
Psednocnemis, West, Rick C., Nunn, Steven C. & Hogg, Stephen, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.280919 |
|
DOI |
https://doi.org/10.5281/zenodo.6168296 |
|
persistent identifier |
https://treatment.plazi.org/id/DE0287A9-FF9B-556F-FF4A-70E0FBC9479A |
|
treatment provided by |
Plazi |
|
scientific name |
Psednocnemis |
| status |
|
Lyra :
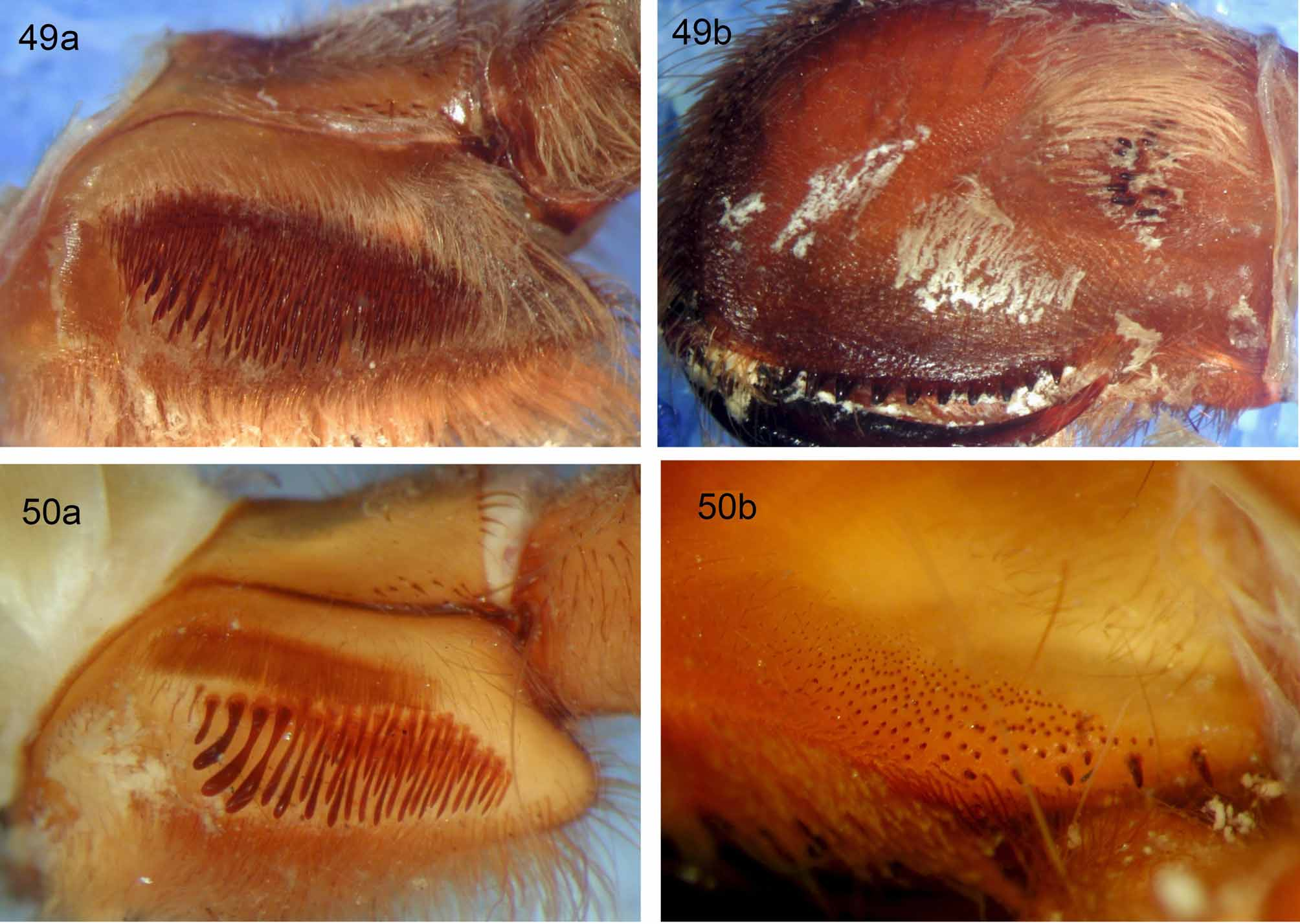
5. Female lyra : (0) rudimentary ( Fig. 47 View FIGURES 46 – 48 ) or absent; (1) oval with proximally truncate form ( Fig. 49a View FIGURES 49 a – 50 b. 49 a, b ); (2) almost circular (Fig. 52a); (3) reinform ( Fig. 51a View FIGURES 51 a – 52 b. 51 a, b ); (4) 1–3 rows of heavy clavates (not reniform) (Fig. 50a). Length = 6, CI = 0.66, RI = 0.86. In those selenocosmiine species that show loss of prolateral maxillary lyra , several issues arise, namely this trait (loss of lyra ) is the singular defining feature of Yamia ( Schmidt & Haupt 2004, Raven 2005, Zhu & Zhang 2008). Yet, many undescribed Orphnaecus species (West & Nunn, pers. obs.) and some Australian species (R. Raven, pers. comm. 2008) also show losses of maxillary lyra . Secondly (and most critical to character state coding), Phlogiellus species show wide ranges of lyrate bacillae counts, but no distinct division between presence/absence (some species such as P. baeri show 1–3 bacillate setae). Therefore, instead of using the ambiguous state “absent”, due to overly subjective character delineation combined with false multi-state character coding ( Sereno 2007), it becomes logical to include the rudimentary condition within the same transformational character state (0). This resolves those subjective character state tendencies, as noted by Jenner (2004), Sereno (2007, 2009), Assis (2009) and Vogt et al. (2010) which can threaten the value of morphological characters. This was so critical to Jenner (2004) and Sereno (2009), they stated that such artificial/weak character state delineation can “ threaten the very future of morphology based cladistics as a vigorous discipline ” (surely a strong enough argument as to support our reasoning). Other genera display unique maxillary lyrate arrangements, Selenocosmia have an almost circular patch of very short bacillae (Fig. 52a), some Orphnaecus possess a reniform patch ( Fig. 51a View FIGURES 51 a – 52 b. 51 a, b ), while Chilobrachys lyra comprises 1–3 rows of heavy claviform bacillae (Fig. 50a).
