
Pugettia quadridens (De Haan, 1837), 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4672.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:01B7FFA9-1EC8-41A7-8DDE-418D9F4882B8 |
|
persistent identifier |
https://treatment.plazi.org/id/FF0187EC-FF91-CE5F-EAE7-F895FAF5F7A7 |
|
treatment provided by |
Plazi |
|
scientific name |
Pugettia quadridens (De Haan, 1837) |
| status |
sensu stricto |
Pugettia quadridens (De Haan, 1837) View in CoL sensu stricto
[Japanese name: Yotsuha-mo-gani]
( Figs. 2–13 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 )
Pisa (Halimus) quadridens De Haan, 1837 : pl. 24, fig. 2 [type locality: Japan].
Pisa (Menoethius) [sic.] quadridens View in CoL .—De Haan 1839: 97–98 (in part).— Yamaguchi & Baba 1993: 353, fig. 113 (in part); 2003: 38 (in part).
Menaethius quadridens .— Adams & White 1849: 20.
Pugettia quadridens View in CoL .— Sakai 1936: 88 (in part), pl. 20, fig. 2, text-fig. 37; 1965: 73 (in part?), pl. 32, figs. 4, 5.— Yamaguchi et al. 1976: 35; 1987: 13, pl. 4, fig. 3a, b.— Ikeda 1981: 15.— Takeda 1982: 120, 1 unnumbered figure.— Griffin & Tranter 1986: 97 (in part).— Yamaguchi & Baba 1993: 353, fig. 113 (in part); 2003, 38 (in part).— Muraoka 1998: 24 (in part).— Minemizu 2000: 209, 1 unnumbered figure; 2002: 209, 1 unnumbered figure.— Marumura & Kosaka 2003: 32 (in part).— Ikeda & Kuramochi 2004: 12, 2 unnumbered figures.— Takeda et al. 2006: 196 (list); 2011: 49, fig. 16-69.— Ng et al. 2008: 101 (list).— Yamaguchi & Henmi 2008: 80, figs. 1a, 2a–c, 3a–c.— Nunomura 2010: 52 (in part).— Wicksten & Stachowicz 2013: 359 (list).— Watanabe 2014: 41, 1 unnumbered figure.— Yoshizaki 2018: 30 (list), 48 (with 11 unnumbered figures), 173 (1 unnumbered figure of frontal region), 181 (1 unnumbered figure of left chela).
Pugetti [sic.] quadridens View in CoL .— Sakai 1938: 255–257 (in part), text-fig. 28a, pl. 26, fig. 1.
Pugettia quadridens quadridens View in CoL .— Sakai 1976: 196–197 (in part), text-fig. 103a, pl. 68, fig. 1.— Miyake 1983: 206 (list); 1998: 206 (list).— Wada 1995: 387, pl. 103, fig. 5.
Pugettia nipponensis View in CoL .— Miyake 1983: 36, pl. 12, fig. 5; 1998: 36, pl. 12, fig. 5. [not Pugettia nipponensis Rathbun, 1932 View in CoL ]
Pugettia quadridens pellucens View in CoL .— Marumura & Kosaka 2003: 32 (in part). [not Pugettia quadridens pellucens Rathbun, 1932 View in CoL ]
? Menaethius guadridens [sic.].—Matsuura 1895: 23.
? Pugettia quadridens View in CoL .— Stimpson 1857: 219; 1907: 24–25.— Miers 1879: 23 (in part).— Ortmann 1893: 43 (in part?).— Rathbun 1894: 71–72 (in part?); 1902: 28 (in part).— Doflein 1902: 655–656.— Terazaki 1903: 15–16, unnumbered figure.— Parisi 1915: 285–286.— Balss 1924: 24 (in part).— Urita 1926: 32.—Kikuchi 1931: 19.— Sakai 1934: 294.— Kamita 1935: 63.— Kim et al. 1979: 110.— Kim & Kim 1982: 146.— Kim & Chang 1985: 45.— Komai 1999: 86.
? Pugettia quadridens quadridens View in CoL .— Kim 1985: 79–80.— Kim & Kim 1986: 325.— Ito & Honma 2001: 27
Material examined. Lectotype: male ( CL 24.2 mm) ( RMNH D 42298), Japan, coll. H. Bürger, 1825–1834.
Paralectotypes: 4 females ( CL 16.9–23.6 mm) ( RMNH D 42298), same data as lectotype .
Non-types: Japan. One female (18.9 × 15.1 mm) ( CBM-ZC 14872), near low tidal mark, Ahnfeltia paradoxa turf, Misaki, Iwaki, Fukushima, hand collection, coll. N. Ohtsuchi, 2 Feb. 2017; 1 male (18.3 × 14.1 mm), 1 ovigerous female (17.0 × 13.3 mm) ( CBM-ZC 14873 ), Gelidium elegans turfs, near low tidal mark, Oarai, Kashima Sea, hand collection, coll. N. Ohtsuchi & S. Houki, 8 May 2012 ; 2 males (18.9 × 15.0, 19.6 × 15.0 mm), 2 ovigerous females (17.4 × 13.5, 18.0 × 15.3 mm), 1 female (with a rhizocephalan parasite, 17.6 × 13.6 mm) ( CBM-ZC 14874 ) , 1 male (19.4 × 15.1 mm), 1 ovigerous female (21.0 × 16.6 mm) (NSMT-Cr 26063), A. paradoxa turfs, intertidal, Inubosaki, Chiba, hand collection, coll. N. Ohtsuchi & S. Houki, 9 May 2012 ; 1 male (18.5 × 14.2 mm) (NSMT-Cr 26064), intertidal, Futomi, Kamogawa, Chiba, Boso Peninsula, hand collection, coll. N. Ohtsuchi, 9 Apr. 2005 ; 1 female (14.3 × 10.4 mm) ( TOYA Cr-20553), Shirahama, Shirahama-machi, Chiba, coll. N. Nunomura, 18 Apr. 1995 ; 1 male (19.5 × 15.3 mm) ( CBM-ZC 7581 ), intertidal, Okinosima, Tateyama, Chiba, Boso Peninsula, coll. Y. Matsuzawa, Aug. 2003 (exmined in Ohtsuchi et al. 2014) ; 3 males (13.9 × 10.4–19.1 × 14.7 mm), 2 ovigerous females (15.4 × 11.1, 18.0 × 13.5 mm) (NSMT-Cr 7561), Kohyatsu, Tateyama, Chiba, Boso Peninsula, coll. student of Ochanomizu Univ. (examined in Ohtsuchi et al. 2014) ; 4 males (19.7 × 15.4–22.9 × 18.0 mm), 2 females (20.5 × 15.9, 22.2 × 17.5 mm), 2 ovigerous females (20.7 × 15.9, 21.9 × 17.4 mm) (OMNH-Ar 10699), 5 males (14.5 × 11.5–20.3 × 15.9 mm), 3 females (17.6 × 14.1–20.1 × 16.6 mm), 1 ovigerous female (20.6 × 16.2 mm) ( CBM-ZC 14875 ), Grateloupia cornea turfs, near low tidal mark, Tsurugizaki, Miura Peninsula, hand collection, coll. N. Ohtsuchi, J. Hayakawa & S. Houki, 26 Feb. 2012 ; 3 males (6.9 × 4.7–8.9 × 6.2 mm) ( CBM-ZC 14876 ), 2–4 m, Ge. elegans turfs, Nagai, Yokosuka, Miura Peninsula, SCUBA +air-lifting sampler, coll. J. Hayakawa & T. Onitsuka, 10 Oct. 2008 ; 1 female (10.5 × 7.3 mm) ( CBM-ZC 14877 ), 2–4 m, Marginisporum crassissimum turfs, same locality as previous, SCUBA +air-lifting sampler, coll. N. Ohtsuchi & J. Hayakawa, 5 Apr. 2010 ; 1 female (11.2 × 8.1 mm) (NSMT-Cr 26065), Sargassum fusiforme beds, near low tidal mark, Nagai, Yokosuka, Miura Peninsula, hand collection, coll. N. Ohtsuchi & S. Houki, 24 Oct. 2010; 1 female (16.7 × 12.8 mm) (RUMF-ZC-4338), same locality and habitat as previous, hand collection, coll. N, Ohtsuchi, 19 Apr. 2016 ; 1 male (16.7 × 12.8 mm) (RUMF-ZC- 4974), same locality and habitat as previous, hand collection, coll. N. Ohtsuchi, 11 Mar. 2017 ; 1 male (14.9 × 10.2 mm) (NSMT-Cr 16481), 6 m, Hayama Beach, Miura Peninsula, Sagami Bay , coll. I. Soyama, 27 Jan. 2005 (examined in