
Parvidrilus meyssonnieri, 2012
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00857.x |
|
DOI |
https://doi.org/10.5281/zenodo.5480034 |
|
persistent identifier |
https://treatment.plazi.org/id/039ADA53-1F5F-697E-FF53-24F26671FC91 |
|
treatment provided by |
Marcus |
|
scientific name |
Parvidrilus meyssonnieri |
| status |
sp. nov. |
PARVIDRILUS MEYSSONNIERI DES CHÂTELLIERS & MARTIN SP. NOV. ( FIGS 1 View Figure 1 , 2 View Figure 2 )
Types: Holotype. MHNL 44003358 View Materials , slide 6, mature specimen, stained in paracarmine and wholemounted in Canada balsam. Gallery of ‘ La Martinière’ (45°40′51′′N, 04°38′51′′E, 350 m asl), Thurins (69), France, 28.ix.2000. GoogleMaps
Paratypes. MHNL 44003359 View Materials , slide 1, six specimens, four immature, one mature and one fragment ; MHNL 44003360 View Materials , slide 3, four specimens, two mature and two immature; slide 6, one immature specimen; all specimens stained with paracarmine and wholemounted in Canada balsam; 28.ix.2000; gallery of ‘ La Martinière’, Thurins (69), France .
Other material examined: Apart from the type material, 11 specimens were observed and mounted on five scanning electron microscope ( SEM) stubs, IRScNB, 28.i.2008; stub 1, three specimens, type locality; stubs 2–5, eight specimens, gallery of ‘Navogne’ (45°17′16′′N, 04°05′58′′E, 520 m asl), Bas-en-Basset (43), France. Thirty-six specimens were wholemounted in Canada balsam or in Amman’s lactophenol, in the collection of one of the authors (MCdC): ‘ La Martinière’ (seven specimens, four mature and three juvenile, 14.ix.1999; 24 specimens, ten mature and 14 juvenile, 28.ix.2000); ‘Navogne’ (three immature specimens, 8.x.1999; one mature specimen, 28.i.2008) GoogleMaps .
