
Eohyosaurus undetermined
|
publication ID |
https://doi.org/ 10.1111/zoj.12246 |
|
persistent identifier |
https://treatment.plazi.org/id/03D887CB-FFDF-744F-914C-46ACFDC9ACED |
|
treatment provided by |
Carolina |
|
scientific name |
Eohyosaurus undetermined |
| status |
|
EOHYOSAURUS WOLVAARDTI GEN. ET SP. NOV.
( FIGS 2–5 View Figure 2 View Figure 3 View Figure 4 View Figure 5 )
Etymology
Genus name from the Greek eos, meaning ‘early’ or ‘dawn’, hyos, meaning ‘pig’ or ‘hog’, and sauros, meaning ‘lizard’ or ‘reptile’. Name refers to the common description of rhynchosaurs as ‘pig-reptiles’ and the early stratigraphical occurrence of the new taxon. The species is named after Frederik Petrus Wolvaardt, who discovered the type and only known specimen in December 2000. The name is therefore intended to translate as ‘Wolvaardt’s early pig-reptile’.
Holotype
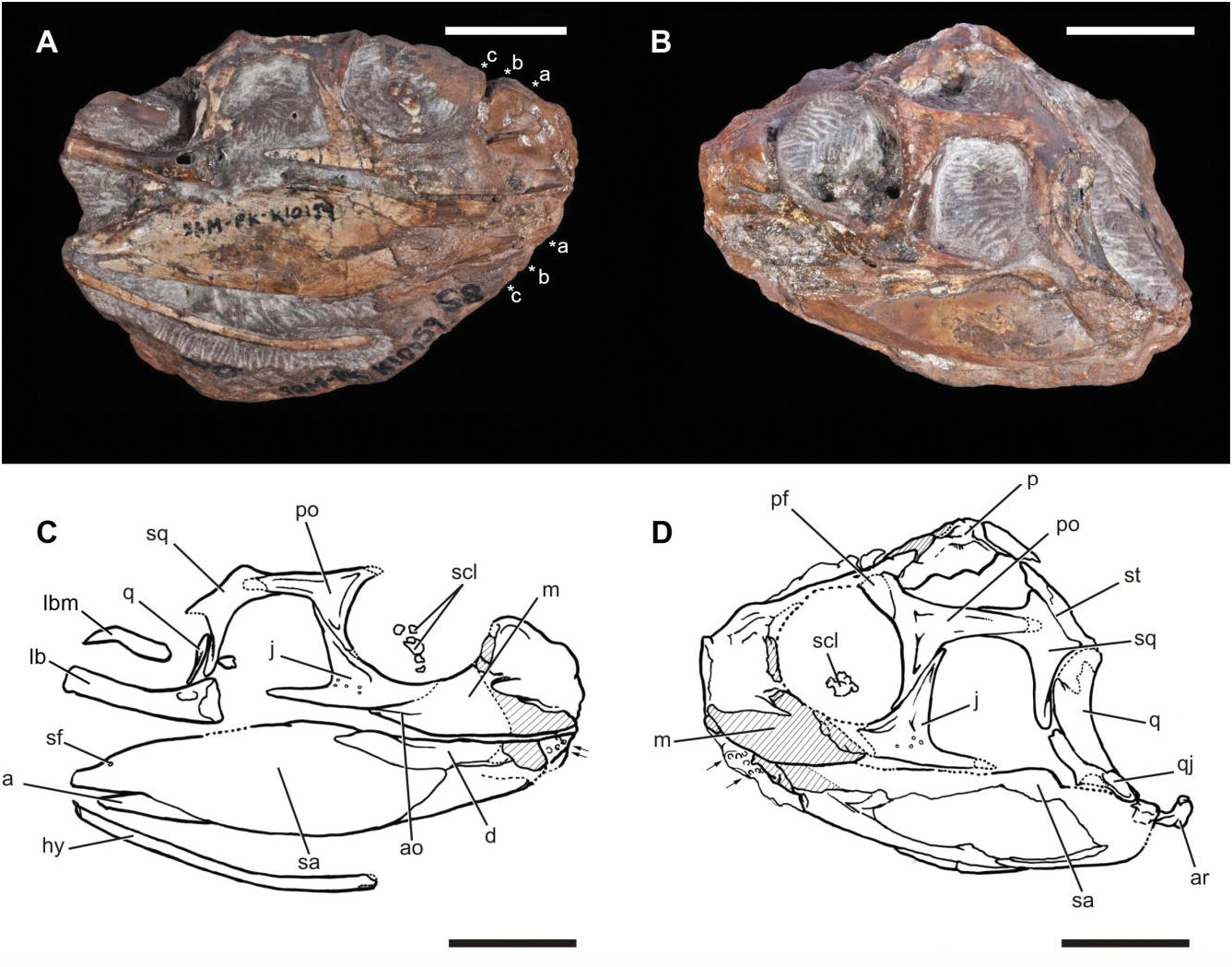
SAM-PK-K10159, partial skull missing the anterior end, with associated incomplete lower jaws ( Figs 2–5 View Figure 2 View Figure 3 View Figure 4 View Figure 5 ).
