
Neoseiulus californicus (McGregor, 1954) sensu Athias-Henriot, 1977
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4500.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:16A34E21-D55D-40E9-BF2D-43D3BD8A6AF2 |
|
persistent identifier |
https://treatment.plazi.org/id/03E987BF-FFD9-FFE7-FF44-F8AEFBA5E32A |
|
treatment provided by |
Felipe |
|
scientific name |
Neoseiulus californicus |
| status |
|
McGregor’s N. californicus View in CoL and N. mungeri match N. barkeri , not N. californicus sensu Athias-Henriot
Our study of the types of N. californicus and N. mungeri indicates that they are essentially identical to the male and female of N. barkeri , respectively. Accordingly, the N. californicus and N. mungeri types are clearly distinct from the male and female of N. californicus sensu Athias-Henriot (1977) . This conclusion is strongly supported by the following set of characters that are shared by N. californicus (holotype), N. mungeri (lectotype, paralectotype), and N. barkeri :
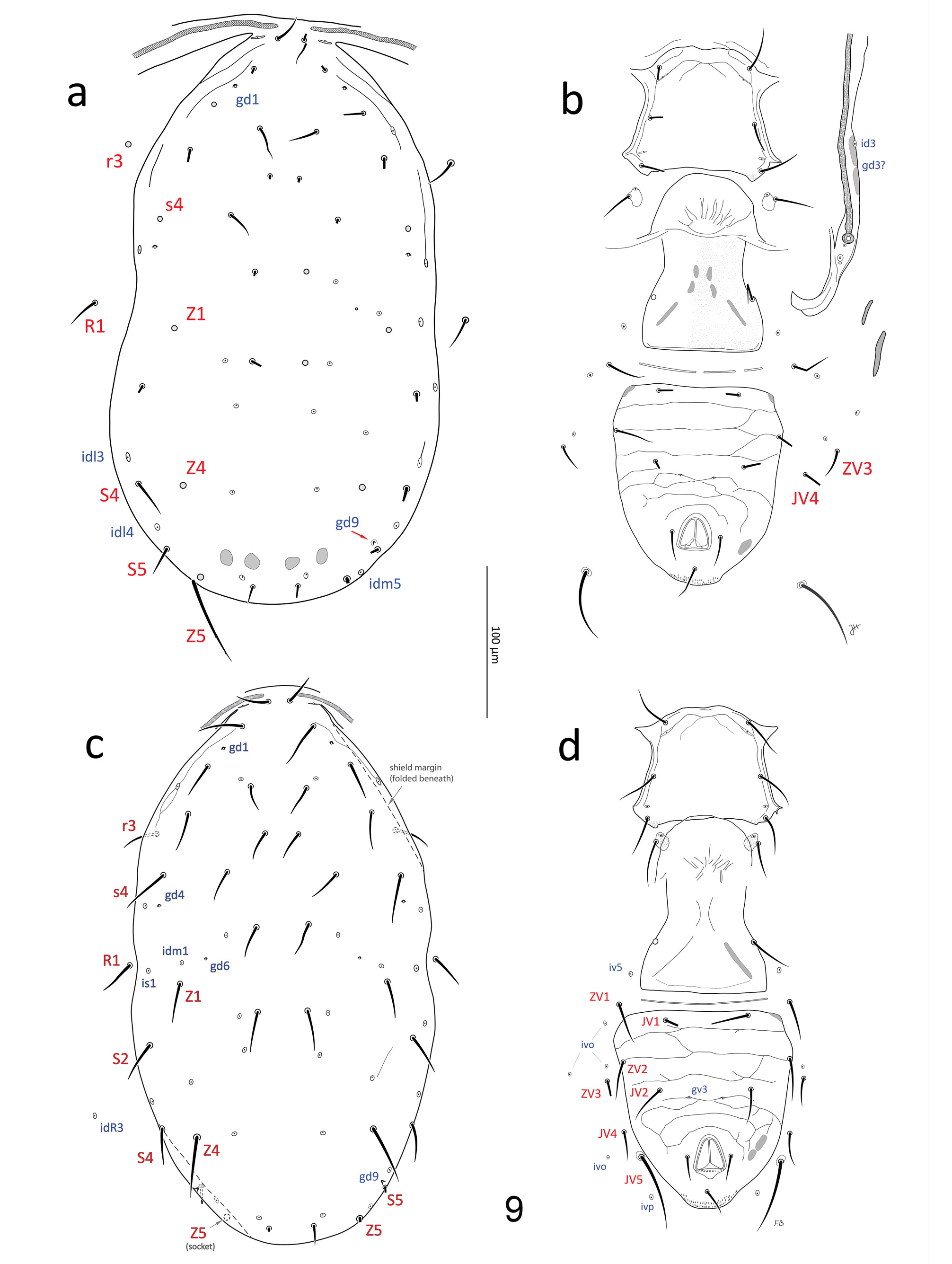
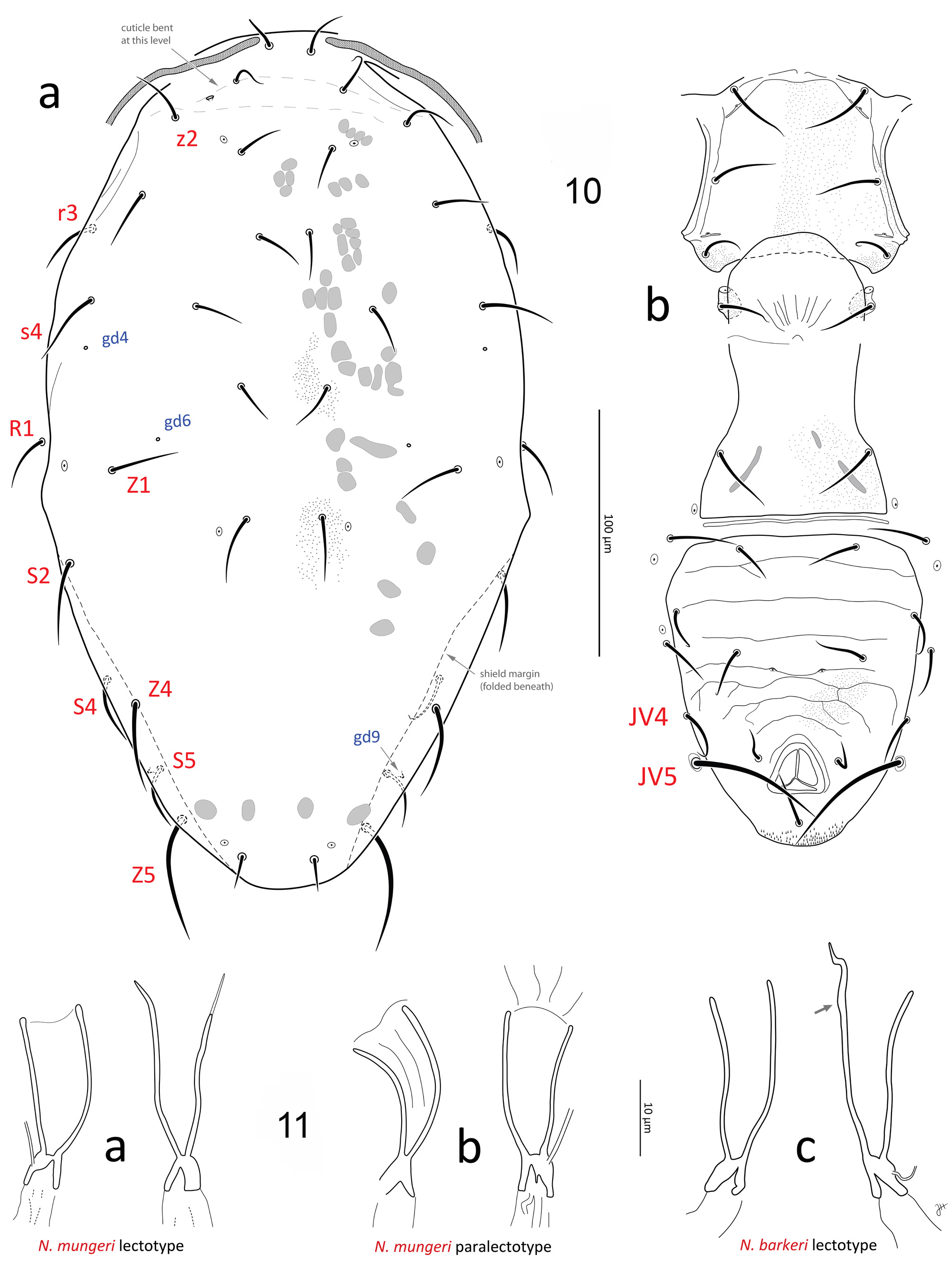
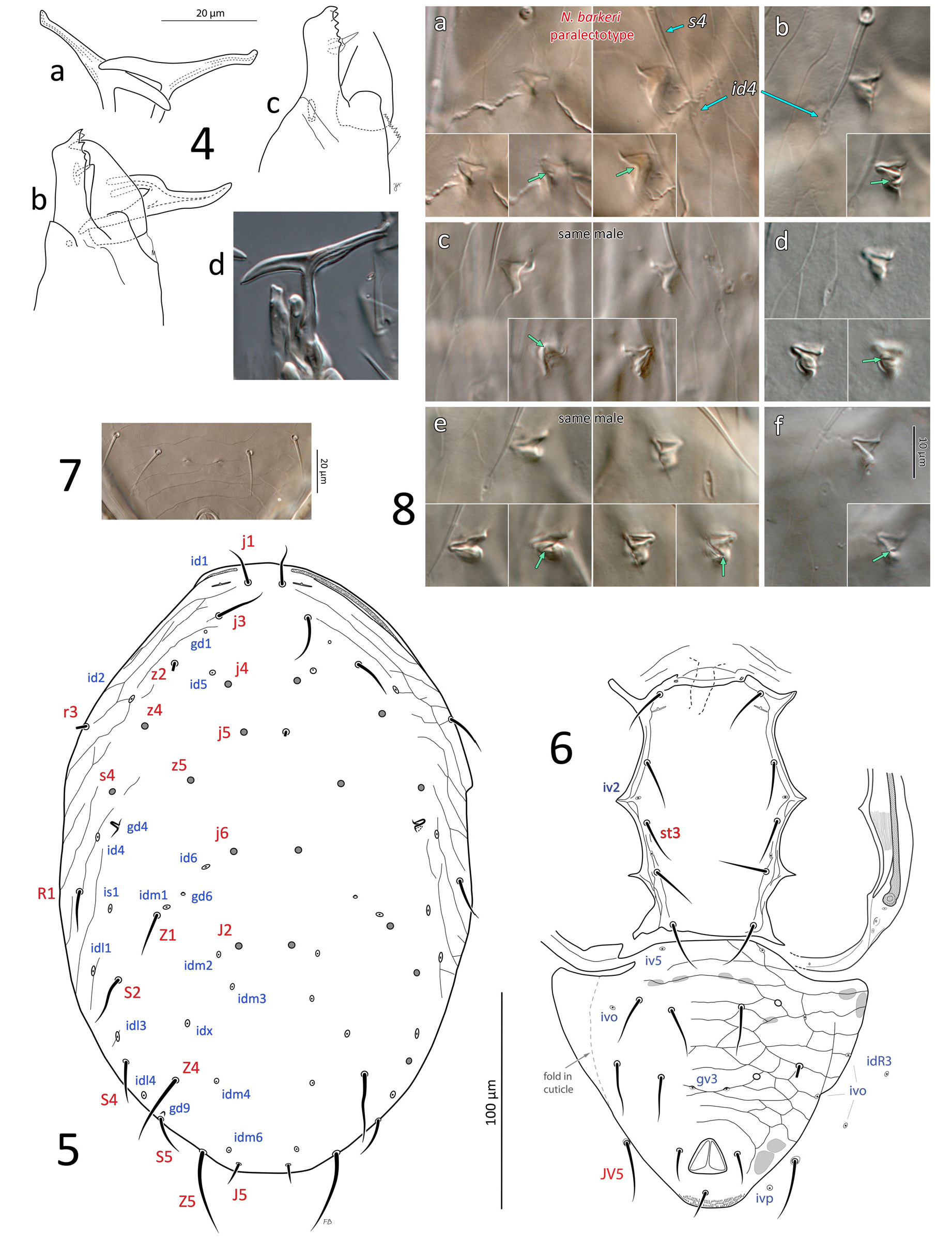
(1) Extensive morphometrics (over 47 characters; Table 2 for N. barkeri and conspecific populations; Table 3 for Nc -AH), including the length of most idiosomal setae. Nc -McG ( Fig. 3a, c View FIGURE 3 ) and N. mungeri ( Fig. 9a–b View FIGURE 9 ), show essentially all compatible morphometrics with male and female N. barkeri (slide lots #3–18, Table 1), including the N. barkeri lectotype ( Fig. 10a–b View FIGURES 10–11 ) and a paralectotype ( Fig. 3b, d View FIGURE 3 ; lot #7), and males and females from a culture (lot #16). Although many dorsal setae of the two female types of N. mungeri are missing, the remaining ten setae have lengths compatible with those of N. barkeri (Table 2); furthermore, the five additional females from California with similar collection data to that of N. mungeri (lots #3–6; Fig. 9c–d View FIGURE 9 ), variously identified as “ T. californicus ”, “ mungeri ” or “? mungeri ” by McGregor (or P. Garman), also have dorsal setae of compatible lengths to those of the N. barkeri lectotype and other N. barkeri females examined. The additional male from California (lot #3; Figs 5–6 View FIGURES 4–8 ) with similar collection data to the type of N. californicus , also has morphometrics that match those of N. barkeri paralectotype (male).
(2) The spermatheca of the N. mungeri type falls well within the range of intraspecific variation of shapes that we observed for N. barkeri specimens, including N. barkeri lectotype ( Figs 11a–b View FIGURES 10–11 vs 11c, 17a–y; see Remarks section p. 30 for more details), and closely match the spermathecae illustrated for N. barkeri in various descriptions ( Athias-Henriot, 1961, 1966; Ehara, 1972; Ragusa & Athias-Henriot, 1983; Tuovinen, 1993; Papadoulis et al., 1996; Swirski et al., 1998; Chant & McMurtry, 2003; Kolodochka, 2006; Amano et al., 2011).
(3) The T-shaped spermatodactyl of the N. californicus type matches the characteristic shape seen in male N. barkeri (and of other members of the N. barkeri species group, e.g. N. tareensis (Schicha) ( Schicha, 1987) , N. usitatus (Van Der Merwe) ( Ragusa & Athias-Henriot, 1983)) , with distinctly elongate foot, including elongate heel with rounded tip and toe portion slightly recurved apically. Lengths of spermatodactyl foot, and toe and heel portions identical to N. barkeri paralectotype and other male N. barkeri ( Fig. 4a, b, d View FIGURES 4–8 ).
(4) Shield ornamentation, including (a) mostly smooth dorsal shield, except for lineation-reticulation covering the anterior marginal region laterad of setae j1, j3, z2, z4, s4 ( Figs 9a, c View FIGURE 9 , 10a View FIGURES 10–11 , 12 View FIGURES 12–15 ), along with areas surrounding R1 and laterad of S 2 in N. californicus type ( Fig. 3a View FIGURE 3 ; but see other male from California with almost identical collection data, Fig. 5 View FIGURES 4–8 , for clearer reticulation) and male N. barkeri (paralectotype, Fig. 3b View FIGURE 3 ); (b) smooth sternal and sternogenital shields, except few lineae along lateral and anterior margins ( Figs 3c–d View FIGURE 3 , 6 View FIGURES 4–8 , 9b, d View FIGURE 9 , 10b View FIGURES 10–11 , 13 View FIGURES 12–15 ).
(5) Ventrianal shields with gland openings gv3 small, slightly crescent-shaped, and separated by a distance 0.37– 0.44 x distance between JV2 setae; ventrianal shield of N. mungeri type and female N. barkeri (including lectotype) with truncate anterolateral corners, and weak ornamentation of mostly transversal lineae ( Figs 9b, d View FIGURE 9 , 10b View FIGURES 10–11 , 13–15 View FIGURES 12–15 ).