Etymology: Named after Marcel Meyssonnier, as a tribute to this outstanding and tireless speleologist, and long-time friend and team member of one of the authors (MCdC), and whose involvement and support were instrumental in the discovery of this new species.
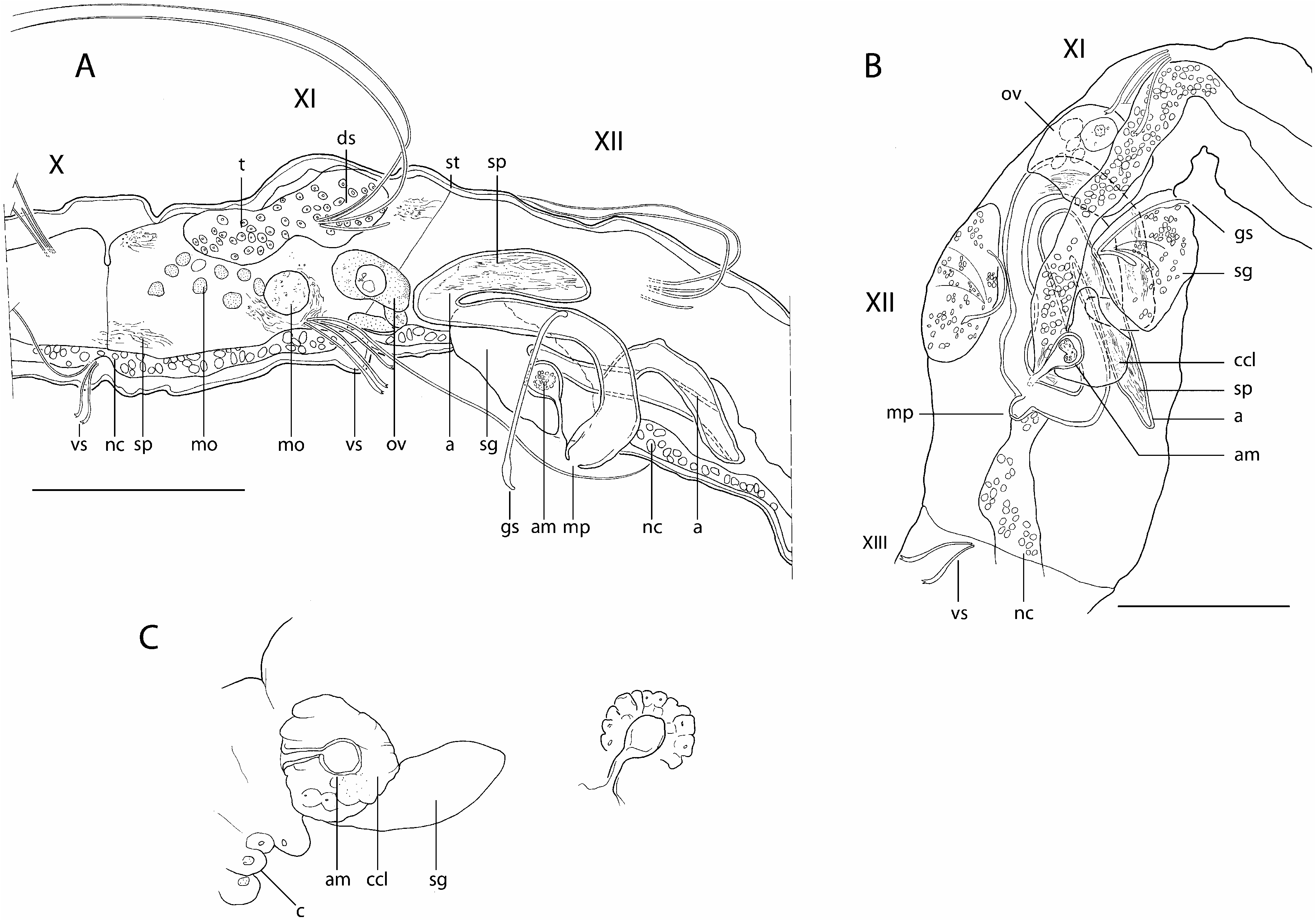
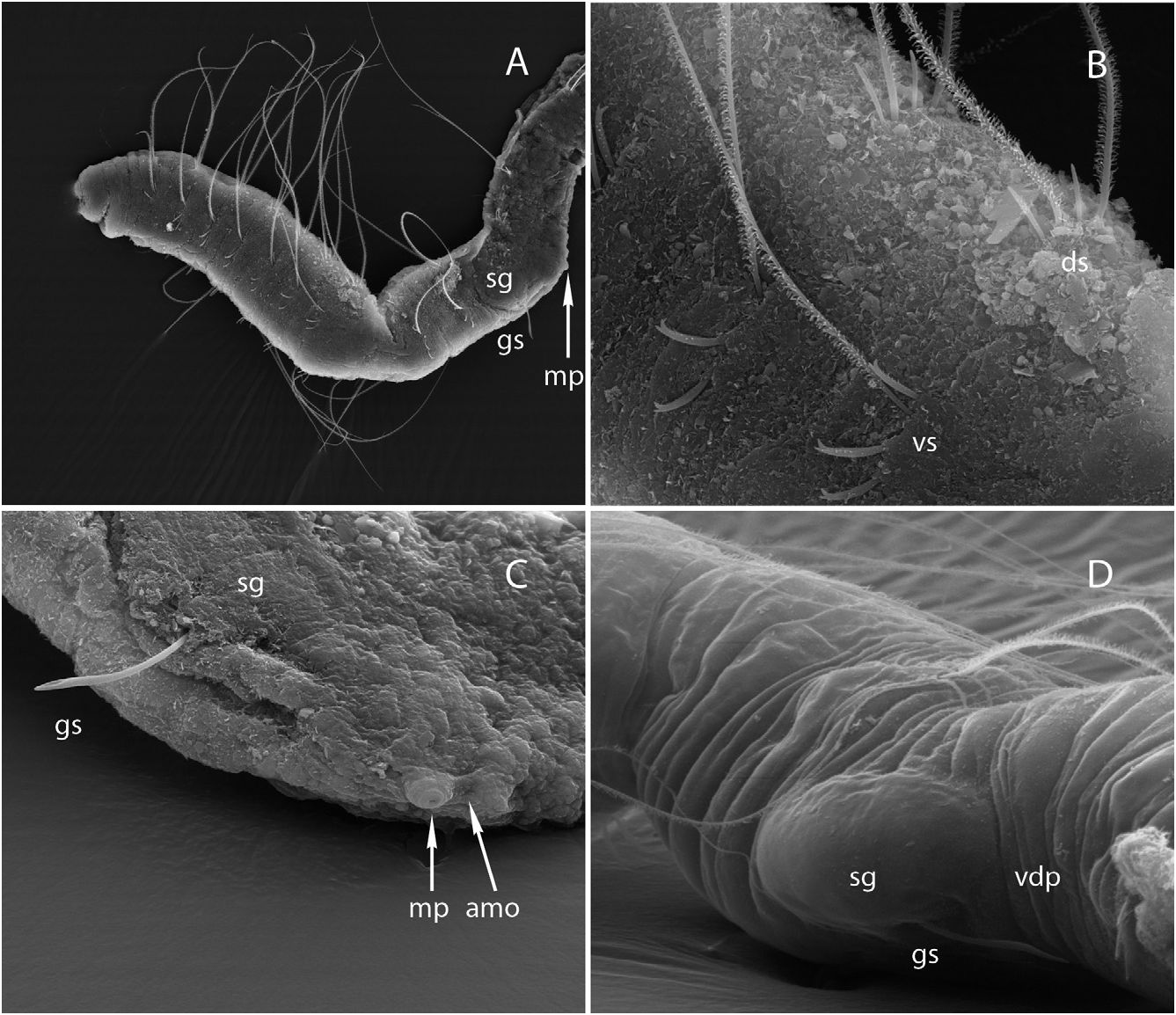
Description: Length 1.3-1.6 mm, 21–30 segments (seven complete mature specimens). Width 55–85 Mm at V (mean = 59 Mm ± 11, N = 24), 50–120 Mm at XII (mean = 72 Mm ± 19, N = 24). Prostomium rounded, 35–50 Mm long, 20–38 Mm wide at base. Body wall not papillate, cuticle smooth. Clitellum poorly developed, limited to large, swollen, transparent cells, only visible on living animals ( Fig. 1C, c View Figure 1 ), budding on a mid-ventral depression of body wall ( Fig. 2D View Figure 2 , vdp) that extends behind genital pores, between ventral setae in XII and XIII.
Dorsal (dorsolateral) and ventral (ventrolateral) setae present from III, posteriorly situated in each segment ( Figs 1A, B View Figure 1 , 2B View Figure 2 , ds, vs). Dorsal bundles with two (three) straight, thin single-pointed needles (15– 28 Mm long), distal end bluntly pointed, alternating with (one) two thin and flexible hair setae (length 150–230 Mm at V, 200–260 Mm at VIII, 170–225 Mm at XI). Hair setae densely pilose (with soft hairlets) on one side of the shaft (pilosity mostly observed on specimens during study using SEM). Ventral bundles with (two) three (four) sigmoid, bifid crotchets with upper tooth slightly thinner and shorter than lower tooth, and accompanied by one (two) pilose