Locality and horizon
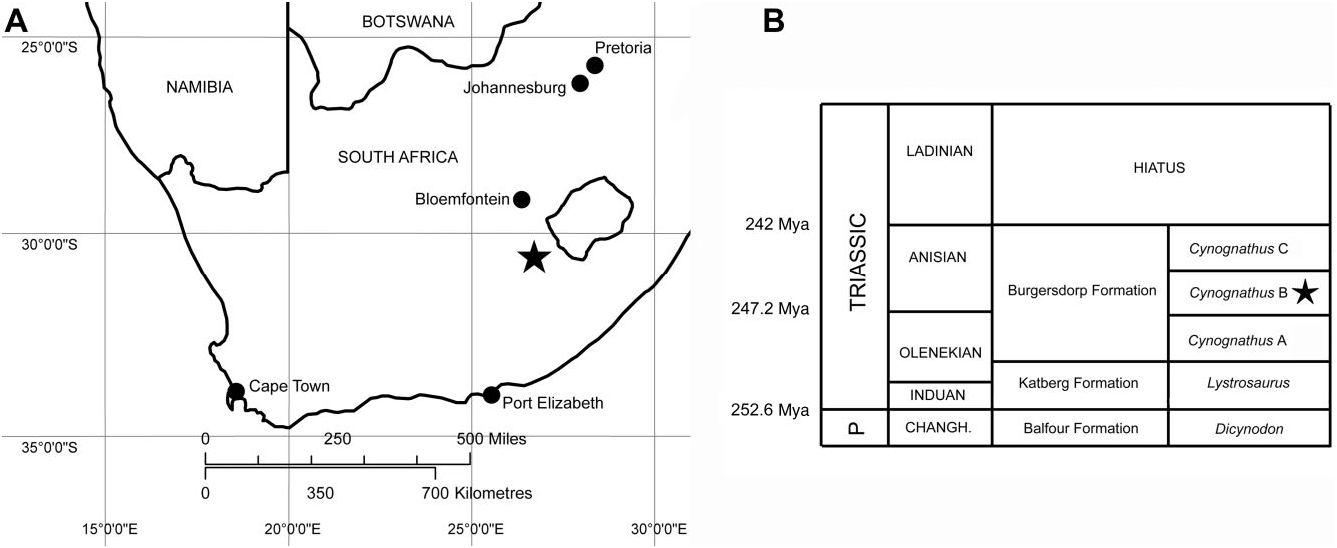
Farm Lemoenfontein 44, Rouxville District, Free State Province, South Africa. Burgersdorp Formation, Cynognathus Assemblage Zone, Subzone B (early Middle Triassic: early Anisian; Fig. 1 View Figure 1 ).
The specimen was found as loose float on boulderstrewn slopes at the base of a cliff (coordinates: 30°36′43.7′′S, 26°37′22.9′′E). The same locality has also yielded remains of the archosauriform Erythrosuchus , the cynodont Trirachodon , the kannemeyeriid dicynodont Kannemeyeria , procolophonids, and the bauriid therocephalian Microgomphodon oligocynus (S. Kaal, pers. comm. 2013; D. Wolvaardt, pers. comm. 2013; Abdala et al., 2014).
Diagnosis
Characterized by the following autapomorphy: jugal with elongate dorsal process that forms the entire anterior margin of the infratemporal fenestra and that articulates anteriorly with the entire posterior margin of an elongate ventral process of the postorbital. Distinguished from other rhynchosaurs on the basis of the following unique combination of characters: (1) maxillae and dentaries mediolaterally expanded (present in Howesia browni and Rhynchosauridae sensu Dilkes, 1998 ; mediolaterally expanded maxillae/dentaries absent in Mesosuchus browni ); (2) teeth present on the occlusal and lingual surfaces of the maxillae and dentaries (present in H. browni and Rhynchosauridae ; occlusal teeth only present in M. browni ); (3) maxilla lacks a longitudinal occlusal groove and dentary lacks occlusal blade (groove and blade present in Rhynchosauridae ; groove and blade also absent in M. browni and H. browni ); (4) occlusal margin of maxilla offset ventrally from the ventral margin of the main body of the jugal (occlusal margin not offset ventrally in M. browni ; offset ventrally in Rhynchosauridae , and possibly in H. browni ); (5) presence of a short anguli oris crest on the lateral surface of the maxilla (crest absent in M. browni ; present in Rhynchosauridae but placed on the jugal; condition uncertain in H. browni ); (6) posterior process of jugal is short and terminates at approximately 50% of the anteroposterior length of the infratemporal fenestra (process proportionately longer in M. browni , and contacts quadratojugal in most Rhynchosauridae ; process is also short in H. browni ); (7) elongate posterior process of the postorbital terminates above the anterior margin of the ventral process of the squamosal (posterior process shorter and does not terminate above the ventral process of the squamosal in M. browni and H. browni ; present in Rhynchosauridae ); (8) elongate ventral process of the squamosal extends for more than 50% of the posterior margin of the infratemporal fenestra (ventral process does not extend for more than 50% of the posterior margin in H. browni ; present in M. browni and Rhynchosauridae ); (9) sagittal crest on parietal (sagittal crest absent in M. browni ; present in H. browni and Rhynchosauridae ).
Description
General
SAM-PK-K10159 is relatively complete posteriorly, but the anterior end of the skull is incomplete ( Figs 2 View Figure 2 , 3 View Figure 3 ). On the left side, the premaxilla and anterior half of the maxilla are entirely missing, the external surface of the posterior half of the maxilla has broken away, the prefrontal and lacrimal are represented by small fragments lacking their external surfaces, and the external surface of the anterior process of the jugal has broken away ( Fig. 2B, D View Figure 2 ). On the right side, the premaxilla and the tip of the maxilla are missing, much of the anterior part of the preserved maxilla is represented by an impression of the medial surface of the element, and only fragments of the lacrimal and prefrontal are preserved ( Fig. 2A, C View Figure 2 ). Dorsally, the nasals, frontals, anterior end of the parietals, and much of the postfrontals are missing, although a natural cast of the endocranium and nasal capsule is exposed. In the lower jaw, the anterior end of the dentary is missing on both left and right sides, and much of the external surface of the left mandible is missing, but some details of the morphology of the latter are revealed by a natural mould. The left side of the skull has additionally been displaced ventrally relative to the right side, and also appears to have been compressed anteroposteriorly. Posteriorly, the parietals appear to have been displaced dorsally relative to the occipital region.
As preserved, the maximum anteroposterior length along the sagittal plane of the skull on the left side, from the broken anterior margin of the maxilla to the posterior margin of the quadrate condyles, is 68.2 mm. Based on the dimensions of the orbit and infratemporal fenestra ( Table 1), SAM-PK-K10159 is similar in size to a referred specimen of the early rhynchosaur M. browni (SAM-PK-6536; Dilkes, 1998) that has a basal skull length (premaxilla to quadrate condyles) of 89 mm, and we suggest that the complete length of SAM-PK- K10159 View Materials would have been similar. Our estimate of skull length suggests that the skull was longer than broad, similar to the condition in M. browni ( Dilkes, 1998) , H. browni ( Dilkes, 1995) , and the rhynchosaurids R. articeps ( Benton, 1990) , S. stockleyi ( Huene, 1938) , and Bentonyx sidensis ( Langer et al., 2010) .
Although the dorsal rim of the orbit is not preserved on either side of SAM-PK-K10159, the orbit appears to be slightly anteroposteriorly longer than dorsoventrally tall on the right side (that of the left side appears to be anteroposteriorly compressed). Despite deformation, the orbits face mostly laterally, unlike the condition in the hyperodapedontines I. genovefae ( Whatley, 2005) , Teyumbaita sulcognathus (UFRGS- PV-0232, UFRGS-PV-0298; Montefeltro et al., 2010) and Hyperodapedon spp. ( Hyperodapedon mariensis , FZB- PV-1867; Hyperodapedon sanjuanensis , MACN-Pv 18185; Hyperodapedon huenei , UFRGS-0132), in which the orbits face mostly dorsally. Fragments of the right sclerotic ring appear to be present in the right orbit, and some fragments of the left sclerotic ring may be present on the left side ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : scl).
The infratemporal fenestra has a trapezoidal outline on the right side (the left side is anteroposteriorly compressed), being longer anteroposteriorly at its ventral margin than at its dorsal margin. The supratemporal fenestra has a broadly rounded anterior margin, expands in transverse width posteriorly reaching a maximum width just posterior to the midlength of the parietal, and then tapers to a rounded posterolateral corner.
Maxilla
The maxilla is poorly preserved on both sides of the skull, but its external morphology is best preserved on the right side ( Figs 2 View Figure 2 , 4 View Figure 4 : m). As in all other rhynchosaurs and non-archosauriform archosauromorphs, an antorbital fenestra and fossa are absent. The occlusal margin is nearly straight in lateral view, but the preserved part of the anterior end of the left maxilla and the impression of the anterior part of the right maxilla suggests that it curved gently dorsally toward the contact with the premaxilla. This curvature is not as marked as typically occurs in rhynchosaurids (e.g. Benton, 1983a, 1984, 1990; Langer & Schultz, 2000; Nesbitt & Whatley, 2004; Montefeltro et al., 2010; Mukherjee & Ray, 2014), but does differ from the completely straight occlusal margin present in M. browni (SAM-PK-6536; Dilkes, 1998). The posterior end of the occlusal margin of the maxilla is offset ventral relative to the ventral margin of the jugal, contrasting with the condition in M. browni (SAM-PK- 6536; Dilkes, 1998), in which the ventral margin of the jugal is in line with the occlusal margin of the maxilla.
The posterior-most portion of the maxilla tapers dorsoventrally, apparently forming an anterodorsallyto-posteroventrally orientated suture with the jugal, although the exact path of this suture is difficult to establish owing to poor preservation. The contact of the maxilla with the lacrimal is unclear on both sides of the skull.
The lateral surface of the maxilla is mostly flat, and is slightly expanded laterally at the occlusal margin. Additionally, the lateral surface of the maxilla bears a distinct ridge ( Figs 2 View Figure 2 , 4 View Figure 4 : ao), here proposed to be homologous to the anguli oris crest of rhynchosaurids ( Sill, 1970; Langer & Schultz, 2000). In Eohyosaurus wolvaardti this ridge extends from the contact of the maxilla with the ventral margin of the jugal, along the lateral surface of the former bone, and terminates anteriorly below the anteroventral corner of the orbit. Below this ridge the lateral surface of the maxilla is slightly bevelled inwards. Although we propose that this feature is homologous with the anguli oris crest, its position is slightly different from that in most rhynchosaurids, where it is positioned on the boundary between the jugal and the maxilla ( Sill, 1970; Langer & Schultz, 2000).