Ohtsuchi et al. 2014) ; 2 males (21.6 × 17.6, 23.8 × 18.6 mm) (NSMT-Cr 26061), intertidal, S. fusiforme beds, rocky beach behind Morito Shrine, Hayama, Miura Peninsula, hand collection, coll. N. Ohtsuchi, 11 Apr. 2009; 1 male (27.3 × 22.4 mm) (NSMT-Cr 26059), 1 ovigerous female (21.8 × 18.2 mm) (NSMT-Cr 26060), 3 males (16.7 × 13.1–27.4 × 23.3 mm), 1 ovigerous female (21.1 × 16.0 mm) (NSMT-Cr 26062), same habitat and locality as previous, hand collection, coll. N. Ohtsuchi, 29 Apr. 2010 ; 1 male (12.1 × 8.9 mm) ( KPM-NH 104997 ), Hayamacho, Miura-gun ; 1 male (21.0 × 17.3 mm) (WMNH-Na-Cr 0312), Oiso, Kanagawa, Sagami Bay , coll. S. Nagai, Sep. 1970 ; 1 males (13.6 × 10.2 mm), 1 female (with rhizocephalan parasite, 13.3 × 10.2 mm) ( KPM-NH 104582 ), Manazuru-machi, Ashigara-shimogun, 1962 ; 6 males (10.1 × 7.2–27.4 × 21.7 mm), 1 female (16.9 × 12.3 mm), 3 ovigerous females (17.1 × 13.4–19.4 × 14.9 mm) ( KPM-NH 104045 ) , 1 male (19.3 × 15.7 mm) ( KPM-NH 104188 ), Sagami Bay ; 1 ovigerous female (17.7 × 12.6 mm) (NSMT-Cr 6062), Izu Ohshima I., coll. I. Soyama, 6 May 1979 ; 5 males (4.3 × 2.9–6.2 × 4.4 mm), 3 females (4.2 × 2.9–6.2 × 4.5 mm), 4 juveniles (NSMT-Cr 26066), near low tidal mark, Ge. elegans turfs, Hinodehama, Izu Ohshima I., hand collection, coll. N. Ohtsuchi, 10 Aug. 2006; 1 male (25.9 × 19.1 mm) (NSMT-Cr 11306), Himaga I., Mikawa Bay , Mie, coll. T. Nishikawa, 7 Apr. 1993 ; 2 males (22.1 × 17.0, 23.2 × 18.3 mm) ( KPM-NH 104513 ), Wagu, Shima, Shima, Mie ; 1 male (20.6 × 16.0 mm) (OMNH-Ar 9916), Wagu, Tousi-cho, Toba-shi, Mie, coll. T. Yamamoto, 11 Apr. 1982 ; Three males (20.3 × 17.0–26.8 × 21.7 mm) ( KPM-NH 104476 ), Kii Nagashima, Kitamuro-gun, Mie ; 1 male (10.7 × 7.4 mm) (NSMT-Cr 11645), Kushimoto, Kii Peninsula, Wakayama, coll. I. Soyama, 21 Jun. 1994 ; 1 ovigerous females (17.7 × 13.2 mm), 1 females (with rhizocephalan parasite, 18.7× 14.2 mm) (NSMT-Cr 4894), Shirahama, Kii Peninsula, Wakayama, coll. Y. Koyama, 1 Apr. 1972 ; 2 males (3.2 × 4.6, 6.0 × 4.3 mm), 1 female (5.2 × 3.8 mm), 3 juveniles (NSMT-Cr 26067), Ge. elegans turfs, 1 m, Engetsu I., Banshozaki, Seto, Shirahama, Wakayama, snorkeling+hand collection, coll. N. Ohtsuchi, 28 Aug. 2015 ; 2 males (20.7 × 16.0, 21.8 × 17.1 mm) (SMBL-Art 1079), Seto, Shirahama-cho, Nishimuro-gun, Wakayama, 15 Apr. 1936 ; 2 males (15.3 × 11.0, 17.3 × 13.2 mm), 2 females (18.3 × 14.4, 15.2 × 11.9 mm) (SMBL-Art 1094), same locality as previous, 18 Feb.1939 ; 5 males (19.0 × 15.2–16.3 × 12.6 mm), 2 ovigerous females (19.3 × 14.2, 14.2 × 10.8 mm), 2 females (with a rhizocephalan parasite, 16.9 × 13.2, 16.5 × 12.9 mm) (SMBL-Art 1091), Ezura, Shirahama-cho, Nishimuro-gun, Wakayama, Jul. 1939 ; 1 male (24.9 × 19.8 mm) ( KPM-NH 104081 ) , 1 male (25.0 × 19.7 mm) ( KPM-NH 104364 ) , 4 males (22.8 × 18.2–28.4 × 22.4 mm) ( KPM-NH 104378 ) , 2 males (23.6 × 19.1, 26.2 × 20.7 mm) ( KPM-NH 104448 ) , 1 female (with rhizocephalan parasite, 10.9 × 8.2 mm) ( KPM-NH 104614 ) , 1 female (25.0 × 14.6 mm) ( KPM-NH 104957 ), Minabe, Hidaka-gun, Wakayama ; 2 females (with rhizocephalan parasite, 14.0 × 10.0, 14.1 × 10.6 mm) ( KPM-NH 104624 ), Sakai, Minabe, Hidaka-gun, Wakayama ; 1 male (13.1 × 9.7 mm) (OMNH-Ar 6644), Kuroshima, Yura-cho, Hidakagun, Wakayama, 3 May 1984 ; 2 females (10.4 × 7.7, 9.6 × 7.7 mm) (OMNH-Ar 6576), Nishihiro, Hirokawa-cho, Arida-gun, Wakayama, coll. T. Yamashita, 19 Aug. 2000 ; 1 ovigerous female (15.7 × 12.2 mm) (OMNH-Ar 2392), Tagurazaki, Kada, Wakayama-shi, 18 May 1980.— 6 males (13.9 × 10.2–22.5 × 18.4 mm) (OMNH-Ar 9913), same locality as previous, coll. Fukui, 24 Mar. 1974 ; 2 ovigerous females (15.6 × 12.2, 12.7 × 10.0 mm) (OMNH-Ar 3932), Minami-Tarumi, Tomogashima, Kada, Wakayama-shi, 26 Apr. 1997; 1 male (27.1 × 22.3 mm) (OMNH-Ar 524), 1 male (23.1 × 17.9 mm) (OMNH-Ar 525), Jogasaki, Kada, Wakayama, coll. Y. Fukui & N. Nunomura, 19 May 1974 ; 1 ovigerous female (19.6× 15.8 mm) (OMNH-Ar 2384), Okawa-touge-shita, Okawa, Wakayama-shi, 13 Jul. 1980; 1 ovigerous female (18.6 × 14.4 mm) (OMNH-Ar 6212), Myojinzaki, Misaki-cho, Osaka, coll. H. Ariyama, 29 Apr. 1994 ; 1 male (18.8 × 14.0 mm), 1 female (12.2 × 9.4 mm) (OMNH-Ar 2448), bedrock, Toyokunizaki, Misaki-cho, Sennan-gun, 17 Feb. 1980; 2 males (18.0 × 13.7, 20.8 × 16.6 mm) (OMNH-Ar 9911), Nagasaki, Misaki-cho, Sennangun, Osaka, coll. R. Yamanishi, 11 May 1978 ; 4 males (23.5 × 18.2–15.3 × 11.6 mm), 2 ovigerous females (15.5 × 11.9, 18.6 × 14.5 mm) (OMNH-Ar 6024), Jogasaki (?), Misaki-cho, Osaka, coll. H. Ariyama, 16 Apr. 1995 ; 1 male (16.5 × 12.7 mm) (OMNH-Ar 2779), Yamatojima, Iwaya, Awaji-cho, Tsuna-gun, Hyogo, 8 May 1982 ; 1 ovigerous female (15.7 × 12.7 mm) (OMNH-Ar 5953), off Shioya, Tsuna-cho, Tsuna-gun, Hyogo, 7.8 m, coll. R. Yamanishi, 9 May 1985 ; 1 male (20.6 × 16.0 mm), 1 ovigerous female (16.4 × 12.9 mm) (OMNH-Ar 9917), Takino-chaya, Tarumi, Kobe, coll. Y. Shibata, 13 Mar. 1960; 1 male (19.7 × 14.4 mm) ( CBM-ZC 8516 ), ca. 5 m, Kurahashi I., Hiroshima, Seto Inland Sea, commercial trawler, coll. K. Kuramoto, 13 Apr. 2005 ; 1 male (18.5 × 14.7 mm) (OMNH-Ar 6500), Nakamichi, Aio-cho, Yoshiki-gun, Yamaguchi, coll. T. Watanabe, 4 May 2003 ; 1 female (with rhizocephalan parasite, 15.2 × 11.8 mm) (OMNH-Ar 6512), same locality and date as previous, coll. K. Hatooka ; 1 female (16.7 × 12.4 mm) (KPM- NH 104006), Matsuyama, Ehime, 1967 ; 1 male (32.1 × 26.0 mm) ( KPM-NH 104903 ), Tosa Bay ; 1 male (13.3 × 9.4 mm) ( TOYA Cr-3230) , 1 female (8.4 × 6.2 mm) ( TOYA Cr-3231), Yokata, Toyama-shi, Toyama, coll. N. Nunomura, 8 Sep. 1982 ; 1 male (11.7 × 9.0 mm) (OMNH-Ar 9919), Shibagaki Kaigan, Hakui-shi, Ishikawa, 5 Aug. 1974 ; 1 male (24.1 × 19.2 mm) (TRPM-AAr-0000498), off Tottori, 4 Aug. 1998 (photographed in Takeda et al. 2011) ; 3 males (11.9 × 8.5–18.9 × 14.3 mm), 1 male (with rhizocephalan parasite, 12.4 × 9.9 mm) (NSMT-Cr 6771), 1 male (10.0 × 7.2 mm), 1 female (11.8 × 8.5 mm) (NSMT-Cr 6772), Tsuji I., Amakusa Archipelago, Kumamoto, coll. Y. Fukuda, 18 Mar. 1980 ; 1 male (19.5 × 14.2 mm) ( KMNH IvR 100009), Shiroiwazaki, Tomioka, Amakusa, Kumamoto, Amakusa Sea, coll. Baba, 6 Apr. 1933 ; 1 ovigerous female (14.4 × 10.7 mm) ( KMNH IvR 100010), Nomozaki, Nagasaki, Nagasaki Peninsula, Amakusa Sea, setnet around the rocky reaf, coll. K. Matsubayashi ; 1 male (15.5 × 11.2 mm) (ZMUC-CRU- 20233), Nagasaki, coll. James Jordan (examined in Griffin & Tranter 1986) ; 1 male (13.0 × 9.1 mm) ( KMNH IvR 100012), 1 ovigerous female (14.2 × 11.0 mm) ( KMNH IvR 100013), 1 ovigerous female (17.5 × 13.3 mm) ( KMNH IvR 100014), Tsuyazaki, Fukutsu, Fukuoka, Genkai Sea, coll. Sakai, 23 Jun. 1960 ; 1 male (14.2 × 10.0 mm) (WMNH- Na-Cr 0312-2), lower tidal line, Kume-jima I., Okinawa Prefecture, Ryukyu Islands, southern Japan , coll. S. Nagai, 18 Nov. 1992 .