hair setae in preclitellar segments (80–170 Mm long); ventral hair setae absent from XII; crotchets without nodulus, 17–28 Mm long, shaft out of the body wall faintly pilose, with distal half enlarged on the convex side. Within ventral bundles, the uppermost seta bifid, followed by one hair seta; these two setae separated from the other two bifid setae by a distance sometimes greater than that between them. One modified genital seta per bundle in XII, 55–60 Mm long, straight with enlarged, spearhead ectal tip, the latter with a spoon-like curvature ( Figs 1A, B View Figure 1 , 2A, C, D View Figure 2 , gs).
No eyes. Brain long, extending into IV, posteriorly indented. Ventral nerve cord in contact with epidermis, beneath muscles of the body wall. Digestive tract complete; eversible pharynx with dorsal pad, sinuous, ciliated oesophagus, with pharyngeal glands present in IV- VI, followed by a simple intestine, slowly dilating from X- XI and ending in a terminal anus. Layer of chloragogen cells surrounding digestive tract possibly present; details not observable in specimens thus far examined. Coelomocytes absent. Dorsal glandular body-wall pouches very small and difficult to observe, but observed in VIII and XII in one specimen), opening mid-dorsally at about the intersection with setal line. Voluminous glands associated with genital setae (20–48 Mm long; 20–32 Mm wide), bulging at the body surface; fine internal ducts running inside the glands and converging on the shaft of genital setae; setal glands asymmetrically arranged on each lateral side of the body, the left gland anteriad to the right one ( Figs 1A–C View Figure 1 , 2B View Figure 2 , sg).