Where the anterior part of the right maxilla has broken away, an impression of the bone demonstrates that the ventral part of this element is strongly transversely expanded, and that this transverse expansion becomes stronger posteriorly. This transverse expansion forms a nearly horizontal, ventrally facing occlusal surface laterally. Medially the occlusal surface forms a near right angle with the lingual surface of the maxilla. This transverse expansion is very similar to that present in H. browni ( Dilkes, 1995) and rhynchosaurids ( Benton, 1984; Langer & Schultz, 2000; Nesbitt & Whatley, 2004; Montefeltro et al., 2010), but differs from the transversely narrower occlusal surface present in M. browni ( Dilkes, 1998) .
Jugal
The jugal forms the ventral margin of the orbit and much of the anterior and ventral margins of the infratemporal fenestra ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : j). The lateral surface of the main body of the jugal possesses several subcircular pits that are irregularly distributed ( Figs 2 View Figure 2 , 4C View Figure 4 ). Similar pits are present on the jugal of H. browni (SAM-PK-5884). The orbital margin is marked by a low thickening that is continuous with a similar thickened margin on the orbital margin of the postorbital. This thickened orbital margin is present to some degree in all rhynchosaurs, but is particularly marked in the basal rhynchosaurids R. articeps (SHYMS G-132/ 1982), R. brodiei (WARMS Gz6097), S. stockleyi ( Huene, 1938) and B. sidensis (BRSUG 27200). However, this orbital thickening is less well developed in F. spenceri ( Benton, 1990) and hyperodapedontines ( Benton, 1983a; Whatley, 2005; Montefeltro et al., 2010).
The posterior process of the jugal is short, and terminates at approximately the anteroposterior midlength of the infratemporal fenestra ( Fig. 4C View Figure 4 : pp.j). This short process is similar to that of H. browni ( Dilkes, 1995) and R. articeps (NHMUK R1236) , but differs from the proportionately longer process of M. browni ( Dilkes, 1998) and most rhynchosaurids ( Benton, 1983a, 1990; Langer & Schultz, 2000; Montefeltro et al., 2010), which extends nearly as far as or beyond the anterior margin of the ventral process of the squamosal (e.g. H. huenei : UFRGS-PV-0132T). The posterior process of the jugal of SAM-PK-K10159 is orientated almost directly posteriorly, as also occurs in M. browni (SAM-PK- 6536) and H. browni (SAM-PK-5884). By contrast, in rhynchosaurids the posterior process of the jugal is more strongly directed lateroventrally as well as posteriorly and, as a result, the temporal region of the skull is considerably broader than the orbital region (e.g. R. articeps: NHMUK R1236 , SHYMS G-132/1982).
Although the medial surface of the posterior process of the jugal is covered with sediment, it seems highly likely that the jugal−quadratojugal bar was incomplete, as in H. browni ( Dilkes, 1995) , M. browni ( Dilkes, 1998) , and R. articeps (NHMUK R1236) . The dorsal process of the jugal is elongate, and forms the entire anterior margin of the infratemporal fenestra, tapering to a very slender strip of bone at its dorsal tip ( Fig. 4C View Figure 4 : dp.j). By contrast, the dorsal process of the jugal is proportionately shorter in H. browni ( Dilkes, 1995) , M. browni ( Dilkes, 1998) , R. articeps , and R. brodiei ( Benton, 1990) . Moreover, in SAM-PK-K10159 the postorbital bar is formed along its entire length by an articulation between the dorsal process of the jugal and the elongate ventral process of the postorbital. This differs from the condition in B. sidensis ( Langer et al., 2010) , S. stockleyi ( Huene, 1938) , and hyperodapedontines ( Chatterjee, 1974; Benton, 1983a; Langer & Schultz, 2000; Whatley, 2005; Montefeltro et al., 2010), in which the bar is formed mostly by the dorsal process of the jugal. The very elongate dorsal process combined with the presence of a well-developed ventral process of the postorbital represents an autapomorphy of E. wolvaardti . The anterior margin of the dorsal process of the jugal contacts the posterior margin of the ventral process of the postorbital, differing from the condition in T. sulcognathus and Hyperodapedon spp. in which the ventral process of the postorbital fits into a slot in the dorsal margin of the dorsal process of the jugal ( Whatley, 2005; Montefeltro et al., 2010).
Prefrontal/lacrimal
The anterior border of the orbit is delimited by the mould of the medial projections of the ventral processes of the prefrontal and possibly lacrimal. This mould shows that the anterior wall of the orbit was well extended medially, as also occurs in R. brodiei (WARMS Gz6097). There is a nearly horizontal groove that is preserved as a natural mould situated anteroventral to the orbit, and which would most likely have been placed within the lacrimal. This groove appears to have exited on the anterior margin of the orbit, in the same position as the pair of foramina that are present for the exit of the naso-lacrimal duct in the lacrimal of R. brodiei (WARMS Gz6097).
Frontal
The frontal is completely missing, but the natural mould of its ventral surface is preserved ( Fig. 3A, C View Figure 3 ). This mould shows the morphology of the olfactory tract and, more anteriorly, the transversely expanded olfactory bulbs. The morphology of the olfactory apparatus resembles that of other basal archosauromorphs (e.g. Tasmaniosaurus triassicus: Ezcurra, 2014 ).
Postorbital
The postorbital is a triradiate bone ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : po). The posterior process is elongate and extends posteriorly, reaching a point level with the posterior border of the infratemporal fenestra ( Fig. 4C View Figure 4 : pp.po), as also occurs in all rhynchosaurids (e.g. R. articeps: NHMUK R1237 , SHYMS G-132/1982; B. sidensis: Langer et al., 2010 ; T. sulcognathus: Montefeltro et al., 2010 ; Hyperodapedon spp. : Chatterjee, 1974; Benton, 1983a; Langer & Schultz, 2000). Although not complete on either side, its posterior termination was gently rounded, as shown by the articular surface preserved on each squamosal. By contrast, in M. browni (SAM-PK-6536; Dilkes, 1998) and H. browni ( Dilkes, 1995) the posterior process of the postorbital is proportionally shorter, ending anterior to the posterior border of the infratemporal fenestra, and tapers to a point rather than being broadly rounded. The orbital margin of the postorbital of SAM-PK- K10159 View Materials is thickened ( Fig. 4C View Figure 4 : t.po), and posterior to this thickening the lateral surface of the bone is gently concave.
The ascending process of the postorbital forms a broad transverse suture with the postfrontal. The contacts in the postorbital−postfrontal−parietal region are not clear, but the ascending process appears to continue medially as a broad process that forms the entire anterior margin of the supratemporal fenestra, as in M. browni (SAM-PK-6536; Dilkes, 1998), H. browni ( Dilkes, 1995) , R. articeps (SHYMS G-132/1982, Benton, 1990), and S. stockleyi ( Montefeltro et al., 2010) .
Postfrontal
The lateral part of the postfrontal is preserved on the left side of the skull, contributing to the posterodorsal part of the orbital margin ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : pf). The postfrontal appears to be excluded from the anterior border of the supratemporal fenestra.
Parietal
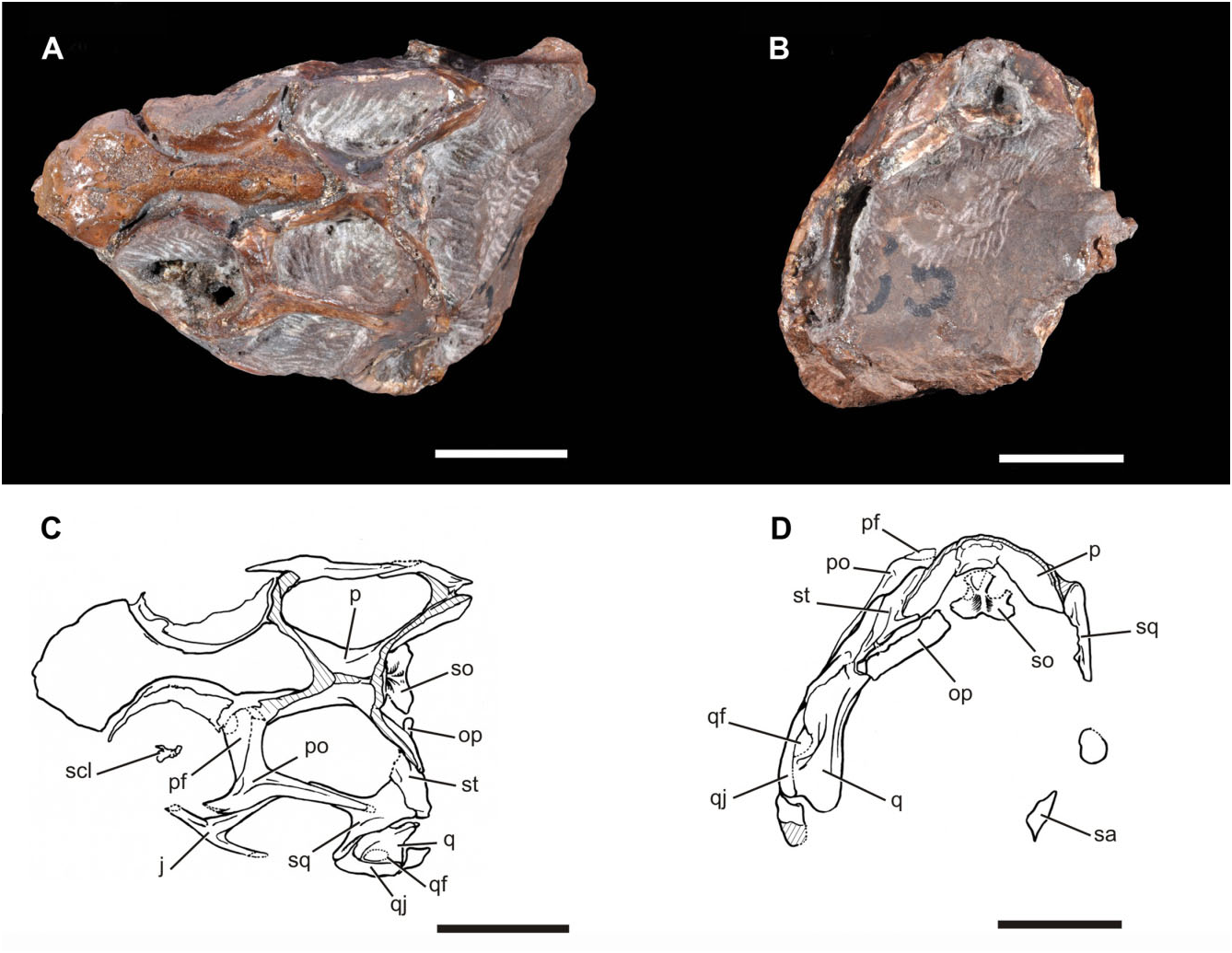
The parietals are fused to one another, without any trace of a median suture ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : p), resembling the condition present in other rhynchosaurs ( Dilkes, 1998). The anterior ends of the parietals are damaged, and the presence or absence of a pineal foramen cannot be determined. A sagittal crest is present along the full length of the midline of the parietals as preserved ( Fig. 4E View Figure 4 : sc). This differs from the condition in M. browni (SAM-PK-6536; Dilkes, 1998), in which the sagittal midline bears a concavity that is bounded laterally by ridges that define the supratemporal fenestrae, but resembles the condition in H. browni ( Dilkes, 1995) and rhynchosaurids ( Benton, 1983a, 1990; Langer et al., 2010; Montefeltro et al., 2010). As preserved, the posterolateral processes of the parietals are directed posterolaterally in dorsal view. The posterolateral orientation is similar to the condition in M. browni (SAM- PK-6536), H. browni (SAM-PK-5885), R. articeps (SHYMS G-132/1982), and S. stockleyi ( Montefeltro et al., 2010) , but differs from the laterally orientated processes in B. sidensis (BRSUG 27200), I. genovefae ( Whatley, 2005) , T. sulcognathus (UFRGS-PV-0232T, UFRGS-PV-0298), and Hyperodapedon spp. (e.g. H. huenei : UFRGS-PV-0132T; H. mariensis : FZB-PV- 1867, Montefeltro et al., 2010). In addition, the distal tips of the posterolateral processes do not curve anteriorly as occurs in S. stockleyi ( Montefeltro et al., 2010) and the ‘Mariante rhynchosaur’ (UFRGS-PV-0168T).
In posterior view the posterolateral processes are ventrally deflected at an angle of nearly 40° from the horizontal ( Fig. 3B, D View Figure 3 ). This ventral deflection is a unique condition amongst rhynchosaurs in which the posterolateral processes are typically directed nearly directly laterally in posterior view. This may represent an autapomorphy of E. wolvaardti , but this is difficult to confirm given the post-mortem deformation that has affected the skull (including the dorsal displacement of the parietals relative to the supraoccipital), and this character is not included in the diagnosis. The posterolateral processes of the parietals are dorsoventrally deep, but taper at their distal ends where they fit into a notch within the supratemporal. As in all other rhynchosaurs, there is no postparietal bone.
Supratemporal
The supratemporal is well preserved on the left side and lies between the posterolateral process of the parietal and the medial surface of the squamosal ( Figs 2 View Figure 2 , 3 View Figure 3 : st). This condition is different from T. sulcognathus and Hyperodapedon spp. , in which this bone is absent ( Benton, 1983a; Langer & Schultz, 2000; Langer et al., 2010; Montefeltro et al., 2010). The supratemporal in SAM-PK-K10159 is transversely broad in occipital view but contrasting with this species it also forms a small part of the posterior margin of the supratemporal fenestra as in M. browni (SAM-PK-6536). Posteriorly the supratemporal contacts the dorsal edge of the tip of the paroccipital process, and forms most of the dorsal margin of the post-temporal fenestra. This condition differs from that present in B. sidensis (BRSUG 27200; Langer et al., 2010), in which this bone lacks a ventromedial process and, as a result, is limited to a more lateral portion of the margin of the fenestra.
Squamosal
The squamosal has four distinct processes ( Figs 2 View Figure 2 , 3 View Figure 3 : sq). The anterior process is broadly overlapped by the extensive posterior process of the postorbital. The anterior process forms only small portions of the lateral border of the supratemporal fenestra and the dorsal border of the infratemporal fenestra. The ventral process is proportionally elongate, extending for more than 75% of the height of the infratemporal fenestra ( Fig. 4C View Figure 4 : vp.sq). This is similar to the condition in M. browni (SAM-PK-6536; Dilkes, 1998), F. spenceri ( Benton, 1990) , and T. sulcognathus (UFRGS-PV-0232T, Montefeltro et al., 2010), but differs from the abbreviated process that is present in H. browni ( Dilkes, 1995) and R. articeps (SHYMS G-132/1982). Additionally, the narrow anteroposterior width of the ventral process is similar to the condition in M. browni ( Dilkes, 1998) , H. browni ( Dilkes, 1995) , R. articeps (SHYMS G-132/ 1982), S. stockleyi ( Montefeltro et al., 2010) , and F. spenceri ( Benton, 1990) , but differs from the wide ventral process of the squamosal that is present in the hyperodapedontines I. genovefae ( Whatley, 2005) , T. sulcognathus ( Montefeltro et al., 2010) , and Hyperodapedon spp. ( Chatterjee, 1974; Benton, 1983a; Langer & Schultz, 2000). The medial process of the squamosal contacts the supratemporal, but it is unclear whether it contacted the parietal. Finally, there is a short posteroventral process that extends between the supratemporal and the head of the quadrate, reaching the paroccipital process. The lateral surface of the squamosal possesses several small subcircular pits.
Quadratojugal
The quadratojugal is a strap-like, slightly posteriorly bowed bone in lateral view ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : qj), resembling that of M. browni (SAM-PK-6536; Dilkes, 1998). The bone is best preserved on the left side, and is damaged slightly at its very anteroventral margin. A short, rounded anterior process of the quadratojugal appears to have been present ( Fig. 4 View Figure 4 : ap.qj), similar to the condition in R. articeps (SHYMS G-132/1982), but differing from the condition in M. browni in which an anterior process is absent ( Dilkes, 1998). The quadratojugal terminates posteriorly adjacent to the quadrate condyles. It forms the lateral margin of an oval quadrate foramen, shared with the quadrate. The size of the quadrate foramen is similar to that of M. browni (SAM- PK-6536) and R. articeps (SHYMS 3), but differs from the more reduced aperture that is present in F. spenceri ( Benton, 1990) , T. sulcognathus ( Montefeltro et al., 2010) ,
and a number of South American specimens of Hyperodapedon spp. (PVSJ 680, UFRGS-PV-0408T, UFRGS-PV-0149T).
Quadrate
The quadrate is strongly bowed along its length, with a concave posterior margin in lateral view ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 : q). Ventrally, the articular surface for the mandible is divided into two strongly convex condyles. The medial condyle is moderately better developed transversely and extends further ventral than the lateral condyle, resembling the condition present in M. browni (SAM- PK-6536), but differing from the condition in rhynchosaurids in which the condyles are equally well developed (e.g. R. articeps, NHMUK R1236 ; B. sidensis, BRSUG 27200; T. sulcognathus UFRGS-PV-0232). The lateral flange of the quadrate forms the dorsal margin of the quadrate foramen and overlaps the posterior margin of the ventral process of the squamosal ( Figs 3 View Figure 3 , 4 View Figure 4 : lf.q, qf).
Occiput
The supraoccipital is plate-like ( Fig. 3 View Figure 3 : so) and similar to that of M. browni ( Dilkes, 1998) and H. browni ( Dilkes, 1995) , but differs from the pillar-like supraoccipital that is present in Rhynchosauridae (e.g. R. articeps, Benton, 1990 ; B. sidensis, Langer et al., 2010 ; T. sulcognathus, Montefeltro et al., 2010 ). The supraoccipital has a very low midline ridge. The paroccipital process is strap-like, and as preserved is posterolaterally orientated, but appears to have been displaced from its articulation with the supraoccipital. Its ventral margin is straight as in M. browni , T. sulcognathus ( Montefeltro et al., 2010) , and some specimens of Hyperodapedon (e.g. UFRGS-PV-0149T), differing from the convex ventral margin that is present in R. articeps (SHYMS G-132/1982), S. stockleyi ( Huene, 1938) , the ‘Mariante rhynchosaur’ (UFRGS-PV- 0168T), B. sidensis (BRSUG 27200), and H. huenei (UFRGS-0132). The paroccipital process forms the ventral margin of the slit-like post-temporal fenestra.
Lower jaw, general
The dentary, surangular, angular, and articular are preserved ( Fig. 2 View Figure 2 ). The mandibular anatomy is generally very similar to that of M. browni (SAM-PK-6536; Dilkes, 1998). There is no mandibular fenestra, as in all rhynchosaurs. The maximum dorsoventral height of the mandible is less than a quarter of the mandibular length, differing from hyperodapedontines in which the mandible is higher than a quarter of the mandibular length ( Benton, 1990; Dilkes, 1998; Whatley, 2005; Montefeltro et al., 2010).
Dentary
The dentary is incomplete anteriorly on both sides, but a natural mould on the right side indicates that the bone tapered in dorsoventral height towards its anterior end, and would probably have ended approximately level with the anterior margin of the maxilla ( Fig. 2 View Figure 2 : d), as in M. browni (SAM-PK-6536; Dilkes, 1998). Our estimation suggests that the dentary comprises less than half of the total length of the mandible, similar to the condition in M. browni and S. stockleyi ( Benton, 1984, 1990; Dilkes, 1998). Natural moulds indicate that, as in the maxilla, the dentary was transversely expanded, and further show that multiple tooth rows were present, and that teeth were present on the lingual as well as occlusal surfaces. Posteriorly, the dentary has an elongate posterodorsal process that overlaps the lateral surface of the surangular.
Surangular
The surangular is a large element that dominates the mandible in lateral view, extending for more than 70% of the mandibular length, and extending posterior to the glenoid fossa, forming the lateral surface of the retroarticular process ( Figs 2 View Figure 2 , 3 View Figure 3 : sa). The surangular has strongly convex dorsal and ventral margins, and a gently concave lateral surface. The lateral border of the glenoid fossa (which is not exposed) forms a low tuberosity. Immediately below this tuberosity there is a small posterior surangular foramen ( Fig. 2 View Figure 2 : sf).
Angular
The angular is exposed as a narrow strip of bone ventral to the surangular ( Fig. 2 View Figure 2 : a).
Articular
The articular is poorly preserved, but it can be determined that it contributed to a well posteriorly developed and upturned retroarticular process ( Fig. 2 View Figure 2 : ar), resembling the condition in H. browni ( Dilkes, 1995) .
Hyoid apparatus
The right ceratobranchial of the hyoid apparatus is preserved, and is a slender, rod-like element ( Fig. 2 View Figure 2 : hy). As preserved, it extends posteriorly beyond the level of the craniomandibular joint, and is missing its posterior end.
Dentition
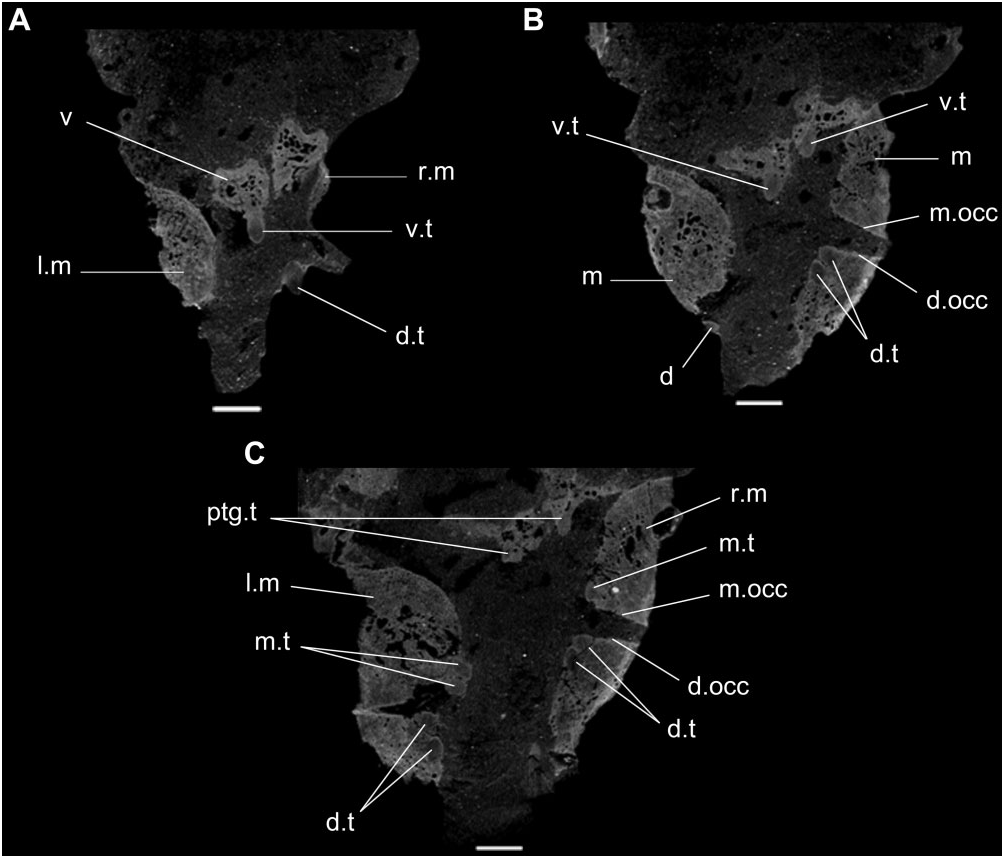
Micro-CT data demonstrate that the maxillae and dentaries are transversely expanded, forming dental plates with multiple rows of conical teeth, with teeth present both on occlusal and lingual surfaces of the bones ( Fig. 5 View Figure 5 : d.occ, m.occ). This condition resembles the jaw apparatus of H. browni ( Dilkes, 1995) . The toothbearing surfaces of the maxilla and dentary are divided into a nearly horizontally orientated occlusal surface, and a nearly vertical, medially facing surface, which bears the lingual teeth ( Fig. 5 View Figure 5 : d.t, m.t). This condition is different from the lingual margin of the tooth plates of H. browni , in which the occlusal surface more gradually merges into the lingual surface ( Dilkes, 1995). As in H. browni ( Dilkes, 1995) , there is no occlusal groove, but in some places the occlusal surface of the maxilla appears to be gently concave. The presence of dental plates in both the maxilla and dentary in SAM- PK-K10159 is different from the groove and blade jaw apparatus of all rhynchosaurids, including the fragmentary Middle Triassic form Ammorhynchus navajoi and the ‘Chañares form’ ( Benton, 1983a, 1984, 1990; Langer et al., 2000; Nesbitt & Whatley, 2004; Montefeltro et al., 2010; Ezcurra et al., 2014).
The most clearly distinguishable and most complete teeth are the most lingual. These teeth are clearly set within shallow sockets and appear to be fused to the surrounding bone, although they are still distinguishable. More labially positioned teeth, and teeth on the occlusal surface, are more completely fused and difficult to distinguish from the surrounding bone, with those on the occlusal surface probably worn completely level with the bone.
As in M. browni there is clearly a single row of tightly packed teeth on the vomer ( Fig. 5 View Figure 5 : v.t), and slightly smaller teeth are also present along the thickened medial parts of the pterygoids ( Fig. 5 View Figure 5 : ptg.t). It is unclear whether or not palatine teeth are present. The palatal teeth of the vomer and pterygoid are similar in size to the lingual teeth present on the maxilla and dentary.
Possible tibia and fibula
There is a partial long bone preserved adjacent to the posterior end of the right side of the skull that partially obscures the craniomandibular joint ( Fig. 2A, C View Figure 2 : lb). Next to it, there is a natural mould of a long bone shaft that possesses the same longitudinal orientation ( Fig. 2A, C View Figure 2 : lbm). The size, curvature of the shaft, and morphology of the preserved articular end suggest that the preserved long bone is the distal half of a fibula and the natural mould next to it may be of a tibia. The probable fibula is bowed along its length and the distal articular surface is subcircular.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Genus |