China. One male (16.2 × 12.6 mm) ( MBM 160503), off east Ping Tan I., Fuzhou, Fujian, coll. Fan Xu, 18 Mar. 1957 .
Redescription. Male. Full-grown males (15.3–32.1 mm PCL, including lectotype). Carapace ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ) pyriform, 1.2–1.4 longer than width (PCL/CW = 1.3±0.0, N = 52), surface smooth to naked eyes but closely covered with microscopic, apically flattened setae; gastric, cardiac, branchial, intestinal regions unclearly separated from each other. Gastric region ( Figs. 2C View FIGURE 2 , 3C View FIGURE 3 ) moderately elevated, sometimes with oblique row of dense hooked setae on either side of midline ( Figs. 2A, C View FIGURE 2 , 3A, C View FIGURE 3 , 12B, C View FIGURE 12 ); mesogastric, metagastric, protogastric region on both sides each with rudimentary protuberance ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ). Hepatic, cardiac, branchial regions ( Figs. 2C View FIGURE 2 , 3C View FIGURE 3 ) moderately elevated; mesobranchial region weakly elevated, not higher than gastric region, with two rudimentary tubercles apically, mesial one larger than lateral one ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ); metabranchial region slightly elevated, without tubercles. Intestinal region ( Figs. 2A, C View FIGURE 2 , 3A, C View FIGURE 3 ) slightly elevated, fused with cardiac region.
Pseudorostral spines ( Figs. 2A View FIGURE 2 , 3A, C View FIGURE 3 ) short, length 0.2–0.3 of post-pseudorostral carapace length (PRL/PCL = 0.2±0.0, N = 33), each with two rows of dense, hooked setae on proximal half dorsally, single row of simple, long setae on proximal half mesially; lateral margins subparallel or slightly divergent. Preorbital spine ( Figs. 3A View FIGURE 3 , 4A, B View FIGURE 4 ) elongated, acuminate at tip, directed anterolaterally ( Fig. 4A View FIGURE 4 ). Supraorbital eave ( Figs. 3A View FIGURE 3 , 4A, B View FIGURE 4 ) extended laterally, distinctly concave on lateral margin, weakly truncated on posterior end. Orbital hiatus ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 , 4A View FIGURE 4 ) small, triangular concavity. Postorbital lobe ( Figs. 2A, C View FIGURE 2 , 3A, C View FIGURE 3 , 4A View FIGURE 4 ) small, triangular, much shorter than preorbital spine, weakly compressed dorsoventrally, directed anterolaterally, incurved distally. Hepatic lobe ( Figs. 2A, C View FIGURE 2 , 3 View FIGURE 3 A–C, 4A) not demarcated from hepatic region, broad, triangular, almost three times longer than postorbital lobe (HpL/PoL = 2.8±0.3, N = 9), compressed dorsoventrally, directed anteriorly, obtuse at tip, directed anterolaterally. Anterolateral carapace margin ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ) often with short rows of hooked setae ( Figs. 3C View FIGURE 3 , 12B, C View FIGURE 12 ); lateral surface inferior to anterolateral margin with 1–3 (2 in general) spines ( Figs. 2A, C View FIGURE 2 , 3A, C View FIGURE 3 ). Epibranchial spine ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ) slightly longer than, or as long as postorbital lobe, distinctly shorter than hepatic lobe, directed anterolaterally, slightly incurved, obtuse at tip, positioned at posterior 0.4 of postorostral carapace length (ESL/PCL = 0.4±0.0, N = 10), confluent to posterolateral carapace margin basally. Posterolateral carapace margin ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ) faintly convex. Posterior carapace margin ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 ) projected roundly.
Subhepatic region ( Fig. 3B View FIGURE 3 ) expanded anterolaterally, fully exposed in ventral view, sometimes with group of sparse, hooked setae. Pterygostomian region ( Fig. 3B View FIGURE 3 ) not particularly inflated, with 3–5 (4 in general) papiliform tubercles along pleural suture. Anterolateral angle of buccal frame moderately produced anteriorly ( Fig. 3C View FIGURE 3 ), not overlapped by anterolateral angle of merus of third maxilliped when closed.
Basal antennal article ( Figs. 2B, D View FIGURE 2 , 4B View FIGURE 4 ) smooth on surface, bearing low, blunt longitudinal ridge mesial to midline; distolateral angle moderately produced into small spine directed anterolaterally; lateral margin thickened dorsoventrally in distolateral part ( Fig. 3C View FIGURE 3 ), bearing to posterior orbital margin, with low tubercle basally. Antennal peduncle ( Fig. 4D View FIGURE 4 ) consisting of two articles; penultimate article ( Figs. 2D View FIGURE 2 , 4D View FIGURE 4 ) weakly compressed on lateral half, distal end as broad as proximal end; ultimate article ( Fig. 4D View FIGURE 4 ) two-thirds of penultimate article in length, weakly compressed, slightly broadened distally.
Third maxilliped ( Figs. 3B View FIGURE 3 , 4E View FIGURE 4 ) smooth on surface. Ischium with shallow, broad median depression, lateral margin shallowly concave. Merus with dilated, moderately upturned anterolateral angle. Exopod more than half of ischium in maximum width, gradually narrowed in distal half, mesial margin with subacute angle on distal one-third ( Fig. 4E View FIGURE 4 ).
Chelipeds ( Figs. 2A, B View FIGURE 2 , 3A, B View FIGURE 3 ) similar in size and shape. Ischium ( Fig. 3B View FIGURE 3 ) strongly swollen ventrally in distal half; mesial margin obtusely ridged, without dentation; distolateral lobe distinct, compressed, rounded apically. Merus ( Fig. 5 View FIGURE 5 D–G) prismatic, length 2.5 longer than width (2.5±0.1, N = 11); dorsal surface ( Fig. 5 View FIGURE 5 D–F) with broad, longitudinal keel with 3–4 (3 in general) low, lamellar teeth, distalmost lowest; outer surface ( Fig. 4 View FIGURE 4 E–G) rugose, with unarmed, blunt longitudinal ridge; ventral surface ( Fig. 5F, G View FIGURE 5 ) with blunt ridge bearing 2–3 (2 in general) low, broad teeth; inner surface ( Fig. 5D, E, G View FIGURE 5 ) unarmed, irregularly rugose, with narrow, longitudinal ridge proximally ended in indistinct lobe; distal margin ( Fig. 5 View FIGURE 5 E–G) with 2 prominent knobs at articulation with carpus (outer knob larger than inner), prominent, obliquely erect, subrectangular lobe with short, acute projection on upper side. Carpus ( Fig. 5 View FIGURE 5 A–C) moderately inflated, with indistinct ridge on dorsal surface ( Fig. 5A View FIGURE 5 ); outer margin obtusely ridged, divided into 2 lobes by broad concavity ( Fig. 5A, B View FIGURE 5 ); ventral surface ( Fig. 5C View FIGURE 5 ) smooth; inner margin obtusely crested, divided into broad, distal lobe and prominent, proximal lobe ( Fig. 5A View FIGURE 5 ). Chelae ( Fig. 6A View FIGURE 6 ) almost twice longer than high (ChL/ChH = 2.0±0.1, N = 29); palm strongly expanded, upper margin obtusely ridged, lower margin poorly defined; immovable fingers with subpentagonal teeth; movable finger uniformly dentate on distal two-thirds, with 2 large, isolated teeth on proximal one-third; fingers widely gaped in proximal two-thirds when closed ( Figs. 2B View FIGURE 2 , 6A View FIGURE 6 ).
Ambulatory legs ( Figs. 3A View FIGURE 3 , 7 View FIGURE 7 A–C) decreasing in length posteriorly, surface generally smooth to naked eyes but closely covered with microscopic, apically-flattened setae. Meri subcylindrical, each with rudimentary, upper distal tubercle ( Fig. 7A View FIGURE 7 ), more than five times longer than height in P2 (5.2±0.5, N = 11), more than three times in P3 (3.5±0.3, N = 11). Carpi each with faint, medial depression on extensor surface, most distinct in P2 ( Fig. 7B View FIGURE 7 ). Propodi weakly flattened in P2 and P3, subcylindrical in P4 and P5, each with setal tufts on proximal 0.8 on flexor margin in P2, 0.6 in P3–5 ( Fig. 7A, C View FIGURE 7 ). Dactyli each with two rows of large, calcareous spines on flexor surface ( Fig. 7A, C View FIGURE 7 ).
Thoracic sternites ( Figs. 2B View FIGURE 2 , 3B View FIGURE 3 , 8A, B View FIGURE 8 ) smooth on surface, with shallow, broad depression on second to fourth sternites on both sides ( Fig. 8A View FIGURE 8 ); second sternite with pair of small depression anteriorly (often continuous to shallow depression on second to fourth sternites); third, fourth sternites faintly ridged medially ( Fig. 8A View FIGURE 8 ); sterno-pleonal cavity without long setae on anterolateral margins ( Fig. 8A View FIGURE 8 ).
Pleon ( Figs. 2B View FIGURE 2 , 3B View FIGURE 3 , 8B View FIGURE 8 ) with six pleomeres and telson; third to sixth pleomeres fixed, with distinct suture. Third pleomere broadest, lateral margins arcuate; fourth pleomere trapezoid, shorter than fifth in midline length; fifth pleomere trapezoid; sixth pleomere rectangular, 0.6 of third pleomere in proximal width (0.6±0.0, N = 22); telson triangular.
Shaft of G1 ( Fig. 9 View FIGURE 9 A–D) straight, trilobate in distal one-sixth; dorsal lobe elongate triangular, more than twice longer than ventral lobe, weakly curved inwards ( Fig. 9D View FIGURE 9 ); ventral lobe triangular, with subacute tip ( Fig. 9B, C View FIGURE 9 ); mesial lobe as long as ventral lobe, projecting nearly perpendicular to dorsal lobe, strongly curled downwards ( Fig. 9B, D View FIGURE 9 ); hiatus between dorsal, mesial lobes wide ( Fig. 9D View FIGURE 9 ); mesial, lateral margins from dorsal to ventral lobe clearly concave medially; lateral margin higher than mesial margin, dilated in median part ( Fig. 9 View FIGURE 9 B–D). Shaft of G2 ( Fig. 9E, F View FIGURE 9 ) stout, slightly narrowed distally, truncated apically; apex with relatively large, triangular, subacute projection.
Adolescent males (9.9–20.3 mm PCL) ( Fig. 10A, B View FIGURE 10 ). Chelae ( Fig. 6B View FIGURE 6 ) proportionally longer (ChL/ChH = 2.4±0.0, N = 11) than in full-grown specimens ( Table 1 View TABLE 1 ), not gaped, both fingers uniformly dentate on cutting margin ( Figs. 6B View FIGURE 6 , 10A, B View FIGURE 10 ). Pleon subtriangular (PW6/PW3 = 0.6±0.0, N = 8) ( Fig. 10B View FIGURE 10 ). G1 apically trilobate as in full-grown males.
Immature males (<6.2 mm PCL) ( Fig. 11A, B View FIGURE 11 ). Carapace relatively slender (PCL/CW = 1.4±0.1, N = 7) ( Table 1 View TABLE 1 ). Chela similar to adolescent specimens in dentation on both fingers, but more slender (ChL/ChH = 2.7±0.1, N = 5) than in adolescent specimens ( Table 1 View TABLE 1 , see also Fig. 11A, B View FIGURE 11 ). Pleon subtriangular (PW6/PW3 = 0.7±0.0, N = 5) ( Fig. 11B View FIGURE 11 ). G1 incompletely folded ( Fig. 9I, J View FIGURE 9 ), otherwise folded but provided with undeveloped mesial lobe projecting anterolaterally ( Fig. 9G, H View FIGURE 9 ).
Female. Full-grown females (12.7–22.2 mm PCL, including paralectotypes). Carapace ( Fig. 3D, F View FIGURE 3 ) similar to males in general proportion (PCL/CW = 1.3±0.0, N = 27; HpL/PoL = 2.7±0.1, N = 12; ESL/PCL = 0.4±0.0, N = 12) (Student t -test, p> 0.05); hepatic, mesobranchial regions more elevated than in males ( Fig. 3D, F View FIGURE 3 ); anterolateral margins, protogastric, subhepatic regions usually with oblique rows of hooked setae ( Fig. 3 View FIGURE 3 D–F). Cheliped merus slightly slenderer than in males (length/height = 2.6±0.1, N = 7) though there was no significant difference (Student t -test, p = 0.12); chelae ( Fig. 6C View FIGURE 6 ) smaller, more slender than in full-grown males (ChL/ChH = 2.6±0.1, N = 17; Student t -test, p <0.01), both fingers uniformely dentated on cutting edges, not gaped when closed. Tufts of few elongate setae sometimes on midline of gastric region, midpoint of metabranchial region, apex of epibranchial spines, summits of cardiac and intestinal regions ( Fig. 13C View FIGURE 13 ). Pleon ( Figs. 3E View FIGURE 3 , 13D View FIGURE 13 ) with six pleomeres and telson, expanded (PW6/PW3 = 1.6±0.1, N = 10). Gonopores ( Fig. 8D, E View FIGURE 8 ) comma-shaped, nearly circular in lateral twothirds, elongate in mesial one-third.
Adolescent females (9.6–16.9 mm PCL) ( Fig. 10C, D View FIGURE 10 ). Chela more slender than in adolescent males (ChL/ChH = 2.6±0.0, N = 5; Student t -test, p = 0.02) ( Fig. 10D View FIGURE 10 ), both fingers uniformly dentate as in full-grown individuals. Pleon ovate (PW6/PW3 = 1.2±0.1, N = 3) ( Fig. 10D View FIGURE 10 ).
Immature females (<6.2 mm PCL) ( Fig. 11C, D View FIGURE 11 ). Carapace relatively slender (PCL/CW = 1.4±0.0, N = 3) ( Table 1 View TABLE 1 ). Chela slender (ChL/ChH = 2.7±0.0, N = 3) ( Table 1 View TABLE 1 , see also Fig. 11D View FIGURE 11 ). Pleon generally rectangular, with subtriangular telson, lateral margins subparralel (PW6/PW3 = 1.2±0.1, N = 3) ( Table 1 View TABLE 1 , see also Fig. 11D View FIGURE 11 ).
Variations. Carapace gets broader (PCL/CW decreases from 1.4 to 1.2) in relation to size growth in both sexes ( Table 1 View TABLE 1 ; see also Figs. 3 View FIGURE 3 , 10 View FIGURE 10 , 11 View FIGURE 11 , 12 View FIGURE 12 ). Gastric, mesobranchial, and metabranchial regions are less elevated in smaller specimens, often missing tubercles or protuberances on each surface. Mesobranchial region is rarely elevated ( Fig. 12C View FIGURE 12 ). Supraorbital eave rarely straight on lateral margin ( Fig. 12B View FIGURE 12 ). Hepatic lobes are sometimes laminate ( Fig. 12 View FIGURE 12 B–D), relative length against postorbital lobe (HpL/PoL) increases from 2.0 to 3.6 in relation to size growth ( Fig. 39 View FIGURE 39 ). Groups of hooked setae on either side of gastric region and anterolateral margin of carapace, as well as elongate setae on the tops of mesobranchial, intestinal regions, are likely to be reduced in large specimens in both sexes ( Figs. 3A View FIGURE 3 , 13A View FIGURE 13 vs. Figs. 9A View FIGURE 9 , 10A View FIGURE 10 , 12 View FIGURE 12 A–C). Ambulatory legs often with a few, elongate setae on upper margins of meri, upper, lower surfaces of carpi in small specimens of both sexes ( Figs. 11 View FIGURE 11 , 13F View FIGURE 13 ).
Size. Largest male: 32.1 × 26.0 mm; largest female: 20.9 × 16.1 mm; smallest ovigerous female: 11.1 × 8.9 mm ( Ohtsuchi et al. 2018; this study).
Coloration in life. Carapace ( Fig. 13A, D, F View FIGURE 13 ) generally dark red, reddish brown, or dark green, each simlar to general coloration of collected habitats ( Fig. 13C View FIGURE 13 ); small specimens with some blotches and/or dense speckles ( Fig. 13F View FIGURE 13 ); female specimen often with irregular speckles, mottling even in full-grown specimens ( Fig. 13D View FIGURE 13 ); patterning reduced in males especially in large specimens ( Fig. 13A View FIGURE 13 ). General coloration of thorax, pleon, chelipeds, ambulatory legs similar to carapace in males ( Fig. 13B View FIGURE 13 ), often slightly reduced in third maxilipeds, thoracic plastron, flexor surface of cheliped meri, ambulatory leg meri in females ( Fig. 13E View FIGURE 13 ). Cheliped meri, palm often with scale-like pattering in full-grown males ( Fig. 13B View FIGURE 13 ). Ambulatory legs with whitish band on the joint of each articulation and the distal parts of dactyli. See also Miyake (1983, 1998: pl. 12, fig. 5, as P. nipponensis ), Wada (1995: pl. 103, fig. 5), Minemizu (2000, 2002: 209, unnumbered figure), Watanabe (2014: unnumbered figure), and Yoshizaki (2018: 48, 173, 181, unnumbered figures).
Distribution. Pacific coast of Japan, from Iwaki, Fukushima to Osaka Bay, including Izu Ohshima Island; Seto Inland Sea; coast of Sea of Japan from Toyama Bay to Nagasaki; East China Sea from Amakusa Archipelago to southeast coast of China mainland (Fuzhou), including Kume-jima Island, Ryukyu.
Habitat. Intertidal to 8 m depth. Commonly found in various kind of macroalgal communities developed on rocky reefs, such as brown algal beds ( Sargassaceae : Sargassum fusiformes , S. hemiphyllum , and S. hornei ; and Dictyotacea: Padina alborescens ) ( Fig. 13C View FIGURE 13 ), red algal turfs ( Phyllophoraceae : Ahnfeltiopsis paradoxa ; Gelidiaceae : Gelidium elegans ; and Halymeniaceae : Grateloupia cornea ), and sometimes articulated coralline algal turfs ( Corallinaceae : Corallina pilulifera , C. crassissima ) ( Sato & Wada 2000; Ohtsuchi et al. 2018; this study).
Decorating materials. Pieces of various macroalgae, mainly of Sargassum spp. or branched red algae on the coast of Shirahama, Kii Peninsula, and Nagai, Sagami Bay ( Sato & Wada 2000; this study).
Ecological notes. This species was suggested as herbivorous ( Sato & Wada 2000), but we found they infrequently feed on the sea urchin Hemicentrotus pulcherrimus (A. Agassiz, 1864) under captive conditions (N. Ohtsuchi pers. obs.). Juveniles and smaller individuals usually inhabit subtidal turfs of various branched red algae e.g. gelidiaceans and Grateloupia cornea , and the majority move to Sargassum beds near low tidal marks on the coast of Miura and Kii Peninsulas ( Ohtsuchi et al. 2018). On the other hand, the individuals of all the ontogenetic stages were collected sympatrically from dense turfs of red algae ( Ahnfeltia paradoxa , Gelidium elegans , and G. cornea ) near low tidal mark on the rocky shore coast of Iwaki (Fukushima, Pacific coast of northeast Japan), Oarai (Ibaraki, Pacific coast of east Japan), and Tsurugizaki (Kanagawa, Miura Peninsula, Pacific coast of east Japan).
Remarks. De Haan (1837, 1839) did not designate any type when the species was described. The larger male in the syntype materials (RMNH D 42298) was subsequently designated as the lectotype by Yamaguchi & Baba (1993: 353). The lectotype ( Fig. 2 View FIGURE 2 A–D) is a large male with enlarged chelipeds, viz. full-grown male. It is remarkable that the movable finger has two large, isolated teeth subproximally ( Fig. 2B View FIGURE 2 ), which is also shown in the plate of De Haan (1837: pl. 24, fig. 2, as Pisa (Halimus) quadridens ). On the other hand, the smaller male, which was designated as paralectotype by Yamaguchi & Baba (1993: fig. 113a), is considerably different from the lectotype in many morphological characters, and it should have been attributed to P. intermedia instead ( Fig. 2E, F View FIGURE 2 ).



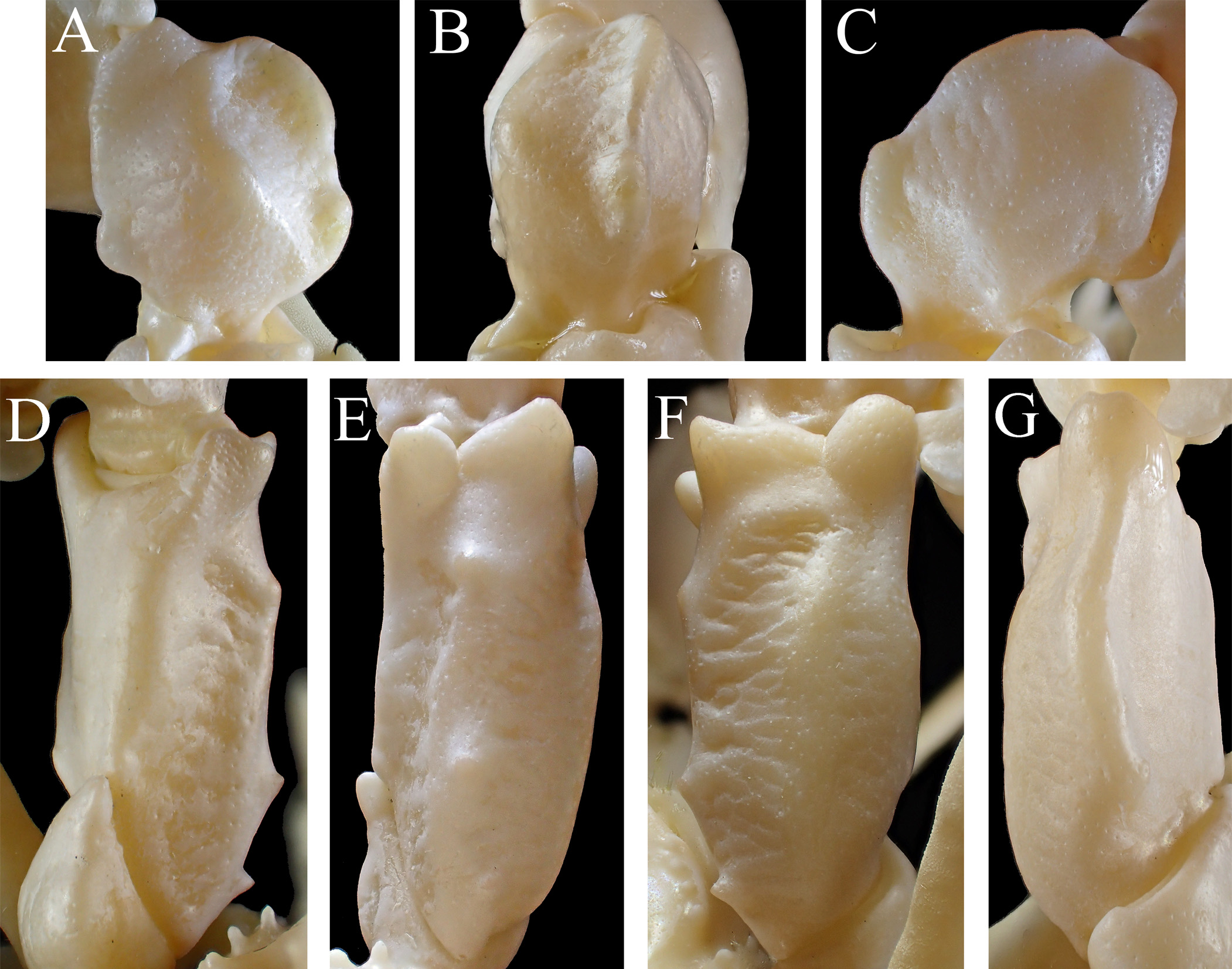
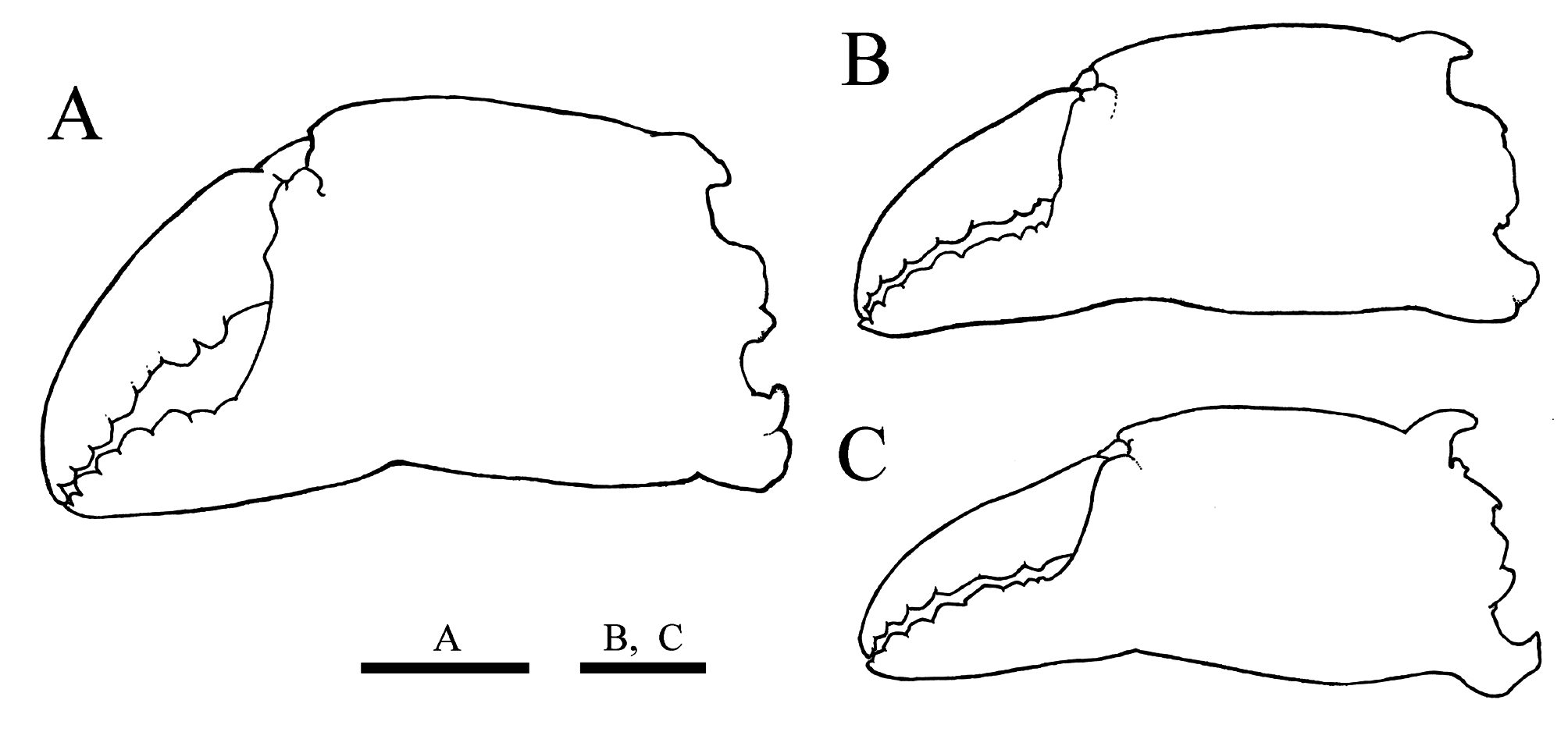
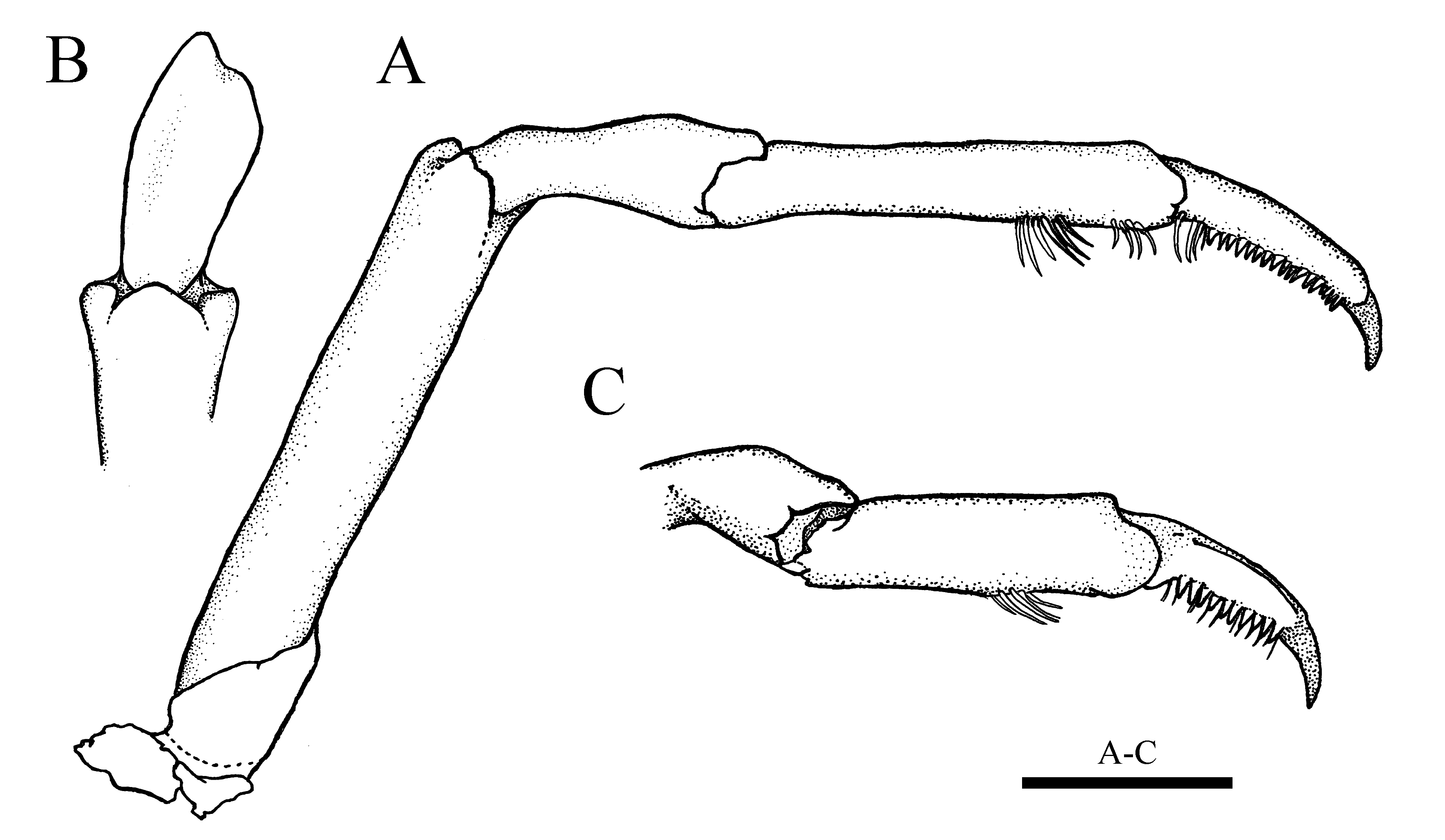
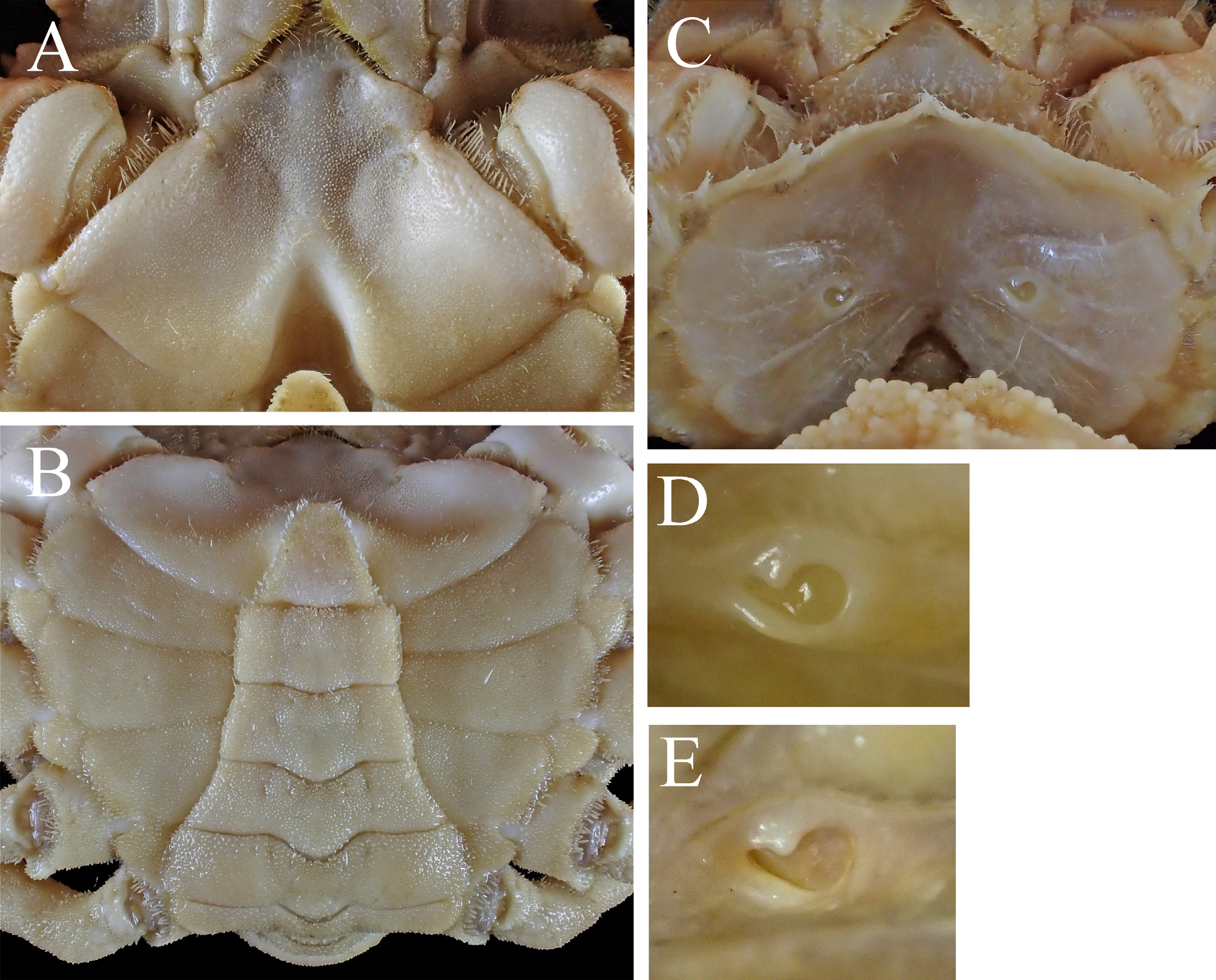





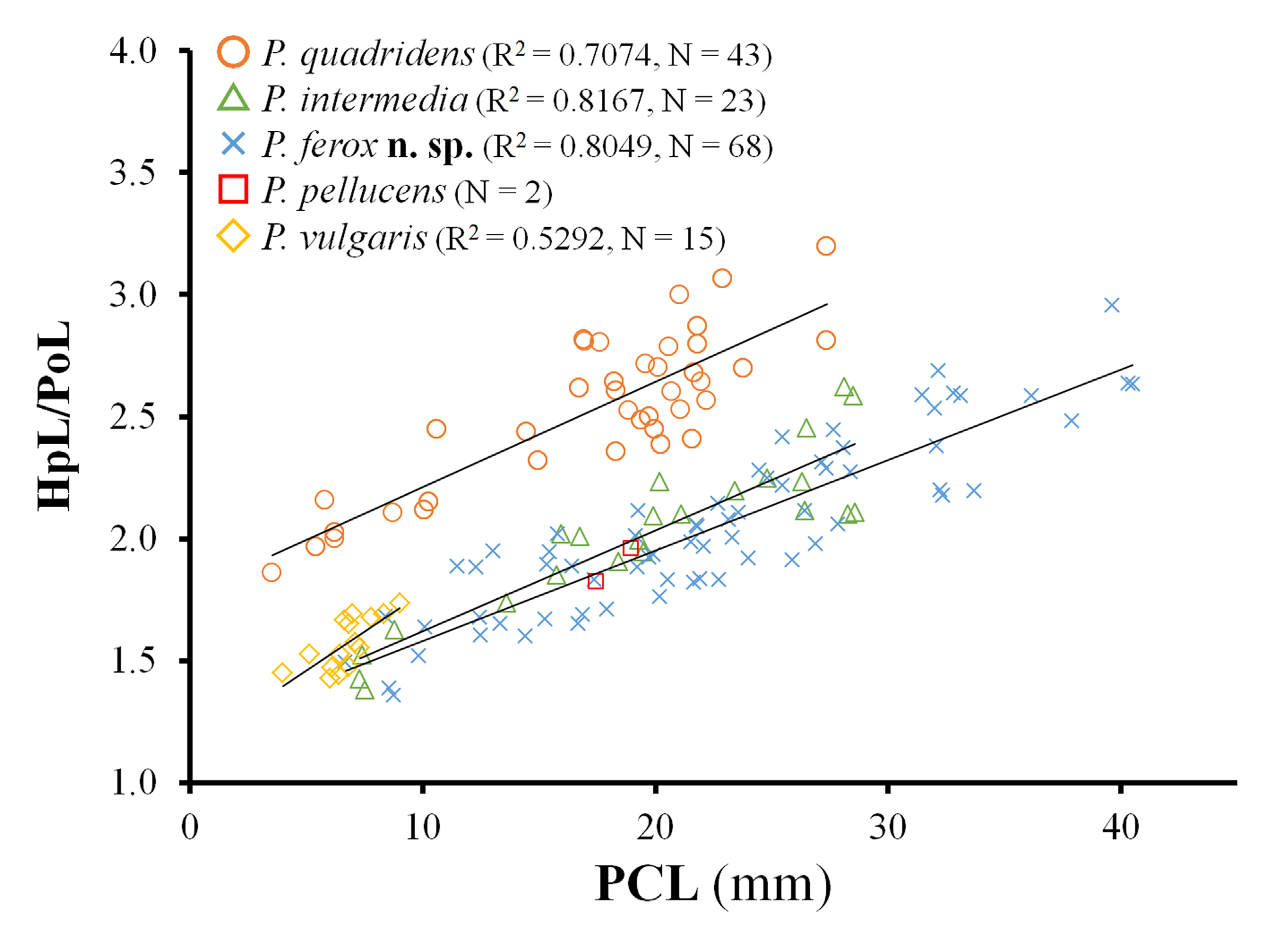
TABLE 1. Comparison of ontogenetic change in the proportion of carapace, pseudorostral spine, chela, and pleon among Pugettia quadridens, P. intermedia and P. ferox n. sp.
| Sex | OS | Pugettia quadridens Mean SD N | Mean | Pugettia intermedia SD N | Mean | Pugettia ferox SD | N | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCL/CW | male | immature | 1.4 | ± | 0.1 | (7)a,A | 1.4 | ± | 0.1 | (3)a,A | 1.4 | ± | 0.0 | (5)a,A |
| adolescent | 1.3 | ± | 0.0 | (17)b,A | 1.3 | ± | 0.1 | (8)ab,A | 1.3 | ± | 0.0 | (30)b,A | ||
| full-grown | 1.3 | ± | 0.0 | (52)b,A | 1.3 | ± | 0.1 | (18)b,AB | 1.2 | ± | 0.0 | (19)c,B | ||
| female | immature | 1.4 | ± | 0.0 | (3)a | 1.4 | (2) | 1.4 | ± | 0.0 | (8)a | |||
| adolescent | 1.3 | ± | 0.0 | (7)a | 1.3 | (1) | 1.3 | ± | 0.0 | (4)a | ||||
| full-grown | 1.3 | ± | 0.0 | (27)b,A | 1.3 | ± | 0.1 | (18)A | 1.3 | ± | 0.0 | (16)b,A | ||
| PRL/PCL | male | immature | 0.2 | ± | 0.0 | (5)a,A | 0.3 | ± | 0.0 | (3)a,B | 0.2 | ± | 0.0 | (3)a,A |
| adolescent | 0.2 | ± | 0.0 | (12)a,A | 0.3 | ± | 0.0 | (9)a,B | 0.2 | ± | 0.0 | (30)a,A | ||
| full-grown | 0.3 | ± | 0.0 | (33)a,A | 0.3 | ± | 0.0 | (17)a,B | 0.2 | ± | 0.0 | (20)a,A | ||
| female | immature | 0.2 | ± | 0.0 | (3)a | 0.2 | (2) | 0.2 | ± | 0.0 | (7)a | |||
| adolescent | 0.2 | ± | 0.0 | (6)a | 0.2 | (1) | 0.2 | ± | 0.0 | (4)a | ||||
| full-grown | 0.2 | ± | 0.0 | (23)a,A | 0.2 | ± | 0.0 | (13)A | 0.2 | ± | 0.0 | (14)a,A | ||
| ChL/ChH | male | immature | 2.7 | ± | 0.1 | (5)a,A | 2.9 | ± | 0.1 | (3)a,AB | 2.9 | ± | 0.1 | (3)a,B |
| adolescent | 2.4 | ± | 0.0 | (11)b,A | 2.6 | ± | 0.1 | (9)b,AB | 2.8 | ± | 0.2 | (30)b,B | ||
| full-grown | 2.0 | ± | 0.1 | (29)c,A | 2.2 | ± | 0.2 | (15)c,B | 2.3 | ± | 0.1 | (19)c,B | ||
| female | immature | 2.7 | ± | 0.0 | (3)a | 2.9 | (2) | 2.8 | ± | 0.1 | (7)a | |||
| adolescent | 2.6 | ± | 0.0 | (5)a | 2.7 | (1) | 2.8 | ± | 0.1 | (8)a | ||||
| full-grown | 2.6 | ± | 0.1 | (17)a,A | 3.0 | ± | 0.2 | (15)B | 2.9 | ± | 0.2 | (16)a,B | ||
| PW6/PW3 | male | immature | 0.7 | ± | 0.0 | (5)a,A | 0.7 | ± | 0.0 | (3)a,A | 0.6 | ± | 0.1 | (3)a,B |
| adolescent | 0.6 | ± | 0.0 | (8)b,A | 0.7 | ± | 0.0 | (6)a,B | 0.5 | ± | 0.0 | (11)b,C | ||
| full-grown | 0.6 | ± | 0.0 | (22)a | 0.7 | ± | 0.0 | (4)a | 0.5 | ± | 0.0 | (11)b | ||
| female | immature | 1.2 | ± | 0.1 | (3)a | 1.1 | - | 1.3 | (2) | 1.1 | ± | 0.1 | (9)a | |
| adolescent | 1.2 | ± | 0.1 | (3)a | 1.3 | (1) | 1.3 | ± | 0.1 | (7)b | ||||
| full-grown | 1.6 | ± | 0.1 | (10)b,A | 1.5 | ± | 0.1 | (5)B | 1.4 | ± | 0.1 | (8)b,C | ||
OS, ontogenetic stage; N, sample size; PCL, postrostral carapace length; CW, maximum carapace width; PRL, length of pseudorostral spine; ChL, chela length; ChH, chela height; PW3, PW6, proximal width of pleomere 3 and 6, respectively. Means in the same row followed by different small letters are significantly different from each other (Tukey-Kramer HSD test or Steel-Dwass test, p <0.05). Means in the same line followed by diffferent capital letters are significantly different from each other (Tukey-Kramer HSD test or Steel-Dwass test, p <0.05). Differently colored boxes indicate significant difference between sexes (Student t -test or Welch two samples t -test, p <0.05).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pugettia quadridens (De Haan, 1837)
| Ohtsuchi, Naoya & Kawamura, Tomohiko 2019 |
Pugettia quadridens pellucens
| Marumura, M. & Kosaka, A. 2003: 32 |
Pisa (Menoethius)
| Yamaguchi, T. & Baba, K. 1993: 353 |
Pugettia quadridens quadridens
| Ito, S. & Honma, Y. 2001: 27 |
| Kim, H. S. & Kim, I. H. 1986: 325 |
Pugettia nipponensis
| Miyake, S. 1983: 36 |
Pugettia quadridens
| Yoshizaki, K. 2018: 30 |
| Watanabe, T. 2014: 41 |
| Wicksten, M. K. & Stachowicz, J. J. 2013: 359 |
| Nunomura, N. 2010: 52 |
| Ng, P. K. L. & Guinot, D. & Davie, P. J. F. 2008: 101 |
| Yamaguchi, T. & Henmi, Y. 2008: 80 |
| Takeda, M. & Komai, T. & Komatsu, H. & Ikeda, H. 2006: 196 |
| Ikeda, H. & Kuramochi, T. 2004: 12 |
| Marumura, M. & Kosaka, A. 2003: 32 |
| Minemizu, R. 2000: 209 |
| Muraoka, K. 1998: 24 |
| Yamaguchi, T. & Baba, K. 1993: 353 |
| Yamaguchi, T. & Harada, K. & Takeda, M. & Kikuchi, T. 1987: 13 |
| Griffin, D. J. G. & Tranter, H. A. 1986: 97 |
| Takeda, M. 1982: 120 |
| Ikeda, H. 1981: 15 |
| Yamaguchi, T. & Takeda, M. & Tokudome, K. 1976: 35 |
| Sakai, T. 1936: 88 |
Pugettia quadridens
| Komai, T. 1999: 86 |
| Kim, H. S. & Chang, C. Y. 1985: 45 |
| Kim, W. & Kim, H. S. 1982: 146 |
| Kim, H. S. & Rho, B. J. & Hong, S. Y. & Shin, S. & Han, C. H. 1979: 110 |
| Kamita, T. 1935: 63 |
| Sakai, T. 1934: 294 |
| Urita, T. 1926: 32 |
| Balss, H. 1924: 24 |
| Ortmann, A. 1893: 43 |
| Miers, E. J. 1879: 23 |
| Stimpson, W. 1857: 219 |
Menaethius quadridens
| Adams, A. & White, A. 1849: 20 |