Clearly delimited testes not present; however, a lump of cells with small nuclei observed dorsally in XI, in posterior end of coelom of XI, is probably unpaired male protogonia ( Fig. 1A, t View Figure 1 ). Diffuse formation of male gametes in XI with many morulae (or ‘cysts’ according to Ferraguti, 1997), 2.8–4.0 Mm wide, freely floating inside the coelomic cavity although usually lying ventrally in XI ( Fig. 1A View Figure 1 , mo); a few globular cytophores in a more dorsal and posterior position in XI, 8.0–11.3 Mm wide, sometimes in a stage of disintegration with the detachment of spermatozoa in the coelom. Sperm funnels and vasa deferentia not seen. Atria elongate (90–190 Mm long, 7–10 Mm wide), either extending into XIII or restricted to XII; in the latter case, atria folded in different configurations: one atrium bent anteriad, with distal end folded up posteriad, the other atrium (or if present, both atria), bent posteriad, with distal end folded up anteriad ( Fig. 1A, B, a View Figure 1 ). Atria merging below the nerve cord ( Fig. 1A, B View Figure 1 , nc) into a common ejaculatory duct, and opening at the tip of a mid-ventral, conical porophore ( Figs 1A, B View Figure 1 , 2A, C View Figure 2 , mp), in posterior part of XII, between setal glands and in front of a median ventral depression of body wall in XII- XIII ( Fig. 2D View Figure 2 , vdp). Atrial ampullae longitudinally striated, with large lumen filled with sperm, mature spermatozoa more concentrated near the distal part of ampullae ( Fig. 1A, B View Figure 1 , sp); no peculiar aggregate of sperm in the ampullae. An (unpaired?) ovary, apparently developing on one side, observed in anterior part of XII, with a ventral attachment on 11/12 ( Fig. 1A, B View Figure 1 , ov). Specimen ovigerous with large mature eggs filling coelom in XII, and extending through XIII. Female funnels not observed. No typical spermatheca observed. However, an unpaired, spherical ampulla in XII, 10 Mm in diameter, ventrally located, ahead of the point of union of atrial ampullae, usually on the right side of the ventral nervous chord but sometimes on the left side in some specimens ( Fig. 1A–C View Figure 1 , am); ampulla opening, through a duct, 14 Mm long, mid-ventrally, close to the basis of male porophore ( Fig. 2C View Figure 2 , amo); duct surrounded by a muscular ring near opening. Spherical ampulla filled with cell material (sperm?) and surrounded by a cell cluster, clearly delimitated, noticeable on living worms but difficult to resolve on mounted slides, possibly glandular ( Fig. 1B, C View Figure 1 ).
COI sequences: EMBL accession numbers: HE800204 View Materials (gallery of ‘ La Martinière’); HE800205 View Materials (gallery of ‘ Navogne’) .
Remarks: Within the parvidrilid species known so far (including all new species being described herein plus the reassigned Parvidrilus gineti – see below), the presence of genital setae is unique to P. meyssonnieri sp. nov. – clearly separating this species from its congeners. The most outstanding feature in P. meyssonnieri sp. nov. is the presence of an enigmatic unpaired, spherical ampulla in XII – the function of which remains obscure. There is a possibility that this structure is used for storing sperm from a concopulant, which would imply that it is a spermatheca. However, the ampulla is surrounded by a cell cluster, which seems more glandular (prostate-like cells) than shaped in order to play the role of a bag for sperm storage. In this respect, such a structure is somewhat similar to the glandular, atrium-like organ in the pre-atrial segment of some Rhynchelmis (Rhynchelmis) species ( Lumbriculidae ), referred to as ‘accessory organ’ (formerly ‘rudimentary atrium’), whose function and derivation are unknown ( Fend & Brinkhurst, 2010). Some material has been observed in the spherical ampulla but it does not resemble mature spermatozoa. Hence, the absence of spermathecae in this species seems plausible. Conversely, considering the ampulla and its surrounding cell cluster as an atrium and a prostate is another possibility – implying that the structures described above as ‘atria’ are in fact the spermathecae and the porophore is spermathecal. However, prostates seem to be absent in Parvidrilus (see below). In addition, when copulatory organs are present in microdriles – whether in the form of porophores or penes – they are always associated with the male aperture ( Stephenson, 1930), thus making the latter hypothesis less convincing.
Distribution and habitat: Parvidrilus meyssonnieri is known only from the ‘La Martinière’ and ‘Navogne’ galleries in the Rhône department, France, in a region lying on metamorphic bedrock. The ‘La Martinière’ gallery opens in the upper alteration layer of a crystalline craton, which forms granitic sand with scarce, particulate organic matter. A rivulet runs along the gallery and passes slowly through a thin layer of coarse sand. Historically (between 1846 and 1919) the Navogne gallery was dug in arkoses and ferruginous sandstone for iron ore exploitation. The water is present in the gallery as isolated puddles with gravelly soil, rich in organic matter.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |