
Phrynobatrachus discogularis, Pickersgill & Zimkus & Raw, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4350.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:62496B8F-AEB3-4AC8-9C66-6922428E3B2A |
|
DOI |
https://doi.org/10.5281/zenodo.6051106 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD5E57-FFD1-8A67-FF46-FAD0FBB7F9B0 |
|
treatment provided by |
Plazi |
|
scientific name |
Phrynobatrachus discogularis |
| status |
sp. nov. |
Phrynobatrachus discogularis View in CoL sp. nov.
Arthroleptis minutus (non Boulenger) Loveridge, 1933, p 384.
Etymology. Named for its distinctive disc-shaped gular apparatus.
Holotype: MCZ A- 101213 from Dabaga, Tanzania (co-ordinates from MCZ database: 08.116667°S, 35.916667°E), collected by A. Loveridge on 1 January 1930.
Paratypes: MTSN 32618–32620 , three adult males from Gendawaki valley near Bomalang’ombe (08°11’41” S, 35°51’58” E, 1880m) collected by Michele Menegon and Sebastiano Salvidio, February 1999 GoogleMaps ; MCZ A- 101176 – 101177; 101183; 101191; 101193; 101195 – 101196; 101203 – 101204; 101213 from Dabaga (details as per holotype); MCZ A- 150180, MP1671 & LR2893 collected at Dabaga (08°06'06.8"S, 35°54'09.9"E, 1850 m) by M. Pickersgill on 25 November 2014. GoogleMaps
Other material. This material is too poorly preserved or deteriorated to use as paratype material. MCZ A- 17060–17061, 17063, 17065–17066, 17068–17070, 17072, 17074 (Kigogo, approx. 08°38'S, 35°12'E); MCZ A- 101173 – 101175, 101178 – 101182, 101184 – 101190, 101192, 101197 – 101202, 101205 – 101212, 101214 (Dabaga, details as above).
Definition. Characteristic morphological traits such as a tarsal tubercle in addition to inner and outer metatarsal tubercles, pedal webbing and well-developed subarticular tubercles, and the absence of a mid-dorsal dermal ridge, indicate the new species is a member of the genus Phrynobatrachus . This is confirmed by the genetic results.
A small species (length of males 15.8–19.5 mm, mean 17.8 mm, N = 43; females 19.8–21.0 mm, mean 20.5 mm, N = 4). Throat of male immaculate ivory-yellow in life (white in preservative) with a distinctive, flattened, disc-like gular pouch with continuous well-defined lateral and posterior edges. Males quite slender, females rather stocky. Legs relatively short, the tibio-tarsal articulation of adpressed hind limb reaching from the axilla to just anterior of the eye. Horizontal eye diameter a little shorter than snout length. Finger tips rounded, sometimes slightly swollen; toe tips rounded to bulbous, occasionally with weak discs on one or several toes. Toes approximately half webbed, but webbing deeply incised resulting in extensive broad fringing; subarticular tubercles pigmented; tympanum hidden to indistinct and partly obscured by postorbital fold, vertically oval, with a fragmented annulus and diameter approximately 0.25 to 0.33 into horizontal diameter of eye. Internasal distance approximately equal to interorbital distance. At their most extensive, two pairs of elongated scapular warts join to produce a lyre shape. Large femoral gland in males at least 0.5 times the length of thigh. In profile, snout has a convex upper surface. Dorsum with isolated white spinules which may be dense enough to largely obscure both pattern and skin glands. Rarely a continuous light dorsal stripe or line, associated with a transverse pale line from heel to heel. Chest and abdomen creamy white with variable degree of dusky speckling which is concentrated at the base of the forelimbs and laterally towards the flanks. Back of thighs without contrasting dark and light bands. Underside of thighs with some degree of dusky speckling.
Description of holotype ( Fig. 4 View FIGURE 4 ). Body length 18.0 mm, head width 6.0 mm, tibia 8.8 mm, foot length 9.0 mm, femur length 8.5 mm. Tympanum barely visible on one side, largely overlain by postorbital fold, about 0.35X into horizontal diameter of eye. Nostril equidistant between eye and snout. Finger and toe tips slightly swollen, but without definite discs. Webbing of toe I passes proximal tubercle, toe II (inner) with basal webbing but extensively fringed; II (outer) with broad web passing basal tubercle and fringed to beyond distal tubercle; III narrowly webbed to the middle tubercle on both sides; IV (inner) to middle tubercle but fringed to distal tubercle; outer webbing on IV just past proximal tubercle but fringed to middle tubercle; webbing of toe V half way between proximal and distal tubercle with fringe to distal tubercle. Webbing is unpigmented. There are indications of a web flange from the outer metatarsal tubercle along the outer edge of the foot to beyond the proximal subarticular tubercle of toe V. Inner metatarsal tubercle small, oval and pigmented; outer metatarsal tubercle is a much smaller, rounded pimple. The tarsal tubercle is small, flange-like and projecting. The tibiotarsal articulation reaches the eye. A pair of oval warts behind the eyes converge slightly towards a more elongated pair in the scapular region. Head shape from above almost bullet shaped; snout convex in profile. Canthus rostralis rounded, loreal region sloping and slightly concave.
Dorsal markings are difficult to discern. This may largely be due to the naturally sombre dorsal pattern in this species. The upper lip has patchy dark edging apart from an irregular paler area below the tympanum that extends towards the forelimb. A scattering of inconspicuous white spinules along upper labium. There is a dark streak from back edge of the eye to the axilla associated with a postorbital fold, bordering the pale patch dorsally. Above, brown with minute dark speckles and some larger dark spots on the dorsal and scapular warts. A triangular dark patch between the eyes isolates a relatively clear paler triangle on snout. Lower flanks with heavy dark spotting. Tibia with indistinct dark transverse bands that only extend about half the width of the limb (semibars). There is a vague dark rhomboid over the anus but no associated dark banding on the backs of the thighs. Ventrally, off-white in preservative with indistinct blackish marbling at the base of the forelimbs. Thighs are light yellow with a peppering of melanophores posteriorly. The dorsum has scattered, widely-spaced, white-tipped spinules that are smaller and sparser on limbs. There are inconspicuous asperities mentally and on the anterior “gular disc,” but otherwise none ventrally.
Paratypes and other material. The tympanum varies from being almost completely hidden by the postorbital fold to partially distinct, vertically oval and 0.25–0.33X the horizontal diameter of the eye. The head width as a percentage of body length ranges from 30–38% (mean 33%, N = 43); head width/tibia ratio 62–84% (mean 72%, N = 42); tibia/body length 42–55% (mean 49%, N = 45); foot length/body length 41–57% (mean 54%, N = 45); femur length/body length 38–50% (mean 45%, N = 44). Metatarsal tubercles are slightly closer together than to the tarsal tubercle.
There is some variation in webbing, largely due to its interpretation as deeply-incised webbing as opposed to well-developed fringing on the digits ( Fig. 4d View FIGURE 4 ). In general, toe I has basal webbing, although the web fringe may extend almost as far as the last phalanx; toe II has 1–2 phalanges free of broad webbing, but with extensive fringing to or almost to the last phalanx; toe III has 2–2.5 phalanges free, but may be fringed to the last phalanx; toe IV has 3 phalanges free, but with fringing at least to the penultimate phalanx, and toe V has 1–2 phalanges free, but again with extensive fringing. The tips of the fingers vary between tapering and bulbous but lack discs; toe tips are equally variable, and there may be weak swellings on one to three digits.
The most recently collected males have well-developed, isolated dorsal spinules, but these are seldom present in the specimens collected by Loveridge between 1927 and 1930. Although Loveridge’s material is now for the most part soft and discoloured and there is considerable sloughing of the outer skin layers, the holotype is firm and in good condition, and still retains evidence of dorsal asperities.
Colour pattern. (In preservative) The dorsal pattern of preserved material is rather sombre, with few remarkable features. The overall impression is of a brown frog with diffuse darker markings sometimes presenting as a pair of dark spots over the scapulae. A dark post-orbital streak is common. Very few specimens have a welldefined fine light vertebral line to the tip of the snout, generally associated with a light line from heel to heel. There are no distinct dark lateral bands, and the markings on the back of the thighs show little or no contrast. The upper labium lacks distinctive markings although there is often a diffuse lighter area between the back of the eye and the forelimb; the lower labium has a narrow dark margin that may be broken up into dark patches. Ventrally, virtually immaculate to possessing vague isolated ventral spots composed of aggregations of melanophores, predominantly towards the sides or pectorally, seldom continuous across the middle of the venter. The gular disc is typically immaculate, seldom with scattered melanophores that tend to be more concentrated anteriorly. Ventrally, females are immaculate or more commonly with dark blotches, particularly in the pectoral region but sometimes weakly over the entire throat and venter. The underside of the thighs in both sexes is dusted with melanophores. MTSN 32619 has a weak dark band on the back of the thighs.
In life, dorsal markings are more distinct although occasionally obscured by warts and asperities. The dorsal surface of the humerus is generally paler than the dorsum, with a pinkish brown hue. Ventrally almost uniformly whitish or more commonly with dusky brown mottlings pectorally and towards the flanks. Thighs yellowish with a salt-and-pepper effect of dark and light speckles more densely concentrated anteriorly and posteriorly. Throat of male clear bright yellow ( Figs 2 & 3 View FIGURE 3 ).
Sexual dimorphism. Females are slightly larger than males, and noticeably stockier; females lack the secondary sexual characteristics of males such as a gular disc, thenar and femoral swellings. The gular disc in males is usually immaculate; the gular region in females is more frequently blotched.
Genetic differentiation. Phrynobatrachus discogularis sp. nov. was compared with all known Phrynobatrachus species with published 16S rRNA sequence data and was identified within clade B of Zimkus et al. (2010). P. discogularis differed between 4–12% when compared with 15 East African species within clade B ( Table 1). This species was most similar to P. keniensis Barbour & Loveridge, 1928 , P. rungwensis ( Loveridge, 1932) , P. uzungwensis Grandison & Howell, 1983 , and P. cf. uzungwensis , an unidentified species from Tanzania ( Table 1).
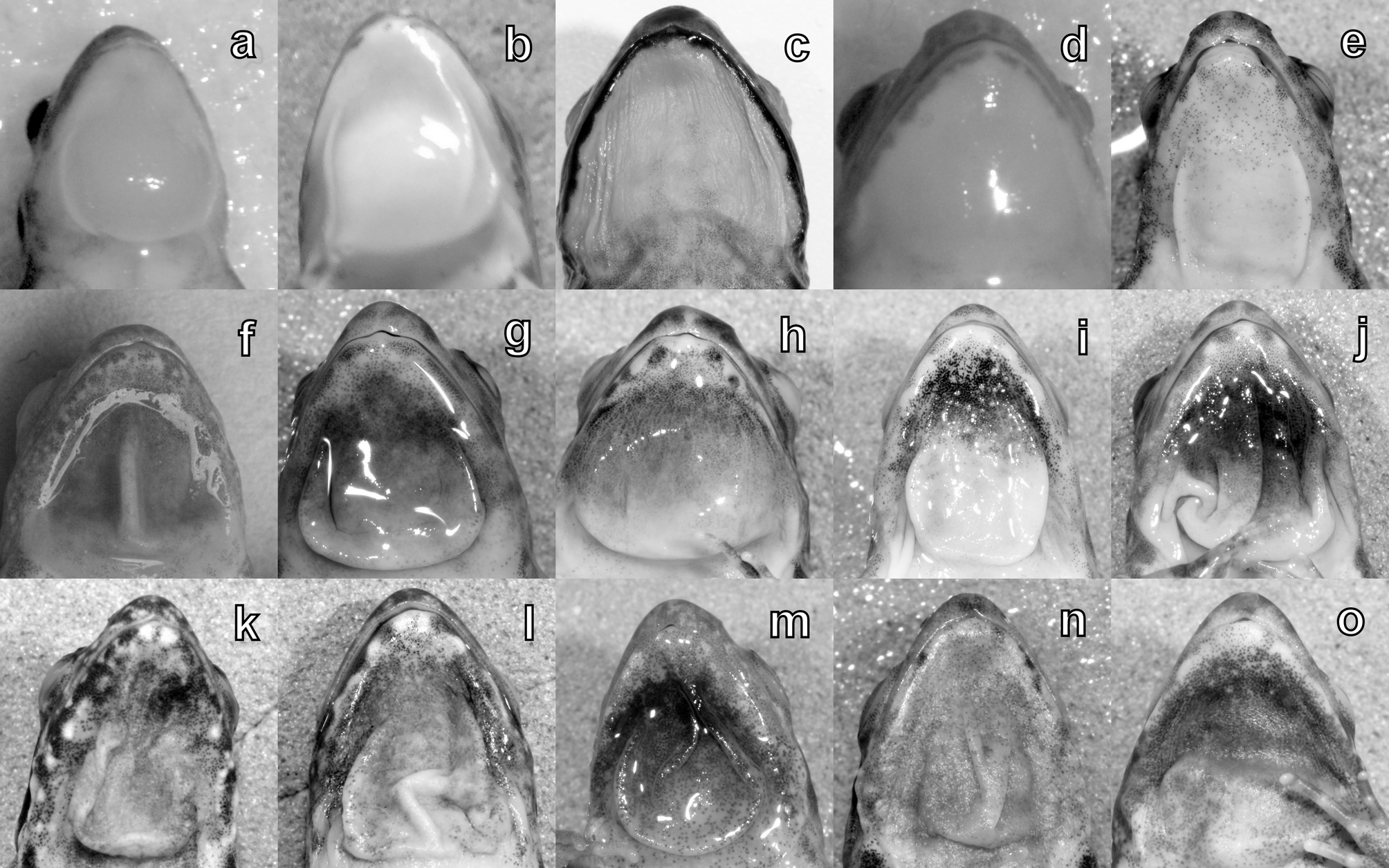
Comparison with other species. Phrynobatrachus discogularis sp. nov. is an unremarkable-looking, rather plain animal. The most distinctive character is the gular apparatus in males: the throat is bright yellow, with lateral gular folds that are well-defined and continuous with a posterior fold, resulting effectively in a rounded or triangular disc most commonly associated with Hyperolius or Afrixalus species. There is little or no bagginess of the gular region itself. While the gular apparatus in many smaller species of Phrynobatrachus may relax into lateral and posterior folds, occasionally resulting in a disc-like gular flap or pouch ( Fig. 5 View FIGURE 5 ), in no other described species has the constancy of this feature been noted apart from in P. gutturosus ( Chabanaud, 1921) from West Africa, and in none of these species is this ‘gular disc’ bright yellow in colour. Phrynobatrachus discogularis further differs from P. gutturosus in its more extensive pedal webbing (rudimentary, not passing proximal subarticular tubercle in P. gutturosus ).
Phrynobatrachus rungwensis View in CoL superficially resembles the new species. Zimkus et al. (2012) found that the webbing of ten P. rungwensis View in CoL specimens ranged from 2.5–3 joints free on the fourth toe; therefore, not unlike P. discogularis View in CoL . The toes may be bulbous to weakly disked, and the gular skin in males forms a posterior flap, resulting in a ‘loose pouch’ ( Schmidt & Inger 1959). Schmidt & Inger originally referred their large series from Upemba National Park to P. gutturosus View in CoL . This material was subsequently identified as P. rungwensis View in CoL by Poynton & Broadley (1985). Unlike discogularis View in CoL , the gular region in males has dark pigmentation (Schmidt & Inger ibid.; Poynton & Broadley ibid.).
Phrynobatrachus breviceps Pickersgill, 2007 View in CoL was described from Mafinga, Tanzania from a single adult male. Although pedal webbing is as extensive as in discogularis View in CoL , the snout is brief, shorter than the horizontal diameter of the eye, while the gular skin is dark and collapses into median and posterior folds.
The yellow gular region distinguishes P. discogularis View in CoL from all eastern African species apart from P. minutus ( Boulenger, 1895) , P. anotis Schmidt & Inger, 1959 View in CoL , P. kreffti Boulenger, 1909 , P. uzungwensis View in CoL , P. kinangopensis Angel, 1924 View in CoL and P. pallidus Pickersgill, 2007 View in CoL .
Phrynobatrachus minutus has tiny dark asperities which are also present ventrally, not prominent and white tipped and confined to the dorsum as in P. discogularis . The gular region in male P. minutus is rarely immaculate - usually with scattered melanophores. Largen (2001) says the gular apparatus sometimes forms of a U-shaped flap with posterior fold and conjoined lateral pleats, but the resultant structure lacks regularity and definition. The webbing is less extensive than in discogularis , reaching to barely passing the proximal subarticular tubercle on toe IV.
Phrynobatrachus anotis closely resembles P. discogularis , but this species lacks even a subdermal tympanum, and the gular skin resolves into a series of lateral pleats without a posterior lip, and the toes have small but distinct discs ( Schmidt & Inger 1959).
Phrynobatrachus kreffti is much larger, with adults ranging in length from 24–39 mm. It possesses large digital discs, and the throat pouch in males collapses into lateral folds ( Fig. 5c View FIGURE 5 ).
Phrynobatrachus uzungwensis is a species from the eastern Udzungwe Mts. The samples used in our comparison perfectly match the type description of that species in all respects ( Grandison & Howell 1983). They are on average larger than discogularis (the type series has a snout –vent range from 16.2–25.0 mm), with wellwebbed toes (two phalanges free on toe IV, the rest webbed to the tip) and prominent digital discs. The snout is somewhat wedge-shaped in profile. The underside of the thigh is yellow with dark spots—not dusted with melanophores giving a salt and pepper effect as in P. discogularis .
Phrynobatrachus kinangopensis resembles uzungwensis in having extensive webbing, and as with that species, males appear to lack an external gular apparatus.
The description of P. albifer (Ahl, 1924) was based on a single, unsexed and possibly immature specimen from Usaramo—now a suburb of Dar es Salaam. Although now in a poor state the holotype ZMB26865 clearly differs from discogularis in its rudimentary pedal webbing, as noted in the original description. Schick et al. (2010) are inclined to regard it as a junior synonym of P. scheffleri ( Nieden, 1910) .
Phrynobatrachus pallidus , also from the coastal region, has much reduced webbing, distinct digital discs, and lateral gular pleats.
Voice. On several occasions at Dabaga recordings were obtained of a short trill or buzz of 0.26– 0.29 s duration and 17–19 pulses per second, followed by a brief click. These were identical to recordings obtained by M. Menegon (personal communication) in the Gendawaki valley near Bomalang’ombe. However, we were unable to verify the species. P. parvulus ( Boulenger, 1905) is syntopic at both localities and has a very similar voice.
Breeding. Unknown. Some of the females collected in January were gravid.
Associated species. Phrynobatrachus uzungwensis has been collected in the surrounding forests, but this species is strictly sylvicolous, while P. discogularis appears to be restricted to open marshy areas. In contrast, P. parvulus is syntopic with discogularis at Dabaga and Bomalang’ombe. Phrynobatrachus natalensis ( Smith, 1849) has been collected at Lusinga, some 10 km from Dabaga (MP3593). The only known specimen of P. breviceps (ZMB66250) was collected at Mafinga, some 70 km from Dabaga and 500 m lower, as was a small series of Phrynobatrachus cf. parvulus (MP4719–4723). Other species recorded from Dabaga and Bomalang’ombe are Amietia tenuoplicata (Pickersgill) , Strongylopus fuelleborni (Nieden) , Ptychadena uzungwensis (Loveridge) , Afrixalus morerei Dubois , Phlyctimantis keithae Schiøtz , Hyperolius pictus Ahl , H. minutissimus Schiøtz and H. substriatus Ahl.
Remarks. Surprisingly, there are very few junior synonyms of central and eastern African Phrynobatrachus species in the literature, and none of these are available for the species described here as new. This is because the names P. minutus and P. mababiensis have long been applied as a wastebasket category to virtually every diminutive Phrynobatrachus from South Africa to the Sudan. Of these smaller species, Phrynobatrachus vanrooyeni Hoffman, 1940 ( South Africa), P. chitialaensis Hoffman, 1944 ( Malawi) , and P. broomi Fitzsimons, 1948 ( South Africa) are considered junior synonyms of P. mababiensis as currently understood ( Frost 2016).
Loveridge (1933) referred his specimens from Dabaga and Kigogo in the Udzungwa Mts. to Arthroleptis minutus (non Boulenger). He specifically described them as having yellow throats and gular discs. This material forms the basis of the type series of P. discogularis sp. nov. as described here. He distinguished this species from his conspecific material with speckled bellies and dark throats, which he correctly identified as Arthroleptis parvulus Boulenger.
Dabaga specimens were common and active during the day in an intensively cultivated valley bottom ( Fig. 6 View FIGURE 6 ). Their preferred habitat is dense vegetation on boggy soil, while the fast-flowing channels artificially created for irrigation are avoided. Loveridge (ibid., p 385) described the habitat at Dabaga as a cattle-trampled bog beside a swiftly flowing brook in the bottom of a valley, and almost certainly refers to the location in the photograph. The site is intensively cultivated and irrigated by a system of channels, but much of the area remains permanently inundated. It is partly bordered by indigenous forest. The location might best be described as “upland meadow”. Menegon (personal communication) described the Gendawaki Valley collection site as ‘pristine grassland about a kilometre from the forest’.
The Udzungwa highlands have long been intensively cultivated and apart from the surviving forests, few pristine habitats remain. Most of these occupy natural drainage areas where the ground is either too steep or too waterlogged to provide viable agricultural land. So far, P. discogularis sp. nov. has tolerated human agricultural practices and the population density is quite high where it occurs. A conservation priority should be to assess the species’ altitudinal and ecological tolerances, its presence and extent of occurrence in the Udzungwa and neighbouring highlands, and the impact of industry and forestry where it occurs.



| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phrynobatrachus discogularis
| Pickersgill, Martin, Zimkus, Breda M. & Raw, Lynn R. G. 2017 |
P. discogularis
| Pickersgill & Zimkus & Raw 2017 |
discogularis
| Pickersgill & Zimkus & Raw 2017 |
discogularis
| Pickersgill & Zimkus & Raw 2017 |
P. discogularis
| Pickersgill & Zimkus & Raw 2017 |
Phrynobatrachus breviceps
| Pickersgill 2007 |
P. pallidus
| Pickersgill 2007 |
P. uzungwensis
| Grandison & Howell 1983 |
P. anotis
| Schmidt & Inger 1959 |
P. kinangopensis
| Angel 1924 |
P. kreffti
| Boulenger 1909 |
P. minutus (
| Boulenger 1895 |