6. Setal fringe ventrally to prolateral maxillary suture: (0) absent; (1) fine pallid setae ( Figs 49a View FIGURES 49 a – 50 b. 49 a, b , 50a); (2) scopulate, dark setae ( Fig. 47 View FIGURES 46 – 48 ). Length = 3, CI = 0.66, RI = 0.8. The genera Poecilotheria , Haplocosmia and some Chilobrachys possess a setal fringe just ventral to the prolateral maxillary suture, but dorsal to the lyra . In Poecilotheria this fringe is dark, almost scopulate ( Fig. 47 View FIGURES 46 – 48 ), while in Haplocosmia ( Fig. 49a View FIGURES 49 a – 50 b. 49 a, b ) and Chilobrachys (Fig. 50a) the fringe is formed from fine, pallid setae.
7. Tubercles on prolateral maxillary surface: (0) absent; (1) present ( Fig. 47 View FIGURES 46 – 48 ). Length = 1, CI = 1, RI = 0. All Poecilotheria possess 1–12 tubercles immediately distad to patch of rudimentary bacillae on prolateral maxillary surfaces ( Fig. 47 View FIGURES 46 – 48 ). These tubercles are not known in any other selenocosmiine genus.
Sexual morphology (female): Pérez-Miles et al. (1996: characters 13, 14) used spermathecal morphology in their cladistic analysis of Theraphosinae . Guadanucci (2005: characters 11, 12) also used spermathecal morphology in his analysis of Ischnocolinae. Pérez-Miles et al. (2008: characters 8–11) used spermathecal morphology to define their new theraphosine genus Ami Pérez-Miles 2008 ; West et al. (2008: characters 23, 24) used spermathecal morphology in their analysis of Theraphosidae . Five characters were used based on spermathecal morphology, all of which (with the exception of character 8, states 0 and 1 = West et al. 2008: character 23, states 2 and 0) are new.
8. Spermathecae: (0) fused ( Figs 28 View FIGURES 28 – 36. 28 a – b b, 30); (1) not fused ( Fig. 31 View FIGURES 28 – 36. 28 a – b b). Length = 2, CI = 0.5, RI = 0.83. Within Selenocosmiinae examined, both fused ( Smith 2004: fig. 6; 2006: fig. 6) and unfused ( Kulczyński 1908: fig. 1; Schmidt 1999: fig. 5; Haupt & Schmidt 2004: figs 7–9; Zhu & Zhang 2008: figs 4 I, 6 I, 8 I, 9 F; West & Nunn 2010a: figs 6, 23, 51, 70, 101; 2010b: figs 17, 37, 84, 141, 160, 181, 200, 201) spermathecae were noted.
9. Lateral bilobe spermathecal morphology: (0) apically swollen ( Fig. 38 View FIGURES 37 – 45. 37 a – b b); (1) not apically swollen ( Fig. 6 View FIGURES 2 – 7 ). Length = 2, CI = 0.5, RI = 0.75. In all Psednocnemis ( West & Nunn 2010b, figs 37, 64, 141; herein, ( Figs 6 View FIGURES 2 – 7 , 40 View FIGURES 37 – 45. 37 a – b ) as well as L. robustus ( Smith 1988; West 1991; West & Nunn 2010a, fig. 101), lateral bilobes lack apical swelling, differing from all other selenocosmiine species with bilobed spermathecal morphology.
10. Bilobed spermathecae with medial and lateral lobes of similar length ( Fig. 43 View FIGURES 37 – 45. 37 a – b ): (0) absent ( Figs 38 View FIGURES 37 – 45. 37 a – b b, 45); (1) present ( Figs 40, 43, 44 View FIGURES 37 – 45. 37 a – b b). Length = 2, CI = 0.5, RI = 0.91.
11. Spermathecal lobes with mushroom cap ends (mushroomiform): (0) absent; (1) present. Length = 2, CI = 0.5, RI = 0.8. All Selenocosmia within this analysis ( Figs 44 View FIGURES 37 – 45. 37 a – b b, 45), L. fuscus and L. lessunda ( West & Nunn 2010a: figs 51, 70), respectively, herein: ( Fig. 38 View FIGURES 37 – 45. 37 a – b b) possess at least one lobe with mushroom cap (mushroomiform) ends.
12. Spermathecae tombstone-shaped ( Fig. 36 View FIGURES 28 – 36. 28 a – b ): (0) absent; (1) present. Length = 1, CI = 1, RI = 0. This trait was noted in an undescribed Orphnaecus species used in our analysis as well as Orphnaecus philippinus ( Schmidt 1999: fig. 5).
Sexual morphology (male): Male palpal bulb characters are well established synapomorphies of and within Theraphosinae ( Raven 1985, Pérez-Miles et al. 1996, Bertani 2000). Bertani (2000) identified homologous keels within Theraphosinae , we follow him in exploring embolus morphology in Selenocosmiinae , utilizing six characters, herein.
13. Terete embolus lanceolate (LTE) ( Figs 41, 44 View FIGURES 37 – 45. 37 a – b a): (0) absent ( Figs 28 View FIGURES 28 – 36. 28 a – b a, 31a, 32, 34, 35a, 37a, 38a); (1) present ( Figs 39, 41, 44 View FIGURES 37 – 45. 37 a – b a). Length = 1, CI = 1, RI = 0.
14. Single retrolateral keel on embolus (R): (0) absent; (1) strong and heavy ( Figs 34, 35 View FIGURES 28 – 36. 28 a – b a); (2) weak ( Figs 37 View FIGURES 37 – 45. 37 a – b a, 38a). Length = 2, CI = 1, RI = 0. In all Phlogiellus ( Fig. 34 View FIGURES 28 – 36. 28 a – b ) and Orphnaecus ( Fig. 35 View FIGURES 28 – 36. 28 a – b a) examined, a strong and heavy single keel (similar position to R, or retrolateral keel of Bertani 2000) runs along the entire embolus, twisting 90 degrees distally. Lyrognathus (except L. robustus ) also possess a single retrolateral (RR) keel ( West & Nunn 2010a: figs 5, 40 and, herein, Figs 37 View FIGURES 37 – 45. 37 a – b a, 38a), although it is far less pronounced.
15. Basal embolus with distinct heavy retrolateral lobe (BL): (0) absent; (1) present ( Figs 34, 35 View FIGURES 28 – 36. 28 a – b a). Length = 1, CI = 1, RI = 0. In all Phlogiellus ( Zhu & Zhang 2008: figs 8 B, 9 I and, herein, ( Fig. 34 View FIGURES 28 – 36. 28 a – b ) and Orphnaecus ( Fig. 35 View FIGURES 28 – 36. 28 a – b a), retrolateral embolus keel terminates basally as an enlarged lobe (BL). A similar lobe is present in Lyrognathus (RBL) ( Figs 37 View FIGURES 37 – 45. 37 a – b a, 38a), but is far smaller, not so distinct. Additionally, an apophysis is present in Haplocosmia himalayana , however, this structure originates from the tegulum, not from the embolus and thus not applicable to this character.
16. Embolus with distal (A) and sub-distal (SA) keels ( Figs 31 View FIGURES 28 – 36. 28 a – b a, 32): (0) absent; (1) present. Length = 1, CI = 1, RI = 0. All Chilobrachys and Haplocosmia used in this analysis ( Figs 31 View FIGURES 28 – 36. 28 a – b a, 32, respectively) possess both A and SA keels.
17. Distal spiral curl on embolus (AC): (0) absent; (1) present ( Fig. 39 View FIGURES 37 – 45. 37 a – b ). Length = 1, CI = 1, RI = 0. In all known male Psednocnemis (herein, Fig. 39 View FIGURES 37 – 45. 37 a – b ; West & Nunn 2010b: figs 58, 116, 135), the embolus terminates distally with distinct distal “spiral curl” (AC).
18. Embolus with multiple distal keels: (0) present; (1) absent. Length = 1, CI = 1, RI = 0. Among all species used in this analysis, those with more than one keel (or what could be interpreted as a keel) were scored with the state “present”. The outgroup taxon Reichlingia annae ( Reichling 1997: fig. 1) possesses a ventral crest (VC) and prolateral inferior (PI) keel. In Poecilotheria ( Smith 2004: fig. 13, 14; Smith 2006: figs 13, 14; herein, Fig. 28 View FIGURES 28 – 36. 28 a – b a) three keels are distinct, one reverse twisting retrolateral (RCR) keel, as well as prolateral superior (PS) and prolateral inferior (PI) keels, corkscrewing along the entire embolus (in the opposite direction to other selenocosmiine species with embolus keels). Chilobrachys displays prolateral superior (PS) and inferior (PI) keels, sub-distal (SA) and distal (A) keels ( Fig. 31 View FIGURES 28 – 36. 28 a – b a) and in Haplocosmia , male embolus is flattened, twisted, with strong distal (A), sub-distal (SA), and retrolateral (R) keels ( Fig. 32 View FIGURES 28 – 36. 28 a – b ).
Legs:
19. Scopula division on tar. III: (0) divided; (1) undivided. Length = 2, CI = 0.5, RI = 0.89. The conditions “divided/undivided tarsal scopula” have been used extensively throughout history of classification of Mygalomorphae but, herein, is applied only to tar. III. Raven discussed tarsal scopula division in regard to “Anterior/Posterior Gradations” ( Raven 1985); Guadanucci (2005) followed from Raven, both concluded a dependency for tarsal scopula division with most developed scopulation in anterior legs, grading to weaker conditions in hind legs. Furthermore, Guadanucci (2005) concluded tarsal scopula division is not dependant upon size and is useful in phylogenetic analysis (contra Pérez-Miles 1994).
20. Scopula division on met. IV: (0) divided; (1) undivided. Length = 1, CI = 1, RI = 0. West & Nunn (2010b, figs 19, 69, 74), following Pocock (1895a; 1900a) used undivided met. IV scopula to diagnose most Lyrognathus species. Gallon (2003) considered undivided met. IV scopula apomorphic for Eumenophorinae. Guadanucci (2005) concluded, in his work on tarsal scopula division, that dependency of tarsal scopula condition between legs means the more entire scopulate condition, the more apomorphic states it will show. This would also apply to metatarsal scopula.
21. Scopula on met. IV retrolaterally stronger: (0) absent; (1) present. Length = 1, CI = 1, RI = 0. West & Nunn (2010b) first noted the presence of scopula on retrolateral side of division on met IV as almost entire, with prolateral side only extending 1/3 to 2/3 basally in Coremiocnemis (group 1) species ( West & Nunn 2010b: figs 21, 87, 163, 206).
22. Met. IV scopula divided by strong diagonal line: (0) absent; (1) present. Length = 1, CI = 1, RI = 0. Both Coremiocnemis kotacana and the Coremiocnemis sp. from Borneo, used within our data matrix, possess met IV scopula divided and separated distally by strong dividing lines; there is no gradual diminishing of scopula ( West & Nunn 2010b: figs 165, 206).
23. Scopula on tarsi laterally extended: (0) absent ( Figs 29, 35 View FIGURES 28 – 36. 28 a – b c, 38c, 42,); (1) present ( Fig. 46 View FIGURES 46 – 48 ). Length = 1, CI = 1, RI = 0. Following West et al. (2008: character 9, state 3). Raven (1985: table 3, character 17) utilized a similar character in his cladistic analysis of Theraphosidae subfamilies, “ tarsi broader than metatarsi: absent/ present ”, a trait he found synapomorphic for Aviculariinae . This character is noted in all Poecilotheria considered for analyses.
Leg morphology:
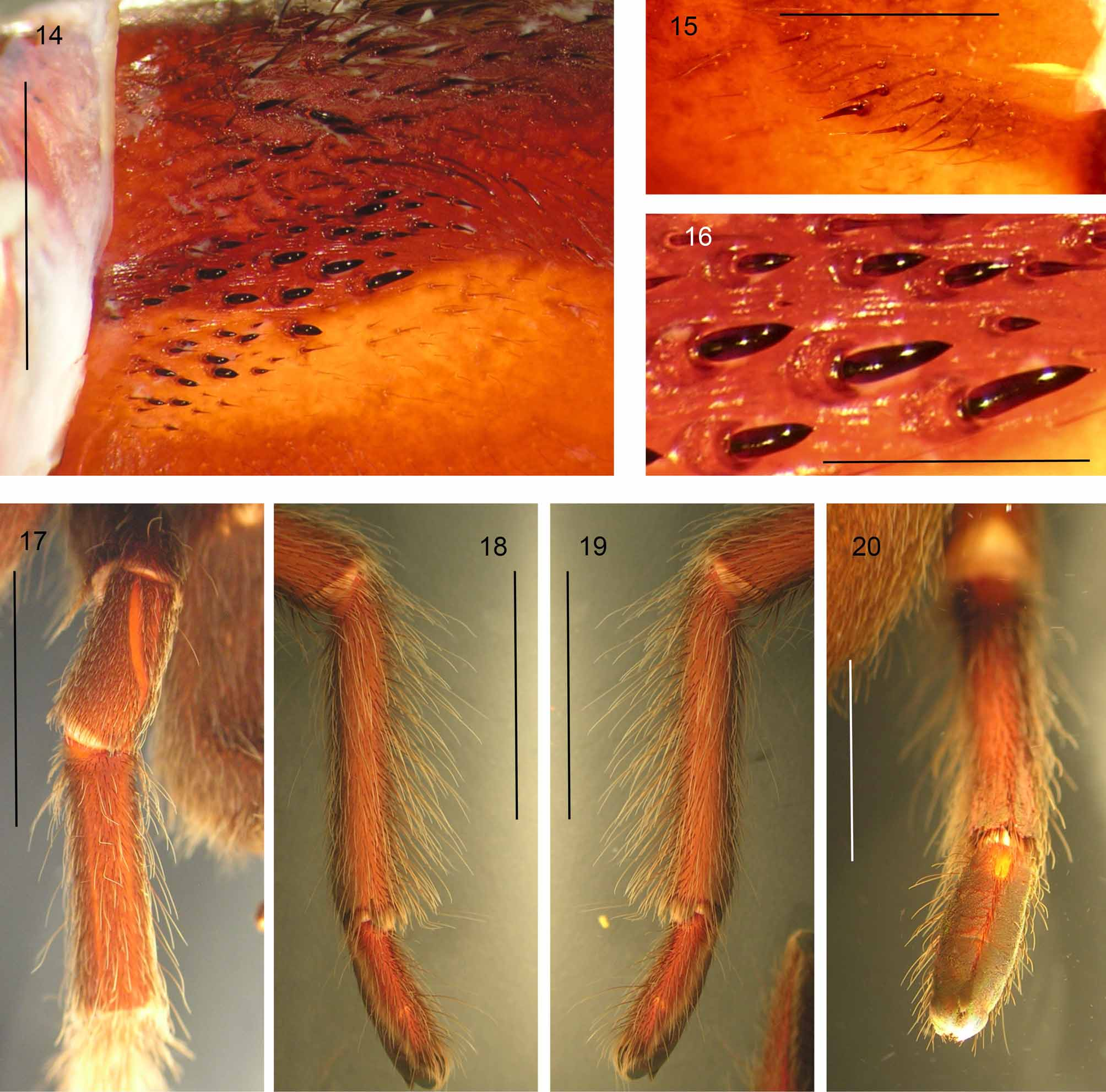
24. Width of Tib. IV in relation to Tib. I: (0) similar to, or smaller than ( Fig. 17 View FIGURES 14 – 20 ); (1) incrassate, much thicker than ( Fig. 38 View FIGURES 37 – 45. 37 a – b c). Length = 1, CI = 1, RI = 0. Pocock (1895a; 1900a) used incrassate tib. IV to diagnose Lyrognathus . Smith (1988), West (1991) and West & Nunn (2010a: fig. 38c) followed Pocock (1895a). Pérez-Miles et al. (1996: character 16) used this character in their analysis of Theraphosinae .