Remarkably, McGregor’s (1954) descriptions not only show that Nc -McG and N. mungeri are distinct from Nc -AH (see Griffiths, 2015), they also show that Nc -McG and N. mungeri are morphologically similar to N. barkeri , despite that the illustrations lack many critical details (particularly the spermatheca and spermatodactyl not being included). Before we found the types, we had already reached the conclusion that, in all probability, McGregor’s species were the same species as N. barkeri . This hypothesis was prompted by our earlier finding of female (5) and male (1) specimens with similar collection data (lots #3–6) to the Nc -McG and N. mungeri types, variously identified by McGregor as ‘ californicus ’, ‘ mungeri ’ and ‘? mungeri ’, but which we identified as N. barkeri . Using the original descriptions of Nc -McG and N. mungeri , we found a series of character states that are discernible in the two descriptions, the types of Nc -McG and N. mungeri , and also N. barkeri . Importantly, these features are distinct from those of Nc -AH, and are presented below (compare also Figs for N. barkeri (3–16) vs. Nc-AH (29–41)).
(1) Male ventrianal shield with four pairs of pre-anal setae [three pairs of setae (ZV1 absent) in Nc -AH] ( Griffiths, 2015).
(2) Dorsal gland opening gd4 conspicuous, posterior to seta s4, far from shield margin, enlarged in male [gd4 absent in Nc -AH; instead, male Nc -AH has gd3 on the margin of dorsal shield, posterolateral to s4; female has gd3 on peritrematal shield] (see comment about gd3 and gd4 near the end of Material & Methods, p. 6).
(3) Setae j1 well separated, distance between j1–j1 subequal to length of j1 [j1 sockets almost touching each other in Nc -AH, distance j1–j1 approximately one third the length of j1].
(4) Dorsal gland opening gd9 close to, almost contiguous with, seta S5 [gd9 distant from S 5 in Nc -AH].
(5) Setae Z4–Z5 relatively short; Z5 0.129 (±0.006) x length of dorsal shield in male specimens, and 0.146 (±0.011) x length of dorsal shield in females [Z4–Z5 longer in Nc -AH, with Z5 0.184 (±0.013) x length of dorsal shield in male, and 0.189 (±0.008) in female].
(6) Sternogenital/sternal shield smooth, except for marginal lineae [mostly reticulate in Nc -AH].
(7) Female ventrianal shield with lateral margins essentially convex [with a slight waist near level of setae JV 2 in Nc-AH].
(8) Female dorsal poroids is1 and idl1 near but not on shield margin [Nc -AH is1 and idl1 on shield margin]; is1 also tends to be positioned more posteriorly in N. barkeri than in Nc -AH, in relation to an imaginary line drawn between setae Z1 and R1.
(9) Female movable cheliceral digit with a single small tooth [three teeth in Nc -AH].
(10) Male dorsal setae R1 inserted on shield distant to shield margin [R1 inserted on shield margin in Nc -AH].
(11) Dorsal setae Z1 and S2 inserted quite distant to shield margin (this is especially clear for N. mungeri and N. barkeri female vs Nc -AH female).
(12) Dorsal gland opening gd1 approximately aligned with setae j3 and z2 (note that McGregor had interpreted and illustrated gd1 as a minute seta) [gd1 situated slightly to moderately anterolaterad of ‘ j3-z2 line’ in Nc -AH].
(13) Postanal seta approximately (0.9–1.2x) as long as para-anal setae, and shorter than pre-anal setae [Nc -AH postanal seta approximately 1.2–1.6x as long as para-anals, and as long as or longer than pre-anal setae].
(14) Sternogenital shield clearly broader at level between coxae II–III than at level between coxae III–IV, and poroid iv2 positioned lateral to seta st3 (however, in female, iv2 is positioned at same (longitudinal) level of st2) [Nc -AH sternogenital shield only slightly broader at level between coxae II–III, and iv2 mesal to seta st3]. Interestingly, two previous authors had also observed similarities between Nc -McG, N. mungeri and N. barkeri ,
in addition to Xu et al. (2013): (1) Athias-Henriot (1959) identified female specimens as N. mungeri to later re-
identify them as Amblyseius barkeri (Hughes) ( Athias-Henriot, 1961: 440) ; (2) Chant (1959) synonymised N.
californicus and N. mungeri under N. marinus based on both McGregor’s types, and on various male and female
specimens that were incorrectly identified as N. marinus . We use the word ‘incorrectly’ because based on our direct
observations, the ‘ N. marinus ’ specimens examined by Chant actually represent N. barkeri and a closely related
species in the N. barkeri group (slide lots #3, 18, 19) (see p. 487 for further explanation). Therefore, ipso facto,
Chant (1959) also observed similarities between Nc -McG, N. mungeri and N. barkeri .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |