
Pheretima (Pheretima) darnleiensis ( Fletcher, 1886 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.179016 |
|
DOI |
https://doi.org/10.5281/zenodo.6240504 |
|
persistent identifier |
https://treatment.plazi.org/id/03FF4131-275B-FFD7-0C8C-C418FB3EF9D3 |
|
treatment provided by |
Plazi |
|
scientific name |
Pheretima (Pheretima) darnleiensis ( Fletcher, 1886 ) |
| status |
|
Pheretima (Pheretima) darnleiensis ( Fletcher, 1886)
Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Synonymy:
Perichaeta cingulata (part, cf. A. robustus ); Vaillant, 1868a: 225, 228; Vaillant, 1868b: 146 – misidentifications. [Non Perichaeta cingulata Schmarda, 1861: 14 , fig. 163 (= Megascolex cingulatus from Sri Lanka as redescribed by Beddard, 1892: 122, figs. 9–13; 1895: 382; Michaelsen, 1900: 229; Stephenson, 1923: 236, fig. 86)].
Perichaeta indica: Horst, 1885: 4 (syn. cingulata: Vaillant – misidentification). [Non Megascolex indicus Horst, 1883: 186 (= Amynthas corticis Kinberg, 1867 according to Easton, 1981: 49)].
Perichaeta Darnleiensis (sic) Fletcher, 1886b: 966. [Dated “ 1886 ” e.g. by Michaelsen (1900) or dated “ 1887 ” by Sims & Easton (1972: 239, 260); Easton (1983: 732; 1984: 119). “ The part containing Fletcher's paper was actually published on 17 November 1886 ” – M.L. Augee, Editor, Proc. Linn. Soc. NSW (pers. comm. to RJB, Oct. 2004)]. From Darnley Island, Torres Straits, Australia. Types missing.
Perichaeta vaillanti Beddard, 1890: 66 ; Beddard, 1895: 422 [Beddard (1895: 421, 422) thought Ph. ( Ph.) philippina (Rosa, 1891) similar except for its two pairs of spermathecae, and Amynthas enganensis (Rosa, 1892) similar except for its longer spermathecal diverticula]; Michaelsen, 1900: 311. From Manila, Luzon, Philippines. Type in British Museum: BMNH 1904:10:5:173.
Perichaeta martensi Michaelsen, 1892: 242 ; Horst, 1893: 324; Beddard, 1895: 416 (?syn. darnleiensis ); Michaelsen, 1896: 240 (syn. eoa ); 1900: 282 (syn. javana, eoa ). From Banka Island east of Sumatra. Types in Berlin: 2148.
Perichaeta bosschae Horst, 1893: 324 ; Beddard, 1895: 432; Michaelsen, 1896: 238; 1900: 256; [the two specimens Beddard (1900b) mis-identified as P. b os sch ae were given the new name of P. baruana Stephenson, 1942: 209 , later included in synonymy of Metaphire malayana ( Beddard, 1900b) by Gates (1972: 199)]. Name misspelt “ boschae ” by Michaelsen (1896: 238). From Borneo (Baram River, Sambas). Type in Leiden Museum: 1829.
Perichaeta martensi javana Michaelsen, 1894: 99 . [ Sims & Easton (1972: 260) misspelt Michaelsen’s martensi javana as “ javanica ” [non Rhodopis javanica Kinberg, 1867 (= Metaphire javanica )]. From Java. Types missing.
Perichaeta darnleiensis: Beddard, 1895: 406 .
Perichaeta padasensis Beddard & Fedarb, 1895: 73 . From Merabah, Padas Valley North Borneo. Types in British Museum: BMNH 1904:10.5:119–26, 1263–4.
? Perichaeta atheca Rosa, 1896a: 520 ; Michaelsen, 1900: 254; 1928: 41; Sims & Easton, 1972: 223 [who maintained it as species incertae sedis yet noted that Michaelsen (1928: 41) “ regarded it as a subspecies of indica (= Pheretima (Pheretima) darnleiensis )”. From Sumatra (Balighe, Si-Rambe and Lake Toba). Types in Genoa: 44026-28-33.
Perichaeta eoa Rosa, 1896a: 528 ; Rosa, 1896b: 609. Types in Genoa (44009-11) and Turin (151).
Perichaeta floweri Benham 1897: 217 . [Spermathecal pores in 5/6/7/8/9; setae 35–45 and male pores superficial according to Michaelsen (1900: 267) but which were “ doubtless ” in copulatory chambers according to Gates (1934: 259) in his inspection of syntypes where he also noted the “ finger-like ” ampullae of the seminal vesicles (see description under Pheretima floweri below)]. From Bukit Timah, Singapore. Types in British Museum: BMNH 1896.9.30.9-11. These three softened syntypes were inspected by Gates (1934: 258) and also by Sims & Easton (1972: 180).
Perichaeta madelinae Benham 1897: 219 . [Described by Michaelsen (1899: 76; 1900: 291) as either Amyntas or Pheretima padasensis madelinae (Benham) ; Reynolds & Cook (1976: 132) mistakenly attribute this taxon to Michaelsen (1899 - see below) as well as to Benham (1897)]. From Mt Kinabalu , North Borneo. Types in British Museum: BMNH 1896:9:30:4.
Perichaeta belli Rosa 1898a: 286 . From Mindoro, Philippines. Type in British Museum: BMNH 1898:10:28.5.
Megascolex cingulatus (part cf. A. robustus ): Vaillant, 1889: 72.
Amyntas padasensis lokonensis Michaelsen, 1899b: 74 [miscited by Beddard (1900a: 615) as page “ 78 ”. Differs from madelinae by smaller size (ca. 45–68 mm) and having fewer setae (ca. 36)]. From Mt Lokon, Tomohon, North Celebes. Type missing from Basel Museum.
Amyntas padasensis madelinae: Michaelsen, 1899b: 76 . [Differs from lokonensis by larger size (ca. 135 mm) and having greater setae (ca. 58)].
Amyntas padasensis padasensis: Michaelsen, 1899b: 74 . [Cited by Michaelsen (1900: 290) as P. padasensis (typica) and differentiated from its sub-species by size> 200 mm (ca. 280 mm) and having extra seminal vesicles in segment 13].
Amynthas martensi: Michaelsen, 1899b: 87 .
Amyntas cingulatus: Vaillant (in part); Beddard, 1900a: 615 [miscited by Sims & Easton (1972: 261) as page “ 625 ”]; [syns. darnleiensis , vaillanti , martensi , indica Horst, eoa , madelinae , padasensis var. lokonensis , belli ; also in a footnote Beddard (1900a: 615) says that Perichaeta cingulata of Schmarda is of course a Megascolex as then defined]; Beddard, 1900b: 892 [syns: indica (part), darneleiensis, vaillanti , martensi martensi , m. javana, eoa , madelinae , belli , padasensis lokonensis ). Many of these synonyms were accepted by Sims & Easton (1972) who, however, also added padasensis and bosschae ].
Amynthas padasensis: Beddard, 1900a: 624 .
Amynthas bosschae: Beddard, 1900a: 625 ; non Beddard, 1900b: 892 (= M. baruana ).
Amyntas peregrinus (?part inc. floweri ): Beddard, 1900a: 644.
Pheretima belli: Michaelsen, 1900: 255 .
Pheretima bosschae: Michaelsen, 1900: 256 .
Pheretima darnleyensis (sic, lapsus): Michaelsen, 1900: 263 (illegal emendation).
“ Pheretima darleyensis (sic): Michaelsen, 1900, p. 267” reported by Easton (1982: 732) is perhaps a lapsus for the above lapsus.
Pheretima floweri: Michaelsen, 1900: 267 .
Pheretima indica (part): Michaelsen, 1900: 275; Michaelsen, 1910a: 260; Michaelsen, 1922: 41 (syn. kuchingensis ); Michaelsen, 1923: 2; Michaelsen, 1928: 40 (part inc. indica typica = padasensis ); Stephenson, 1930b: 1; Stephenson, 1931b: 273; Michaelsen, 1932a: 614; Stephenson, 1932b: 219; Ude, 1932: 147 (part inc. typica); Michaelsen, 1935: 108; Gates, 1935b: 82, 83 (syn. floweri ), 94; Gates, 1936c: 389 (syn. quadragenaria (part) s. Michaelsen, 1924); Gates, 1937a: 320; Gates, 1937d: 193; Gates, 1940: 413; Gates, 1949: 26. [Note: Gates (1935b) was referring to a concept of P. indica ( Horst), 1883 that Gates later re-described as being P. darnleiensis (i.e., not as indica (= A. corticis )].
Pheretima martensi: Michaelsen, 1900: 282 (syn. m. javana, eoa ).
Pheretima padasensis typica: Michaelsen, 1900: 290 .
Pheretima padasensis lokonensis: Michaelsen, 1900: 290 .
Pheretima padasensis madelinae: Michaelsen, 1900: 291 .
Pheretima vaillanti: Michaelsen, 1900: 311 .
Pheretima decipiens Beddard, 1912: 180 . Syn. nov. [Beddard said this species, which he described lengthily but poorly, seems to be “ exactly like P. benguetensis ”, he failed to give any convincing argument for their separation and, moreover, since Ph. benguetensis was listed in synonymy of Ph. darnleiensis by Sims & Easton (1972) then Ph. decipiens most probably belongs too. One specimen (of how many?) had only three pairs of spermathcae]. From Luzon, Philippines. Types unknown.
Pheretima benguetensis Beddard, 1912: 183 , fig. 1. [This name was maintained in synonymy and provisionally separated in a pheretimoid checklist by Blakemore (2004b: 133) because it was erroneously listed in a publication by Nakamura (1999: 27) as having only two pairs of spermathecal pores in 6/7/8 – this now known to be a mistake with reference to Beddard’s original description. Length to 190 mm]. From 1,524m in the Province of Benguet, Luzon, Philippines. Types unknown.
Pheretima kuchingensis Stephenson, 1916: 337 . From Kuching, Sarawak, Malaysia. Type in Calcutta: 6539.
Pheretima quadragenaria (part): Michaelsen, 1924b: 391. [Note: Perrier’s species was originally described with only one pair of spermathecae in 7/8].
Pheretima borneoensis Ude, 1925: 108 ; Michaelsen, 1928: 55 (syn.? bosschae ); Ude, 1932: 131; Michaelsen, 1934b: 45. From Borneo. Types in British Museum: BMNH 1933:10:6:26.
Pheretima indica birangi Michaelsen, 1928: 42 . From Borneo. Types missing.
Pheretima floweri: Gates, 1934: 258 ; [Gates, inspected the three macerated British Museum syntypes with this label “ Perichaeta floweri . [18]96.9.30 9–11. Bukit Timah, Singapore. S.S. Flower”; one specimen was useless and was discarded, his redescription was based on the external characters of an undissected specimen and the internal characters of both remaining specimens were both rather nondescript].
Pheretima padasensis: Kobayashi, 1941: 397 .
Pheretima (Pheretima) darnleiensis: Sims & Easton, 1972: 260 [syn. vaillanti , martensi , m. javanica [lapsus pro. javana], bosschae , padasensis , p. lokonensis, p. madelinae , eoa , floweri , madelinae , belli , kuchinensis, quadragenaria (part), benguetensis , borneoensis , indica birangi ]; Easton, 1982: 732; Easton, 1984: 119 (syn. indica: Gates, 1937: 320 ); Blakemore, 1994: 368 ( vaillanti , martensi , bosschae , padasensis , eoa , floweri , madelinae , belli , etc.); 2002: 216 (syn. vaillanti , martensi , bosschae , martensi javana , padasensis , eoa , floweri , madelinae , belli , padasensis lokonensis , benguetensis , kuchingensis , borneoensis , indica birangi ); 2004: 136 (syn. vaillanti , martensi , m. javana, cingulatus (part), bosschae , padasensis , p. lokonensis, p. madelinae , eoa , floweri , belli , kuchinensis, quadragenaria (part), benguetensis , borneoensis , indica birangi ).
Diagnosis: Pheretima usually with four or, rarely, five (or 3?) pairs of spermathecae in (4/5)5/6-8/9 about 0.2 circumference apart ventrally. Male pores on 18 ca. 0.2 circumference apart in slit-like copulatory pouches. Genital markings lacking. Intestinal caeca in or near 27, simple (or with slightly complex margins). Size range 45–400 (and up to 700 mm in life) by 2.5–6.0 mm.
Distribution: the tropical Indo-Australasian Archipelago and islands: Malaysia, Singapore, Berhala Island east of the Malay Peninsula, Borneo, Brunei, Indonesian Islands: e.g. Sereina (Sipora) of the Mentawei Islands west of Sumatra [(Rosa, 1896b) reported by Sims & Easton (1972: 263) as “ Metawei Is. ”), Krakatau Island, Sumatra, Java, Bali; Australian Territory of Christmas Island south of Java [e.g. by Michaelsen (1935) and Gates (1935b: 94)], Philippines, North Celebes, Moluccas, Caroline Islands of Micronesia north of Papua New Guinea (Kobayashi, 1941), Fiji [Levuka, Ovalau – ( Gates, 1937c) as noted by Sims & Easton (1972: 263)], Kei Islands in the Arafura Sea just west of West Papua or Irian Jaya [by (Michaelsen, 1910a) as cited by Sims & Easton (1972: 263)], and Darnley Island in the Torres Straits – after whence it was named but which is unlikely to be its true origin. Whereas, Easton (1982: 732) thought that records from Caroline, Kei, Fiji, and Darnley islands (at least?) were the result of introductions, Sims & Easton (1972: 263) are dismissive of reports from Sri Lanka, Manila [Vaillant (1889) cf. Perichaeta vaillanti ], and South Africa. Moreover, historical reports from Hawaii and Brazil of “ Perichaeta indica Horst ” are more likely to refer to Amynthas corticis than to Ph. darnleiensis proper. Full lists for species from various regions, including the Pacific and ‘Papuasia’, are presented in Blakemore (2005; 2006a).
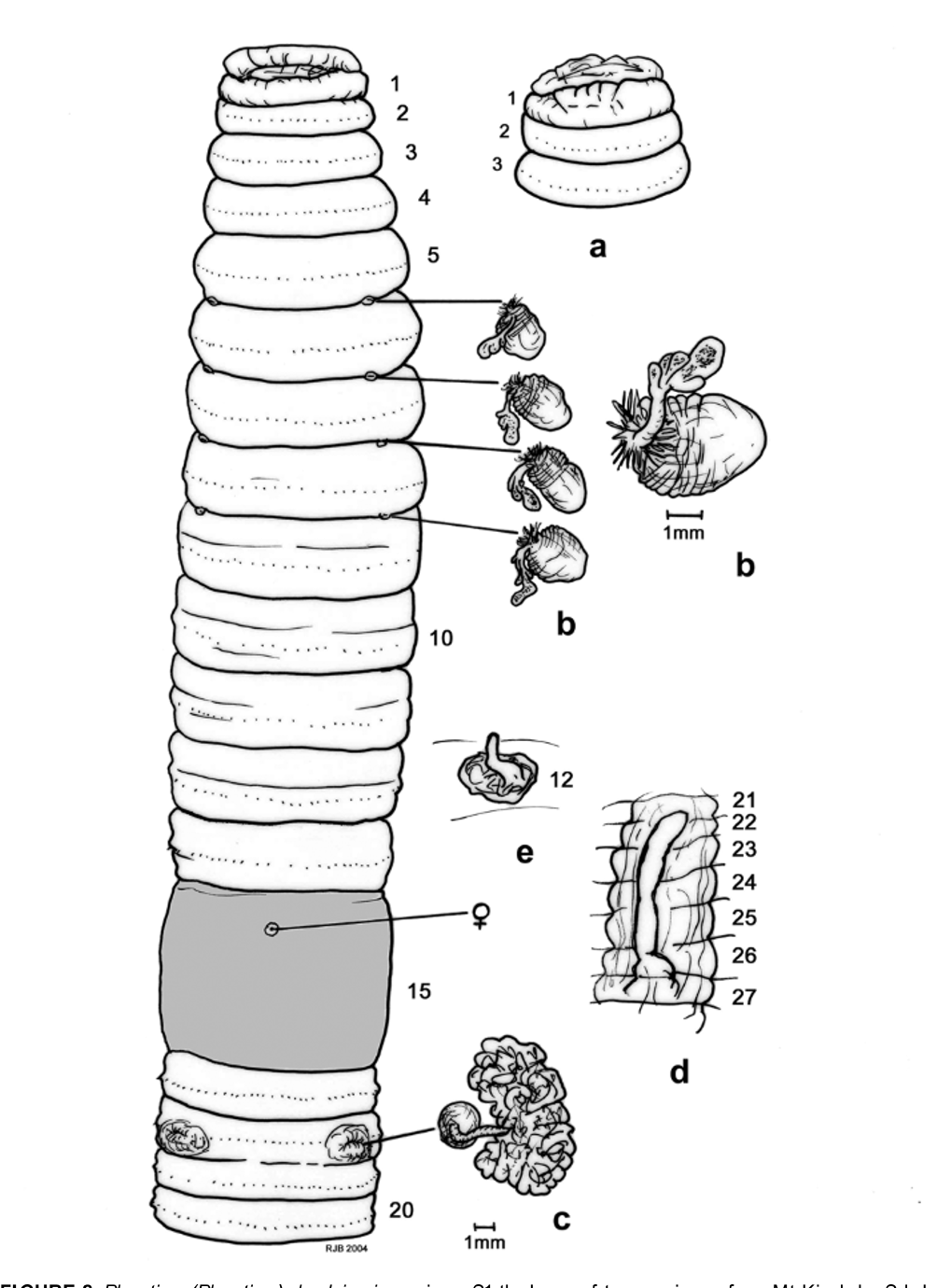
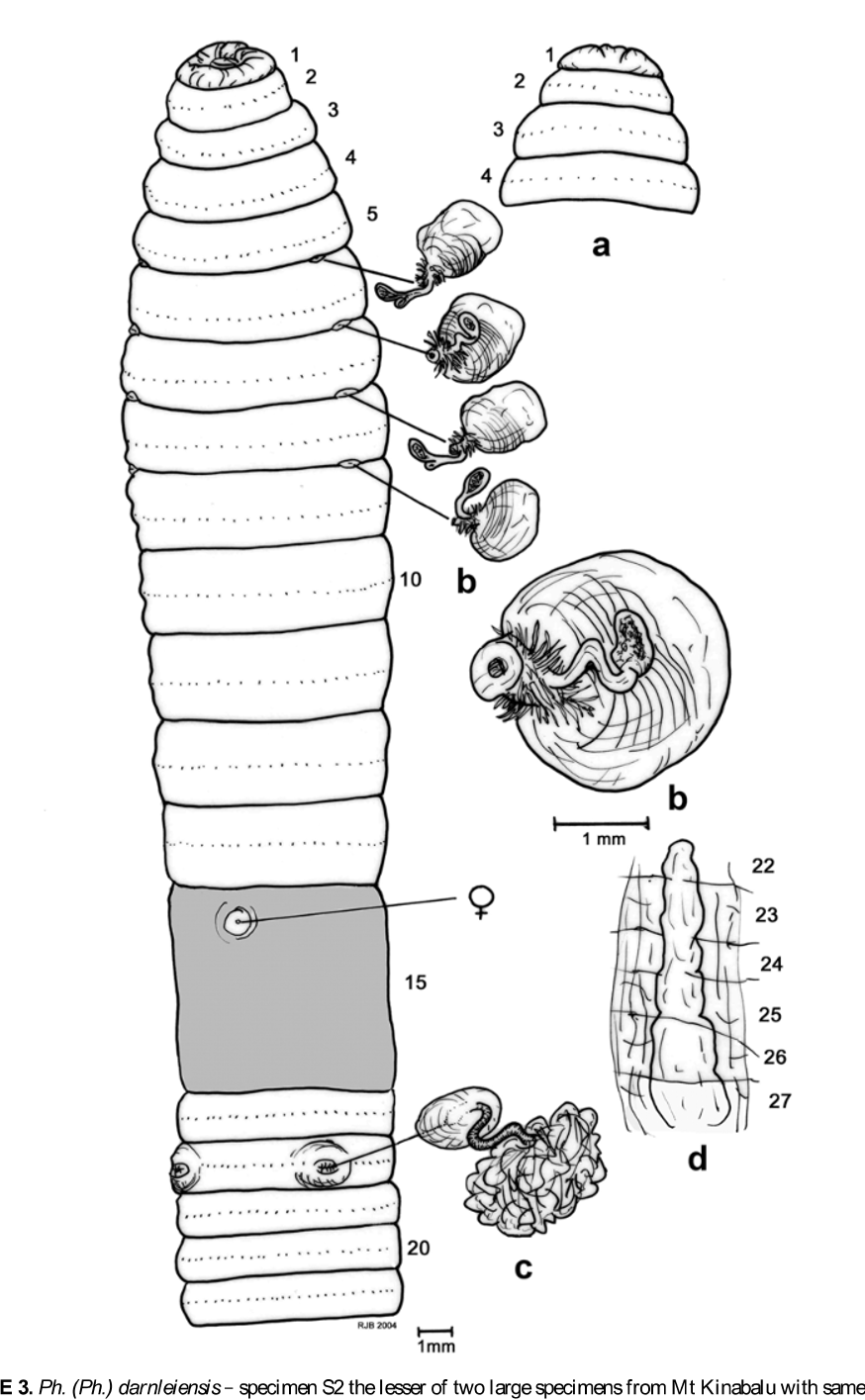
Material examined: two mature specimens (both dissected and figured by RJB, the larger one at 385 mm designated as S1, the shorter one at 305 mm, S2), from Mt Kinabalu , Sabah, Malaysia collected by Dr M.T. Ito in 2003; several smaller less mature specimens collected from Mt Kinabalu by T. Kawaguchi on 16– 17.ix.2004 (e.g. samples labeled: “ 040916 PAKA 3-C”, “ 040917 HELI 5-B”), plus numerous immature specimens collected from lowlands East of Mt Kinabalu by T. Kawaguchi (e.g. those labeled: “ 060305 T3-3”, “ 060302 R-1-15” and “ 060302 R-1-2-3”) that although aclitellate look like striped and more regularly coloured forms of Pheretima ( Ph.) darnleiensis . All specimens held in YNU SERG Soil Ecology laboratory collection in Yokohama pending museum deposition.
Length: range 45–385 mm [i.e., reported as 108 mm (Fletcher), 45–280 mm for sub-species of padasensis , 111–155 by 6 mm ( Michaelsen, 1900) and 75–170 mm ( Sims & Easton, 1972)]. Largest current specimens measure (S1) 385 and (S2) 305 mm, respectively, with the smaller specimens ca. 100–140 mm long. In life, specimens are longer – approaching 700 mm when stretched out (T. Kawaguchi and M. Schilthuizen pers. obs., and weblink http://en.wikipedia.org/wiki/ Kinabalu _giant_earthworm Feb. 2007).
Width: 2.5–6.0 mm mostly commensurate with length.
Segments: 70–180 [155 by Fletcher, repeated by Beddard (1895: 406), 79–108 by Michaelsen or 70–120 by Sims & Easton]; current specimens had 127 (S1 with possible posterior regrowth) and up to 180 segments in (S2).
Colour: blue-grey (natural) or brown (preserved) with darker clitellum (cf. some immature striped specimens from east of Mt Kinabalu were superficially similar, as noted above).
Prostomium: open epilobous.
First dorsal pore: 12/13 (S1) or, less often 11/12 as in (S2).
Setae: numerous, especially in posterior, ca. 25–66 per segment, no ventral gaps nor crowding; variously 4–14 setae intervene between male pores. The original description had 60–66 setae per segment (as also for madelinae and belli , cf. 35–45 for floweri ); current large specimens have 60–70 setae in anterior segments, whereas in smaller specimens 40–60 setae were counted.
Nephropores: not detected.
Clitellum: annular 14–16, setae occluded.
Male pores: on 18 ca. 0.2 circumference (or ca. 3 mm in larger specimens) apart in slit-like copulatory pouches on distinct round porophores with intervening setae numbering 4–8 in smaller specimens to about 14 in larger current specimens.
Female pores: single (or closely paired?) on 14.
Spermathecal pores: usually four, rarely five pairs in (4/5),5/6-8/9 (?or three pairs in 6/7/8/9) ca. 0.2 circumference apart ventrally approximately in line with 10th setae on each side in current specimens.
Genital markings: none.
Septa: 4/5-7/8 thickened; 8/9 membranous (e.g. S2) or absent (S1 and some current smaller specimens), 9/10 absent; 10/11+ thickened.
Dorsal blood vessel: single.
Hearts: hearts 10,11–13; supra-oesophageal vessels present.
Gizzard: between 7/ 8/9 and 10/11; in (S1) occupying 9–10 with equally large ‘crop’ in 7–8.
Calciferous glands: oesophageal pouches absent (but some slight dilations in 11 and 13).
Intestine origin (caeca, typhlosole): 15,½15 (S1, S2) or 16 (some current smaller specimens); caeca “ simple or with complex margins ” ( Sims & Easton, 1972) but only simple form found in current large specimens or slightly incised ventrally in smaller specimens, paired in 27,26–24,22,21; simple lamellar typhlosole present commencing from about 25,26–28 (new observation on current specimens). Gut contains loamy soil.
Nephridia: meroic; dense, tufted (?pepto-) nephridia on posterior septa in 5 and 6 (S1 and smaller specimens), tubules also present on spermathecal ducts, numerous and diffuse elsewhere.
Testes/sperm funnels: holandric, testes in 10 and 11 in testis sacs, seminal vesicles – each with a distinct finger-like dorsal extension – paired in 11 and 12; elongate pseudovesicles present in 13 (seen in S1, S2 and current smaller specimens that also had smaller pseudovesicles in 14), sometimes reported as “ coelomic sacs ” on 12/13 and 13/14 ( Sims & Easton, 1972: 262) [or as “ pseudovesicles ” in 13 and 14 by Gates (1934)].
Ovaries: paired in 13 with densely packed bunches of egg strings; ovisacs not found.
Prostates: large racemose in 17–19, ducts pass into copulatory pouches that each contains a conical penis and an anterior and posterior secretory pad; the coelomic surface of each pouch has small glandular masses above each pad.
Spermathecae: paired in 6–9 (sometimes reported in 5–9); small, duct short and stout and coated with nephridial tubules; diverticulum ectal in origin and typically clavate (in lesser specimens) but sometimes with additional (two or three) lobes to its bulb that is equal in length to the stalk [or “ ampulla ” and “ duct ” by Sims & Easton (1972: 262) although ‘ampulla’ and ‘duct’ usually refer to the main structures of the spermatheca and on the diverticular appendage are more correctly known as the ‘stalk’ and ‘bulb’].
Other: paired septal glands noted at septal locations on intestinal along dorsal blood vessel (S1). Many nematodes found in coelom of (S2).
Remarks. Fletcher (1886b: 967) first reported “ testes ” in the unusual position of 11 and 12, but later (1887b: 603) corrected these to seminal vesicles. Sims & Easton (1972: 264) found extra spermathecae only infrequently in a few of their Mt Kinabalu specimens but decided not the give them separate taxonomic status. Variations in the intestinal caeca from typically simple to lobate (e.g. in borneoensis or bosschae ) were correlated with growth of specimens and were not deemed of taxonomic value by Sims & Easton (1972: 264) who also thought the glandular masses on the copulatory pouches were possibly homologous to the secretory diverticula diagnostic of Parapheretima. Orginally P. padasensis defined specimens with bi- or tri-lobed spermathecal diverticula, but single lobed specimens were also allowed by Michaelsen (1899; 1900) and by Gates (1935: 83).
Sims & Easton (1972: 266) note that holandric, decathecal Metaphire fordi (Michaelsen, 1934) may be confused with Ph. darnleiensis , although M. fordi lacks nephridia on its spermathecal ducts, and Ph. darnleiensis lacks genital markings. Decathecal but metandric species that may be compared are Metapheretima loriae (Rosa, 1898) from New Guinea, the Solomons, and Vanuatu and Me. pickfordi (Gates, 1959) from the Solomons and Vanuatu – both as redescribed by Lee (1981: 554) and both lacking intestinal caeca.
Approximately 140–152 pheretimoids have spermathecal pores in 5/6/7/8/9 ( Blakemore, 2006a and unpublished data) although about 36 of these are Metaphire . Similar holandric, octothecal species are Metaphire bipora (Beddard, 1900) , Metaphire malayana (Beddard, 1900) [syns.? Amyntas pulauensis Beddard, 1900: 904 ;? Amyntas evansi Beddard, 1900: 907 ;? Pheretima baruana Stephenson, 1932 ; Pheretima fovella Gates, 1949: 21 ; and Pheretima strellana Gates, 1949: 34 ] from Malaya, Thailand and Myanmar as described by Stephenson (1932) and Gates (1972: 199), possibly including the Ph. bosschae Horst, 1893 (part) synonym that had genital markings, cf. the types (now = Pheretima darnleiensis )]. Amynthas juliani has genital markings but is similar to Ph. darnleiensis in retaining septum 8/9 whereas Metaphire loriae either has septum 8/9 or lacks it ( Lee, 1981: 554). However, none of these latter species with copulatory pouches is yet known to have the nephridia on spermathecal ducts diagnostic of Pheretima .
Michaelsen (1928: 40, 41) regarded Sumatran Perichaeta atheca Rosa, 1896 as a subspecies of indica [= Pheretima (Pheretima) darnleienis ], although Sims & Easton (1972: 223) listed it as species incertae sedis because its lack of spermathecae prevented its allocation to Pheretima . Another possibly synonymous species is the sexthecal Pheretima poiana Michaelsen, 1913 from Mt Poi (1,100 m altitude), Sarawak, North Borneo that Michaelsen (1913: 89) compared to Ph. philippina (Rosa, 1891) , Metaphire impudens (Michaelsen, 1899) , Metaphire merabahensis ( Beddard & Fedarb, 1895) and Metaphire bindjeyensis (Michaelsen, 1899) . Sims & Easton (1972: 239) have Ph. poiana in a Ph. dubia ( Horst, 1893) species-group that also includes Ph. callosa Gates, 1937 , and Ph. korinchiana Cognetti, 1922 from Korinchi Peak, western Sumatra.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pheretima (Pheretima) darnleiensis ( Fletcher, 1886 )
| Blakemore, Robert J., Csuzdi, Csaba, Ito, Masamichi T. & Kaneko, Nobuhiro 2007 |
Pheretima darleyensis
| Easton 1982: 732 |
Perichaeta martensi javana
| Sims 1972: 260 |
Pheretima (Pheretima) darnleiensis:
| Blakemore 1994: 368 |
| Easton 1984: 119 |
| Easton 1982: 732 |
| Sims 1972: 260 |
| Gates 1937: 320 |
Pheretima floweri:
| Gates 1934: 258 |
Pheretima indica birangi
| Michaelsen 1928: 42 |
Pheretima borneoensis
| Michaelsen 1928: 55 |
| Ude 1925: 108 |
Pheretima decipiens
| Beddard 1912: 180 |
Pheretima benguetensis
| Blakemore 2004: 133 |
| Nakamura 1999: 27 |
| Beddard 1912: 183 |
Amynthas padasensis:
| Beddard 1900: 624 |
Amynthas bosschae:
| Beddard 1900: 625 |
| Beddard 1900: 892 |
Amyntas peregrinus
| Beddard 1900: 644 |
Pheretima belli:
| Michaelsen 1900: 255 |
Pheretima bosschae:
| Michaelsen 1900: 256 |
Pheretima darnleyensis
| Michaelsen 1900: 263 |
Pheretima floweri:
| Michaelsen 1900: 267 |
Pheretima indica
| Gates 1937: 320 |
| Gates 1937: 193 |
| Michaelsen 1935: 108 |
| Gates 1935: 82 |
| Stephenson 1931: 273 |
| Michaelsen 1928: 40 |
| Michaelsen 1900: 275 |
Pheretima martensi:
| Michaelsen 1900: 282 |
Pheretima padasensis typica:
| Michaelsen 1900: 290 |
Pheretima padasensis lokonensis:
| Michaelsen 1900: 290 |
Pheretima padasensis madelinae:
| Michaelsen 1900: 291 |
Pheretima vaillanti:
| Michaelsen 1900: 311 |
Perichaeta padasensis
| Beddard 1895: 73 |
Perichaeta bosschae
| Gates 1972: 199 |
| Horst 1893: 324 |
Perichaeta indica:
| Horst 1885: 4 |
| Horst 1883: 186 |