25. Leg spines on tibia: (0) present; (1) absent. Length = 1, CI = 1, RI = 0. Raven (1985) and West et al. (2008: character 11) discussed leg spination in Barychelidae and considered reductions in leg spination to be a derived condition in Theraphosidae .
26. Number of clavate trichobothria on tar. IV: (0) under 25; (1) 25 and over. Length = 2, CI = 0.5, RI = 0.91. Raven (1985: table 1, character 37) considered reduction/loss of tarsal trichobothria a synapomorphy for Atypoidina.
27. Position of clavates on tar. IV: (0) proximal 2/3 to 3/ 4 in straight line; (1) proximal 1/2 or less in straight line; (2) In proximal 1/ 3 in "U" shape. Length = 3, CI = 0.66, RI = 0.91. Raven (1985) discussed variation of leg trichobothria in Mygalomorphae and used those variations in his systematic revision of the infraorder. Among Selenocosmiinae examined, several clavate trichobothrial conditions were noted. Additionally, Guadanucci (in prep.) noted the clavate arrangement in Poecilotheria (Guadanucci, in prep.: fig. 167) as our state (2).
28. Brush of penicillate setae on retrolateral tib. IV ( Fig. 38 View FIGURES 37 – 45. 37 a – b c): (0) absent; (1) present. Length = 1, CI = 1, RI = 0. West & Nunn (2010a) used the presence of penicillate setae along retrolateral surfaces of tib. IV to diagnose all Lyrognathus ( West & Nunn 2010a: figs 8, 20, 53, 71, 98). We introduce this character for analysis for the first time.
29. Brush of recurved setae along retrolateral met. IV ( Fig. 42 View FIGURES 37 – 45. 37 a – b ): (0) absent; (1) present. Length = 1, CI = 1, RI = 0. West & Nunn noted in Coremiocnemis (group 1) the presence of recurved setae in brushform along retrolateral surfaces of met. IV ( West & Nunn 2010b: figs 21, 87, 165, 206). The description of Psednocnemis gen. nov. indicates this characteristic is diagnostic to Coremiocnemis (except C. tropix Raven 2005 ) which is tested here for the first time.
30. Leg width formula IV, III, I, II: (0) absent; (1) present. Length = 2, CI = 0.5, RI = 0.83.
Chelicerae:
31. Intercheliceral setal fringe proximodorsally: (0) absent ( Fig. 14 View FIGURES 14 – 20 ); (1) present ( Fig. 49 View FIGURES 49 a – 50 b. 49 a, b b). Length = 1, CI = 1, RI = 0. In Haplocosmia , a distinct fringe of long pallid setae is present, intermixed with intercheliceral pegs. This fringe has not been seen outside Haplocosmia , and is considered here for the first time.
32. Intercheliceral pegs with prolateral keeled ridge ( Fig. 49 View FIGURES 49 a – 50 b. 49 a, b b): (0) absent; (1) present. Length = 1, CI = 1, RI = 0. In Haplocosmia , and only within that selenocosmiine genus, intercheliceral pegs possess a distinct prolateral keel. As in character 31, this trait is tested for the first time.
33. Spine cluster on retrolateral proximomedial chelicerae ( Fig. 15 View FIGURES 14 – 20 ): (0) absent; (1) present. Length = 2, CI = 0.5, RI = 0.94. West & Nunn first noted retrolateral cheliceral spines in Lyrognathus ( West & Nunn 2010a: figs 26, 60, 80, 96), Coremiocnemis ( West & Nunn 2010b: figs 9, 47, 69, 107, 170, 189, 215) and in Selenocosmia ( West & Nunn 2010a). The same feature is present in Psednocnemis ( Fig. 15 View FIGURES 14 – 20 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.