the Principle of Priority and of the Principle of Homonymy. The Klug’s name, however, is consistently and largely applied to an Afrotropical species of Onitis , and it is not considered as congeneric of Perty’s species since the latter was transferred to Gromphas by Harold (1859). Therefore, in order to preserve stability, Cupello (2013) asked the ICZN to rule that the name O. aeruginosus Klug is not invalid by reason to being a junior primary homonym of O. aeruginosus Perty , and thus remains as the valid name of this African dung beetle species.
The reported occurrences of G. aeruginosa in Paraguay (Heyne & Taschemberg 1908) and in the Guianas (Vulcano & Pereira 1967) are probably incorrect. The presence in the Colombian departments of Meta and Casanare, as pointed out by Medina & Pulido (2009), is also unlikely; it is possible that the specimens studied by them are misidentified G. lemoinei (in fact, we examined several specimens of G. lemoinei collected in Meta). The specimens from the MZSP collection recorded for Santa Catarina, Brazil, certainly are mislabeled. These specimens were found in a drawer among specimens of G. inermis that had the same labels, correct for that species.
Bionomics: Gromphas aeruginosa seems to be a common species in artificial open environments and in natural Amazonian savannas (Amezquita et al. 1999, Kirk 1992, Ohaus 1909). Specimens are found in pastures feeding on cattle and equine dung, and are also collected in traps baited with human feces (Hamel-Leigue et al. 2009).
Regarding the activity time, Kirk (1992) and Hamel-Leigue et al. (2009) observed that G. aeruginosa is diurnal. However, specimens collected in Cruzeiro do Sul (Acre, Brazil) by R. L. Gonçalves have labels stating that they have been caught in a light trap. This species is also common at lights in towns of Acre and Rondonia in Brazil (FZVM, personal observation). The altitudinal range recorded for this species is 190–500 m in Bolivia (Hamel- Leigue et al. 2009) and 200–350 m in Colombia (Amezquita et al. 1999, Noriega 2002).
Based on the specimen labels examined in this present work, we can say that the adults of G. aeruginosa are collected throughout the year.
Geographic distribution: Amazonian subregion: Napo, Imeri, Varzea, Ucayali, Madeira, Tapajós-Xingú, Pantanal and Yungas. Chacoan subregion: Chaco. COLOMBIA: Huila: Gigante. Amazonas: Letícia. BRAZIL: Amazonas: Benjamin Constant, Borba, Eirunepé, Fonte Boa, Guajará, Itacoatiara, Manacapuru, Manaus, Parintins, São Gabriel da Cachoeira, São Paulo de Olivença, Tabatinga, Tefé. Pará: Altamira, Juruti, Óbidos, Santarém. Acre: Bujari, Cruzeiro do Sul, Feijó, Rio Branco. Rondônia: Porto Velho. ECUADOR: Napo. Orellana: Francisco Orellana (Chiruisla Station). PERU: Loreto: Maynas (Iquitos). San Martin: San Martin (El Porvernir). Ucayali: Coronel Portillo (Iparía; Callería: Pucallpa; Yarinacocha. Huánuco: Leoncio Prado (Tingo María). Madre de Dios: Tahuamanu (Iñapari); Tambopata (Puerto Maldonado: Lago Sandoval). Cuzco: Quispicanchi (Camanti: Quince Mil). BOLIVIA: Beni: (El Trapiche: Estación Biológica del Beni). Santa Cruz: Agustín Saavedra, Andrés Ibáñez (El Torno: La Angostura; Palmasola; Potrerillos del Guenda), Cordillera, Florida (Pampa Grande: Mataral), Ichilo (Buena Vista), José Miguel de Velasco (San Miguel de Velasco), Ñuflo de Chávez (Los Troncos), Obispo Santistevan (Montero; Saavedra). Cochabamba: Chapare. ( Fig. 66 View FIGURE 66 ).
Material examined: 320 males and 271 females (381 with undetermined sex). BOLlVIA: without date and collector— 1 male (BMNH). COCHABAMBA: Chapare, without date and collector— 1 male (MZSP); Chapare, without date, Zischka col.— 1 female (MZSP); Chapare, 400 m., without date, Zischka col.— 1 male (CMNC). SANTA CRUZ: Flórida, Pampa Grande, Mataral, 08.XII.1994, M. Quiroz col.— 1 male (CMNC); Cordillera, Rio Parapetí, I.1964, Martínez col.— 1 male (CMNC); José Miguel de Velasco, San Miguel de Velasco, 26.XII.2009, Vidaurre et al. cols.— 1 male (CEMT); Obispo Santisteva, Cuatro Ojos, without date and collector— 1 male and 1 female (MZSP); Obispo Santisteva, Montero, Rio Piraí, 23.III.1987, F. D. B. col.— 1 female (CMNC); Obispo Santisteva, Rio Piraí, 08.I.1994, S. Graz col.— 1 male and 1 female (CMNC); Obispo Santisteva, Saavedra, CIMCA (Centro Integral de Manejo del Comportamiento Animal), 10.IX.1987, C. J. Pruett col.— 1 male (CMNC); Obispo Santisteva, Saavedra, CIMCA, 09.IX.1988, C. J. Pruett col.— 1 male (CMNC); Gutierrez, Nueva Moka, III.1956, A. Martínez col.— 1 female (MZSP); Ichilo, Buena Vista, 700 pés (518 m.), X.1922, J. Steinbach co.l— 1 male (BMNH); Ichilo, Buena Vista, I.1950, A. Martínez col.— 1 male and 1 female (MZSP); Ichilo, Buena Vista, II.1950, without collector— 3 males (MLPA); Sara, Nueva Moka, I.1963, A. Martínez col.— 1 male and 1 female (MZSP). BRAZIL: ACRE: Bujari, 20.VI.1995, L. M. de Andrade col.— 1 female (CEMT); Bujari, 09.VII.1995, L. M. de Andrade col.— 1 female (CEMT); Cruzeiro do Sul, VI.1952, J. Correa col.— 2 males and 6 females (MNRJ); Cruzeiro do Sul, IX.1952, J. Correia col.— 2 males and 5 females (MNRJ) and 2 males and 2 females (MZSP); Cruzeiro do Sul, IX.1956, R. L. Gonçalves col.— 4 males and 5 females (MNRJ); Cruzeiro do Sul, Fazenda Morada Nova, 07º37’57”S / 72º43’43”W, 25-26.XI.1996, C. S. Motta et al. col.— 4 males (INPA); Feijó, XI.1956, Dirings col.— 22 males and 23 females; Feijó, XII.1956, Werner col.— 1 male and 2 females (MZSP); Iquiri, VIII.1951, without collector— 1 male (MZSP); Rio Branco, 30.V.1988, Craveiro col.— 1 male (CEMT); Rio Branco, 11.VIII.1994, M. L. Guerreiro col.— 1 female (CEMT); Rio Branco, 19.VIII.1994, N. Batista col.— 1 male (CEMT); Rio Branco, 14.IV.1995, M. F. S. de Oliveira & A. M. de Almeida— 1 female (CEMT); Rio Branco, 10.VI.1995, S. M. N. Galdino col.— 1 female (CEMT); Rio Branco, 15.VI.1995, S. M. F. Almeida col.— 1 male and 1 female (CEMT); Rio Branco, Campus Universitário, 15.III.1982, William col.—1 with undetermined sex (DZRJ). AMAZONAS: Benjamin Constant, III-IV.1942, A. Parko col.— 3 males and 1 female (MNRJ); Benjamin Constant, XI.1960, collector illegible— 1 female (MNRJ); Benjamin Constant, XII.1960, L. G. Pereira col.— 2 males and 2 females (MNRJ); Benjamin Constant, 0 9.III.2004, P. H. Silva col.— 1 female (CEMT); Benjamin Constant, 21º25’45’’S 44º36’21’’W, IX.2008, V. Korasaki col.— 1 male (CEMT); Benjamin Constant, Rio Javari, IV.1942, B. Pohl col.— 2 males and 2 females; Benjamin Constant, Rio Javari, IV-VI.1942, A. Parko col.— 1 male and 1 female (MZSP); Benjamin Constant, Rio Javari, VII.1942, A. Parko col.— 1 female (MNRJ); Benjamin Constant, Rio Javari, VIII.1960, Dirings col.— 2 males and 2 females; Benjamin Constant, Rio Javari, I.1961, Dirings col.— 3 males and 1 female (MZSP); Benjamin Constant, Rio Javari, III.1961, Dirings col.— 1 male (MZSP); Benjamin Constant, Rio Javari, I-VIII.1961, Dirings col.—136 with undetermined sex (MZSP); Benjamin Constant, Rio Javari, V.1961, Dirings col.— 1 female (MZSP); Benjamin Benjamin Constant, Rio Javari, VIII.1961, Dirings col.— 13 males and 5 females (MZSP); Benjamin Constant, Rio Javari, XII.1961, Dirings col.— 4 males and 5 females (MZSP); Benjamin Constant, Rio Javari, X.1962, Dirings col.— 5 males and 6 females (MZSP); Benjamin Constant, Rio Javari, XI.1962, Dirings col.— 1 female (MZSP); Benjamin Constant, Rio Javari, IX-XI.1962, Dirings col.—210 with undetermined sex (MZSP); Benjamin Constant, Rio Javari, IV.1963, Dirings col.— 3 females (MZSP); Benjamin Constant, Rio Javari, X.1963, Dirings col.— 3 females (MZSP); Benjamin Constant, Rio Javari, Alto Amazonas, without date, Dirings col.— 2 male and 1 female (MZSP); Borba, III.1943, A. Parko col.— 4 males and 3 females (MNRJ) and 1 male (CEMT); Borba, Mata do Borba, II.1943, A. Parko col.— 2 males (MZSP) and 2 females (MNRJ); Codajás, 10.X.1935, without collector— 2 males and 1 female (MNRJ); Eirunepé (João Pessoa), Rio Juruá, 10.IX.1936, Zellibor-Hauff col.— 1 female (MNRJ) and 1 male and 2 females (MZSP); Fonte Boa, X.1975, F. M. Oliveira col.— 3 males and 2 females (MNRJ); Guajará, VII.1943, A. Parko col.— 1 male (MZSP); Lago Amanã, XI.1983, R. C. Best col.— 1 male and 1 female (INPA); Itacoatiara, XI.1961, Dirings col.— 1 female (MZSP); Manacapuru, Costa do Pesqueiro II, 03º23’35”S / 60º36’57”W, 16.IX.2006, F. F. Xavier col.— 1 male (INPA); Manaus, without date and collector— 3 males and 2 females (MZSP); Manaus, VII.1927, J. F. Zikán col.— 1 female (MNRJ); Manaus, VIII.1927, J. F. Zikán col.— 2 males (MZSP); Manaus, 1936, A. Ducke col.— 1 female (MNRJ); Manaus, INPA/Aleixo, 06.IV.1990, J. A. O. Guerra col.— 2 males (INPA); Manaus, Rio Negro, VII (without year), J. F. Zikán col.— 1 male (FIOC); Manaus, Rio Negro, VIII.1927, J. F. Zikán col.— 1 male and 1 female (MNRJ); Parintins, 1921, Garbe col.— 1 males (MZSP); São Gabriel da Cachoeira, Rio Negro, without date and collector— 1 female (MNRJ); São Paulo de Olivença, Rio Solimões, without date, Dirings col.— 1 male (MZSP); Tabatinga, XII.1956, Pereira col.— 2 males and 1 female (MNRJ); Tefé, without date, Dirings col.— 9 males and 12 females (MZSP); Tefé, without date, J. F. Zikán col.— 1 female (CEMT); Tefé, VIII (without year), J. F. Zikán,— 1 male (MZSP); Tefé, VIII.1921, J. F. Zikán col.— 2 males (FIOC) and 1 male and 1 female (MNRJ); Tefé, IX.1921, without collector— 1 male (MZSP); Tefé, X.1958, Dirings col.—14 with undetermined sex (MZSP); Tefé, VIII.1960, Dirings col.—15 with undetermined sex (MZSP); Tefé, XII.1961, Dirings col.— 3 males and 1 female (MZSP); Tefé, III.1962, Dirings col.— 8 males and 10 female (MZSP); Tefé, VIII.1991, O. Roppa & P. Magno cols.— 128 males and 87 females (MNRJ). PARÁ: Altamira, Rio Xingu, without date, J. F. Zikán col.— 1 male and 1 female (FIOC); Juruti, without date and collector— 1 male and 3 females (CEMT); Óbidos, XII.1953, J. Brazilino col.— 1 male (MNRJ); Santarém, 27- 28.XII.1967, Exp. Perm. Amaz. col.— 1 male (MZSP). RONDÔNIA: Porto Velho, Rio Madeira, X.1932, B. Pohl col.— 2 males and 2 females (MZSP). SANTA CATARINA: Rio Vermelho, XII.1961, Dirings col.— 11 males and 7 females (MZSP); Rio Vermelho, IV.1963, Dirings col.— 3 males and 4 females (MZSP). COLOMBIA: 1905, Fry col.— 1 male and 1 female (BMNH); 1910, Fry col.— 1 male (BMNH). AMAZONAS: Letícia, 700 ft., 08- 09.VIII.1970, H. & A. Howden cols.— 1 male and 1 female (CMNC); Letícia, 700 ft., 19-25.II.1972, H. & A. Howden cols.— 1 male (CMNC); Letícia, 700 ft., 23.II-02.III.1974, H. & A. Howden cols.— 1 male and 1 female (CMNC). CALQUETÁ: Vegas Del Calquetá, without date, O. Rojas col.— 1 female (MIZA). HUILA: Gigante, VII.1979, O. Rojas col.— 6 males and 9 females (MIZA); Gigante, 4.VI.1981, O. Rojas col.— 1 male (MIZA); Gigante, parte alta da cordilheira, without date, O. Rojas col.— 1 female (MIZA); Gigante, parte alta da cordilheira, 1981, O. Rojas col.— 1 female (MIZA); Gigante, parte alta da cordilheira, 06-07.I.1982, O. Rojas col.— 1 male (MIZA); Gigante, sopé da cordilheira, whitout date, O. Rojas col.— 1 male (MIZA). ECUADOR: NAPO: Rio Coca, IX.2000, D. J. Inward & K. A. Jackson cols.— 1 male (BMNH). PERU: UCAYALI: Coronel Portillo, Pucallpa, XII.1978, without collector— 1 female (CEMT); Coronel Portillo, Pucallpa, X-XI.1992, L. E. Peña col.— 1 male and 2 females (CEMT); Coronel Portillo, Pucallpa, Rio Ucayalli, 200m., IX.1947, without collector— 1 male (MZSP); Coronel Portillo, Yarinacocha, 150 m., 8-15.VII.2001, D. Curoe col.— 4 males and 4 females (CEMT); Coronel Portillo, Yarinacocha, Rio Ucayali, 11.X.1946, J. Schunke col.— 1 female (MNRJ) and 2 females (MZSP).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.