
Pristimantis ventrigranulosus, Maciel, Natan Medeiros, Vaz-Silva, Wilian, Oliveira, Renan Manoel De & Padial, José Manuel, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.212095 |
|
DOI |
https://doi.org/10.5281/zenodo.5670994 |
|
persistent identifier |
https://treatment.plazi.org/id/530B87B5-FFF1-064D-FF06-FCDF40A10628 |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis ventrigranulosus |
| status |
sp. nov. |
Pristimantis ventrigranulosus sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Holotype. MNRJ 75886, an adult male, from Fazenda Macaúba (16°35’23”S; 51°47’43”W, approximately 400 m above sea level), Piranhas Municipality, State of Goiás, Brazil, collected on 18th October 2009 by Natan M. Maciel, Wilian Vaz-Silva, and Renan M. Oliveira.
Paratypes. MNRJ 75887−89 and ZUFG 5893−95, six adult males, collected along with the holotype. ZUFG 5909, an adult male from the type locality, collected on 26th July 2009 by Renan M. Oliveira.
Generic and species group assignment. To date, there is no identifiable morphological synapomorphy supporting the genus Pristimantis ( Hedges et al. 2008) . We assign the new taxon to the genus Pristimantis based on geography and overall similarity to the majority of species of Pristimantis described. Also, we assign the new species to the P. conspicillatus species group for having Finger I longer than Finger II, belly granular but nor aerolate, a tarsal fold, tympanic membrane distinct, and by its advertisement call composed of single pulsatile notes modulated in amplitude.
Diagnosis. A member of the Pristimantis conspicillatus group, as defined by Lynch & Duellman (1997), characterized by (see also Table 1): (1) dorsal skin coarsely shagreen with some enlarged non-queratinized granules ( Fig. 1 View FIGURE 1 A), larger toward flanks; venter homogeneously granular, granules small ( Fig. 1 View FIGURE 1 C); posterior surfaces of limbs smooth; discoidal fold conspicuous; dorsolateral folds absent; postrictal glands present; (2) tympanic membrane and annulus round, large, their length about 60% eye length ( Fig. 2 View FIGURE 2 B); supratympanic fold short, very prominent ( Fig. 2 View FIGURE 2 B); (3) head slightly longer than wide ( Figs. 1 View FIGURE 1 A–2A); snout acuminate in dorsal view, subacuminate in lateral view ( Figs. 2 View FIGURE 2 A–B); canthus rostralis straight in dorsal view, sharp in profile; (4) cranial crests absent ( Fig. 2 View FIGURE 2 A); upper eyelid covered by small granules ( Figs. 1 View FIGURE 1 A–2A); (5) dentigerous process of prevomers large, situated posteromedial to choanae; (6) males with subgular vocal sac, and one nuptial pad on thumb; (7) hands with long and slender fingers ( Fig. 2 View FIGURE 2 C), Finger 1> Finger 2 ( Fig. 2 View FIGURE 2 C); subarticular tubercles subconical, prominent ( Fig. 2 View FIGURE 2 C); supernumerary tubercles round, smaller than subarticular tubercles ( Fig. 2 View FIGURE 2 C); terminal discs on Fingers I, II and IV round, enlarged on Finger III, ovate to truncate ( Fig. 2 View FIGURE 2 C); circumferential grooves conspicuous, ungueal flap not indented ( Fig. 2 View FIGURE 2 C); lateral fringes and keels on fingers weak ( Fig. 2 View FIGURE 2 C); (8) ulnar tubercles absent; (9) tubercles on heel and tarsus absent, tarsal fold prominent ( Fig. 2 View FIGURE 2 D); (10) inner metatarsal tubercle ovate, prominent, outer subconical, prominent ( Fig. 2 View FIGURE 2 D); supernumerary tubercles absent ( Fig. 2 View FIGURE 2 D); (11) toes long and slender (foot length 50% SVL) ( Fig. 2 View FIGURE 2 D); lateral fringes or keels weak, basal webbing present ( Fig. 2 View FIGURE 2 D); Toe III reaching midpoint of penultimate subarticular tubercle of Toe IV ( Fig. 2 View FIGURE 2 D); tips of toes rounded to ovate, enlarged, ungual flap not indented ( Fig. 2 View FIGURE 2 D); (12) ventral coloration white with fine mottling ( Fig. 1 View FIGURE 1 C); posterior surface of thighs brown without light spots.
Comparisons with similar species. Pristimantis ventrigranulosus is most similar to the east Andean species P. dundeei , P. fenestratus , P. koehleri , P. samaipatae , P. vilarsi , and P. zeuctotylus , and it can be distinguished from them as follows (character states for P. ventrigranulosus in parenthesis). The most similar species, Pristimantis dundeei , has granular belly, but the granules are only posteriorly present (homogeneously granular), double nuptial pad on thumb (single), shagreen dorsal skin (coarsely shagreen), dorsal flat warts (absent), finger fringes absent (finger fringes weakly developed or absent), and prominent toe fringes (weakly developed). Pristimantis fenestratus has smooth ventral skin (granular), shagreen dorsal skin texture (coarsely shagreen), and double nuptial pad on thumb (single). Pristimantis koehleri has granular ventral skin, only posterolaterally (homogeneously granular), finely shagreen dorsal skin texture (coarsely shagreen), finger fringes absent (weakly developed or absent), toe fringes absent (weakly developed), double nuptial pad on thumb (single). Pristimantis samaipatae has smooth belly (granular), finely shagreen dorsal skin (coarsely shagreen), finger fringes absent (weakly developed or absent), toe fringes prominent (weakly developed), ventral coloration immaculate (weakly spotted), double nuptial pad on thumb (single). Pristimantis vilarsi has smooth belly (granular), enlarged ovate discs on Fingers III and IV (round, moderately enlarged), lacks finger fringes (weakly developed or absent), lacks toe fringes (weakly developed). Pristimantis zeuctotylus has a pair of low dorsolateral folds (absent), smooth ventral skin (granular), very enlarged and ovate finger discs on Fingers III and IV (round, moderately enlarged). Two other species of the P. conspicillatus group inhabiting east of the Andes are P. buccinator ( Flores & Rodríguez 1997) and P. skydmainos ( Flores & Rodríguez 1997) but they can be readily distinguished from P. ventrigranulosus by having conspicuous and complete dorsolateral folds. Two other Strabomantids from the Cerrado biome, Oreobates heterodactylus ( Miranda-Ribeiro 1937) , and Pristimantis crepitans ( Bokermann 1965) , can be readily distinguished from P. v e n - trigranulosus by having smooth ventral skin and large and ovate external finger discs, and by lacking tarsal fold and well-developed vocal sac.
Description of holotype. Dorsal skin, including the head ( Fig. 1 View FIGURE 1 A) and hind limbs coarsely shagreen with some enlarged non-queratinized granules, larger toward flanks; skin on sides of head and dorsal forelimbs smooth; gular region ( Fig. 1 View FIGURE 1 B), hind and forelimbs smooth; belly granular, granules small ( Fig. 1 View FIGURE 1 C). Head slightly longer than wide (head length/head width = 1.1) ( Fig. 1 View FIGURE 1 A and 2A). Snout truncate in dorsal view and protruding in lateral view ( Fig. 2 View FIGURE 2 A–B). Nostrils situated and directed posterolaterally ( Fig. 2 View FIGURE 2 B); nostrils visible from front and in lateral views but not in dorsal and ventral views. Canthus rostralis sloped and slightly concave in lateral view; loreal region nearly flat and vertical ( Fig. 2 View FIGURE 2 B). Tongue ovate, premaxillary and maxillary teeth indistinct, dentigerous process of prevomers large and situated posteromedial to choanae, vocal slits not visible. Eye length 16.5 % of SVL, pupil elliptical horizontally ( Fig. 2 View FIGURE 2 B). Tympanic membrane evident, circular, posterodorsally concealed, tympanic annulus well visible ( Fig. 2 View FIGURE 2 B), its diameter 58% smaller than that of EL; supratympanic fold present, prominent and well visible ( Fig. 2 View FIGURE 2 B).
Hands relatively small, length 29% of SVL ( Fig. 2 View FIGURE 2 C). Relative length of adpressed fingers: II <IV <I <III ( Fig. 2 View FIGURE 2 C). Finger I (3.4 mm) longer than Finger 2 (3.0 mm). Discs on Finger I, II and IV moderately expanded; disc on Finger III expanded, bearing lateral fringes ( Fig. 2 View FIGURE 2 C). A large, heart-shaped, outer metacarpal tubercle on median base of palm ( Fig. 2 View FIGURE 2 C); a smaller inner metacarpal tubercle on base of Finger I ( Fig. 2 View FIGURE 2 C); two prominent and well-developed subarticular tubercles on each finger ( Fig. 1 View FIGURE 1 C). Forearm long (84 % longer than arm) and robust (80 % wider than arm). Shank length 56% of SVL. Relative lengths of adpressed toes: I <II <V <III <IV ( Fig. 1 View FIGURE 1 D); inner metatarsal tubercle elliptical and well developed ( Fig. 2 View FIGURE 2 D); outer metatarsal tubercle smaller than inner ( Fig. 2 View FIGURE 2 D); one subarticular tubercle on each toes I and II, two subarticular tubercules on toes III and IV ( Fig. 2 View FIGURE 2 D); three subarticular tubercules on toe IV. Hands lacking lateral fringes and webbing ( Fig. 2 View FIGURE 2 C); feet presenting webbing slightly developed between toes II −III and III −IV ( Fig. 2 View FIGURE 2 D). Tarsal fold present and well developed ( Fig. 2 View FIGURE 2 D).
Measurements of holotype (in mm). SVL 24.8; HL 9.8; HW 4.4; EL 4.1; EN 3.6; IND 2.9; EE 5.5; TYH 2.4; TYL 2.8; F3 1.3; F4 1.1; FA 5.0; TL 13.9; TH 13.0; FL 11.3; T4 1.2.
Coloration. In life, dorsum grayish brown with dark brown markings consisting of oblique bars beginning in the middorsal region toward to the flanks ( Fig. 3 View FIGURE 3 ). Bars also present on dorsal surfaces of the limbs, arms, fingers and toes ( Fig. 3 View FIGURE 3 ). Internal surfaces of the thighs pale brown, not barred. Inguinal region, dorsal surfaces of the thighs and hands (near to articulation radio-humerus) with orange spots. Interorbital and loreal region with dark brown bars between level of eyes and nostrils ( Fig. 3 View FIGURE 3 ). Tip of the snout white ( Fig. 3 View FIGURE 3 ). Labial and subocular dark brown vertical bars ( Fig. 3 View FIGURE 3 ). Supratympanic fold dark brown. Dorsal portion of iris golden and ventral portion dark gold ( Fig. 3 View FIGURE 3 ). Venter cream white with pale brown reticulate spots on ventral surfaces of thighs, gular region and under axillae. The coloration of preserved specimens resembles life coloration, although brown colors fade to gray and orange spots fade to grayish white.
Variation. The finger discs of some individuals of type series are not expanded and lack fringes: MNRJ 75887(finger II and III, left hand); MNRJ 75886 (finger IV, left hand); MNRJ 75889 (finger I, left hand). Subcloacal granules of three individuals (MNRJ 75886, ZUFG 5893 and MNRJ 75889) are absent. The pattern of the dorsal bars is variable in some of the specimens. For variation in morphometrics see Table 2 View TABLE 2 .
cillatus group and the new species described. Data include ranges (first row for each species) and mean ± standard deviation in
parentheses. * Data not available.
Species Notes/call Call length Note length Note rate Pulses Fundamental (ms) (ms) frequency (Hz)
P. dundeei 8 530 50 13.7 * 1808 − 2497
* * * * * *
Advertisement call. Acoustic parameters and comparisons to other species of the P. conspicullatus group are summarized in Table 3. Advertisement calls of Pristimantis ventrigranulosus have no harmonic structure ( Fig. 4 View FIGURE 4 B) and the notes are pulsed, having an average of 6.2 pulses per note ( Fig. 4 View FIGURE 4 A). Calls are composed by four, five, six or seven notes with dominance of five notes (±50%) ( Fig. 4 View FIGURE 4 A). Mean call length is 289 ± 60 ms (range, 202–411 ms) and mean note length is 43.8 ± 8.8 ms (range, 24–74 ms). Mean of the first band energy frequency is 3386.1 ± 313.7 Hz (range, 2127.2–3652.4 Hz) and mean dominant frequency 4021 ± 270.4 Hz (ranging 3451.8–4334.8 Hz). We prefer not to classify the first energy frequency as fundamental frequency because the dominant frequency is not multiple of this first frequency. The maximum intensity is released on the second frequency band, classified as dominant frequency ( Fig. 4 View FIGURE 4 C). In general, the calls had an ascending modulation at the beginning and the last note is the longest, with intermediate notes having similar length ( Fig. 4 View FIGURE 4 A). Amplitude modulation also increases at the beginning of notes and decreases toward the end. Comparisons with similar species show that advertisement call of new species is more similar to P. k o e h l e r i calls, but call length and dominant frequencies distinguish them ( Table 3).
Natural history. Adult males were found calling on bushes, litter and about 60 – 100cm above the ground after 18:30h, on the 18th October 2009. Calling were positioned nearly vertical with their heads directed toward the ground. They were found only in “Palmeiral” vegetation (sensu Ribeiro & Walter 1998) near to the Piranhas River. This kind of phytophysiognomy represents a secondary formation resulted of anthropogenic disturbance of an area of primary forest classified as gallery forest (sensu Ribeiro & Walter 1998). The actual pattern can be characterized as “Palmeiral” formed by the palm tree named “babaçú” ( Attalea speciosa Mart. ), which has a high capacity to adapt to anthropogenic environments (J. D. A. Pereira, pers. com.). Other species found at the type locality and vicinities include: Ameerega berohoka Vaz-Silva & Maciel 2011 ; Barycholos ternetzi ( Miranda-Ribeiro 1937) ; Chiasmocleis albopunctata ( Boettger 1885) ; Dendropsophus cruzi ( Pombal & Bastos 1998) ; D. minutus ( Peters 1872) ; D. nanus ( Boulenger 1889) ; D. rubicundulus ( Reinhardt & Lütken 1862 “1861”); Elachistocleis cesarii ( Miranda-Ribeiro 1920) ; Eupemphix nattereri Steindachner 1863 ; Hypsiboas albopunctatus ( Spix 1824) ; H. lundii ( Burmeister 1856) ; H. paranaiba Carvalho & Giaretta 2010 ; H. raniceps Cope 1862a ; Leptodactylus fuscus ( Schneider 1799) ; L. labyrinthicus ( Spix 1824) ; Leptodactylus aff. latrans ( Steffen 1815) ; L. martinezi ( Bokermann 1956) ; L. mystaceus ( Spix 1824) ; L. podicipinus ( Cope 1862b) ; Lithobates palmipes ( Spix 1824) ; Osteocephalus aff. taurinus Steindachner 1862 ; Phyllomeduza azurea Cope 1862; Physalaemus centralis Bokermann 1962 ; P. cuvieri Fitzinger 1826 ; Pseudopaludicola saltica ( Cope 1887) ; Rhinella mirandaribeiroi ( Gallardo 1965) ; R. ocellata ( Günther 1858) ; R. schneideri ( Werner 1894) ; Scinax fuscomarginatus ( A. Lutz 1925a) ; S. fuscovarius ( A. Lutz 1925b) ; Scinax aff. ruber ( Laurenti 1768) ; and, Trachycephalus typhonius (Laurenti 1758) ( Oliveira et al. 2010; W. Vaz-Silva, unpubl. data;).
Distribution. Pristimantis ventrigranulosus in known only from the type-locality: Fazenda Macaúba, Piranhas Municipality in the state of Goiás ( Fig. 5 View FIGURE 5 ). The region of Piranhas Municipality is associated with the Araguaia- Tocantins River basin, sub-basin of Caiapó River, affluent of right margin of the Araguaia River.
Etymology. The Latin species epithet ventrigranulosus means granular belly, and refers to the feature that most conspicuously distinguishes the new species.
Remarks. Traditionally, the most important character states used to group species within the Pristimantis conspicillatus group (sensu Lynch & Duellman, 1997), were a smooth belly, together with Finger I longer than Finger II. The first to report a slightly granular belly in members of this group were Heyer & Muñoz (1999) for P. dundeei . Nonetheless, P. dundeei , the only other species of Pristimantis inhabiting the dry forests formations of the Cerrado Biome, was recently removed from the P. conspicillatus group by Hedges et al. (2008), probably due to its granular belly. Pristimantis dundeei is now placed on the P. peruvianus group, a group including Andean and Amazonian species sharing a granular belly and a first finger shorter than the second, except in the case of P. peruvianus and P. dundeei . However, the granular texture of P. dundeei differs from the texture of members of the P. peruvianus group, which have a more vascularized ventral skin and larger flat “granules” that is usually referred as an aerolate belly (J. M. Padial, pers. obs.). In addition, the specimen identified by Hedges et al. (2008) as P. peruvianus , and included in their molecular phylogeny corresponds to P. re i c h l e i (Padial & De la Riva 2009), a species with the first finger shorter than the second and with aerolate belly. Moreover, the advertisement call of P. dundeei is identical in structure to the call of other members of the P. conspicillatus group, and characters of advertisement calls of this group have been shown to content informative phylogenetic information ( Goicoechea et al. 2010). Therefore, given the doubtful status of the P. peruvianus group, and the shared call structure and most morphological characters between P. dundeei and other members of the P. conspicillatus group, including P. ventrigranulous , we reassign P. dundeei to the P. conspicillatus group.

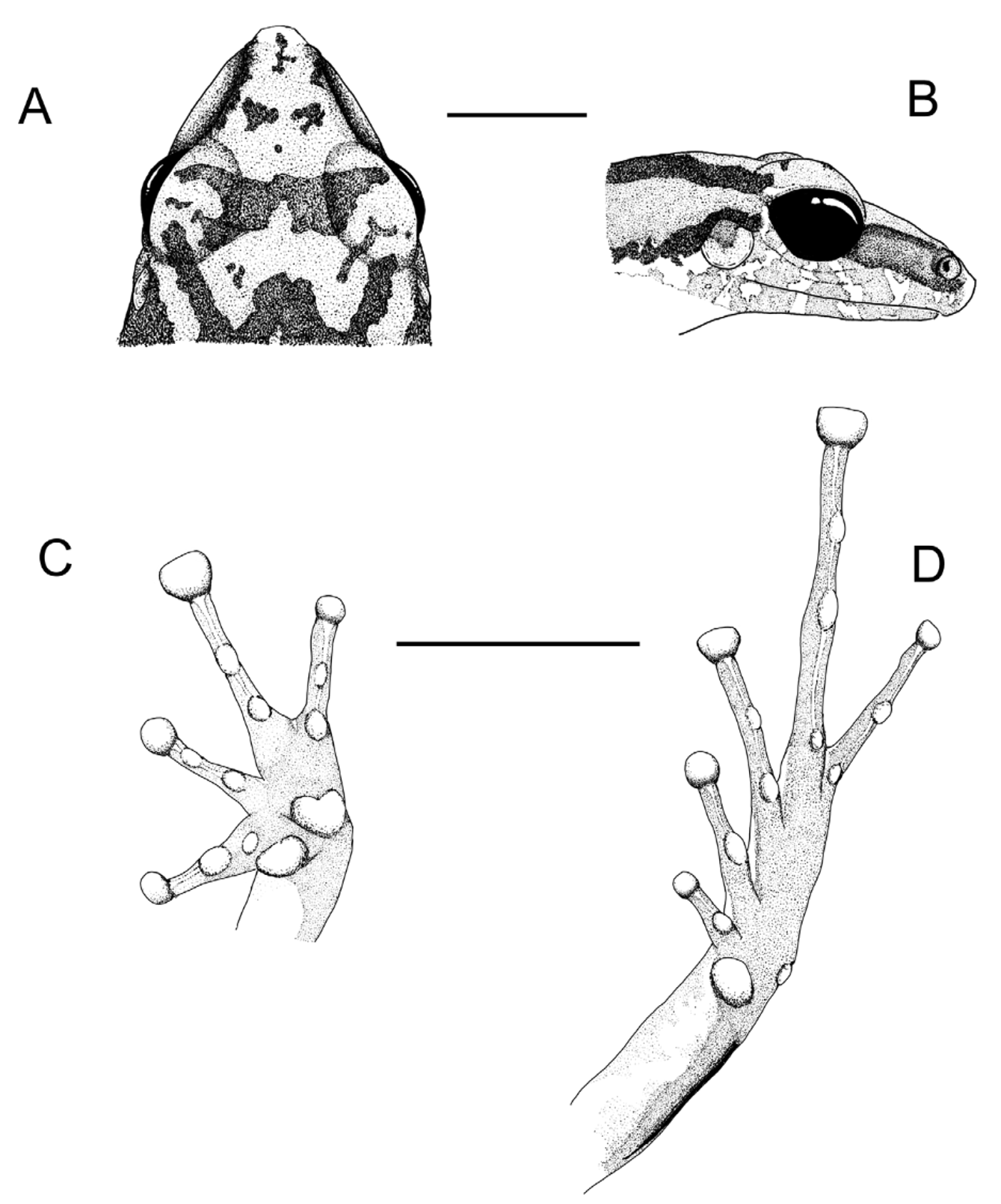


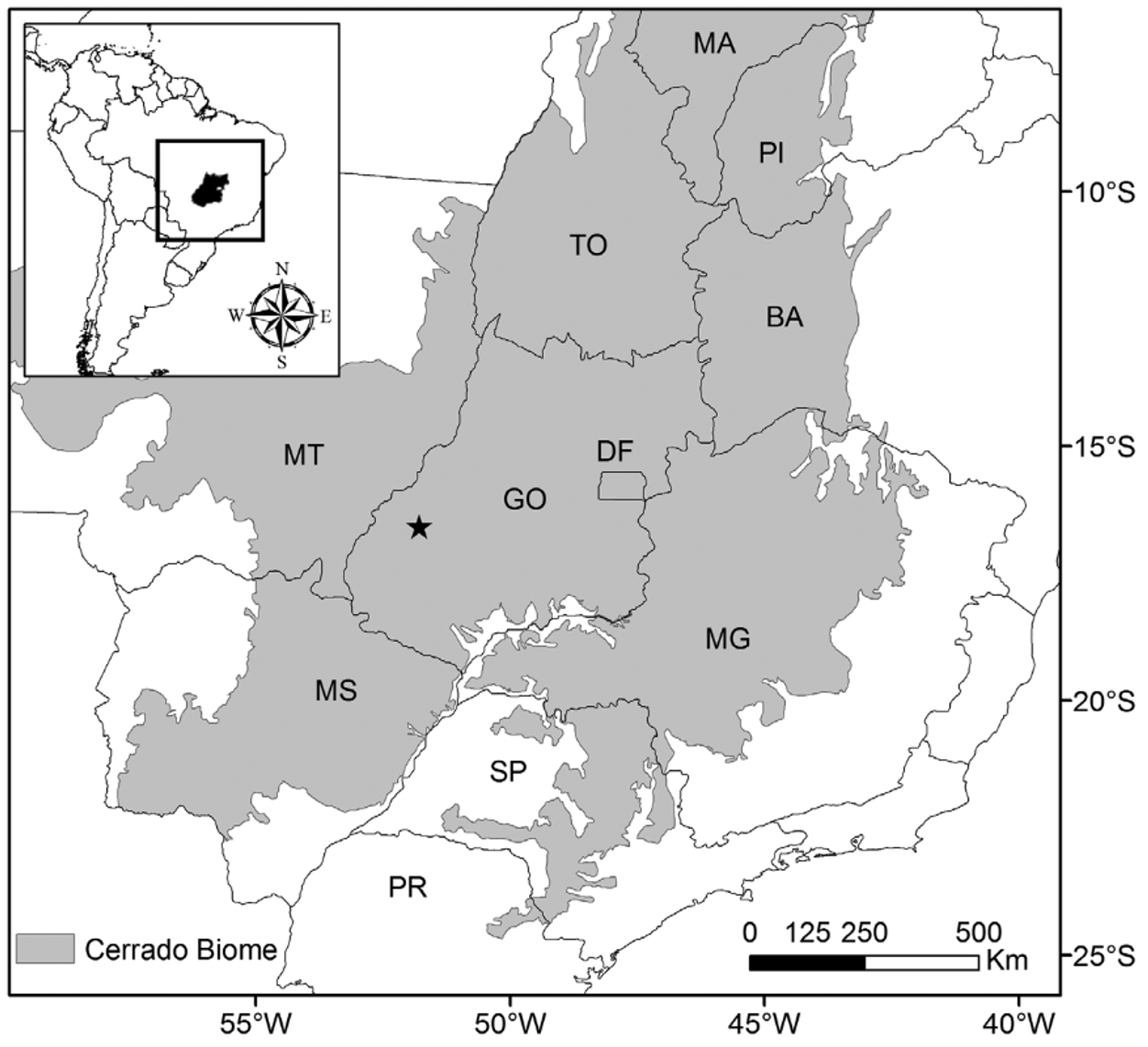
TABLE 2. Means, Standard deviation (SD), and ranges (in mm) of type series (eight males) of Pristimantis ventrigranulosus. Character Mean (± SD) Range
| SVL | 23.5 (± 1.9) | 20.2 – 25.4 |
|---|---|---|
| HL | 9.5 (± 1.0) | 8.2 – 11.4 |
| HW | 8.4 (± 0.9) | 6.9 – 9.3 |
| EL | 3.6 (± 0.4) | 3.1 – 4.2 |
| EM | 3.3 (± 0.4) | 2.6 – 3.7 |
| IND | 2.7 (± 0.1) | 2.5 – 2.9 |
| EE | 5.3 (± 0.4) | 4.8 – 5.9 |
| TYH | 2.2 (± 0.3) | 1.7 – 2.4 |
| TYL | 2.4 (± 0.3) | 1.9 – 2.8 |
| F3 | 1.4 (± 0.2) | 1.1 – 1.6 |
| F4 | 1.1 (± 0.1) | 1.0 – 1.4 |
| FA | 4.7 (± 0.8) | 3.5 – 5.9 |
| TL | 13.8 (± 1.7) | 10.0 – 15.6 |
| TH | 13.2 (± 1.3) | 10.8 – 15.0 |
| FL | 12.4 (± 1.5) | 9.6 – 14.2 |
| T4 | 1.2 (± 0.2) | 1.0 – 1.5 |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.