
Ziphirostrum marginatum du Bus, 1868
|
publication ID |
https://doi.org/ 10.5281/zenodo.5376445 |
|
persistent identifier |
https://treatment.plazi.org/id/03FED57F-FFEB-9F64-819F-FD7AFE48FC8C |
|
treatment provided by |
Marcus |
|
scientific name |
Ziphirostrum marginatum du Bus, 1868 |
| status |
|
Ziphirostrum marginatum du Bus, 1868
Ziphirostrum marginatum du Bus, 1868: 624 .
Ziphirostrum laevigatum du Bus, 1868: 624 . — Van Beneden & Gervais 1880: pl. 27bis, fig. 5.
Ziphirostrum gracile du Bus, 1868: 625 .
Mioziphius belgicus Abel, 1905: 99 , partim, fig. 17, 101, fig. 18.
HOLOTYPE. — IRSNB 3783 View Materials - M.1878, a partial rostrum with a part of the right premaxillary sac fossa and the right supraorbital process (single specimen of Ziphirostrum marginatum sensu du Bus, 1868 ).
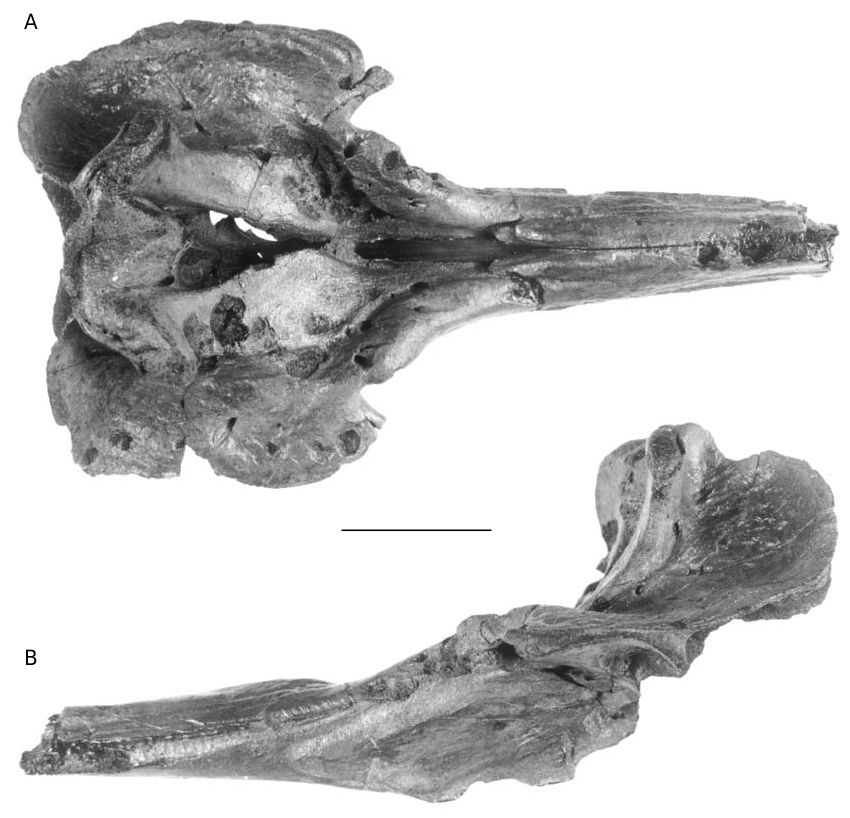
REFERRED SPECIMENS. — 16 additional specimens: IRSNB 3845- M.536, partial skull (identified by du Bus as “ Ziphirostrum hemixemii Van Ben. ” [label on the skull], described and figured as cotype of Mioziphius belgicus by Abel 1905: fig. 17); IRSNB 3847- M.537, partial skull (labelled “ Ziphirostrum hemixemii Van Ben. ” by du Bus, described and figured as cotype of Mioziphius belgicus by Abel 1905: fig. 18); IRSNB 3820, partial rostrum; IRSNB 3828, partial rostrum; IRSNB 3829, partial skull; IRSNB 3833, partial rostrum; IRSNB 3839, partial skull (labelled “ Ziphirostrum hemixemii Van Ben. ” by du Bus); IRSNB 3841, partial rostrum (labelled “ Ziphirostrum hemixemii Van Ben. ” by du Bus); IRSNB 3842, partial skull (labelled “ Ziphirostrum hemixemii Van Ben. ” by du Bus); IRSNB 3843- M.1876, partial skull; IRSNB 3844- M.1874, partial skull; IRSNB 3846- M.1875, partial skull (labelled “ Ziphirostrum hemixemii Van Ben. ” by du Bus); IRSNB 3827- M.1879, fragment of rostrum ( Ziphirostrum gracile sensu du Bus, 1868 ); IRSNB 3830- M.542, partial skull ( Ziphirostrum laevigatum sensu du Bus, 1868 , figured by Van Beneden & Gervais 1880: pl. 27bis, fig. 5); IRSNB IG.8638, partial skull (from Deurne, donated by Hasse, April 30, 1921); IRSNB M.1877, partial skull with four teeth (found by M. Bosselaers in April 2001, at Deurne, eastern suburbs of Antwerp, cited in Bosselaers et al. 2004).
TYPE HORIZON. — No information is available for the holotype. The skull IRSNB M.1877 was found in the Deurne Sands Member, Diest Formation, upper Miocene, middle to late Tortonian ( Louwye 2002).
TYPE LOCALITY. — Area of Antwerp, Belgium, exact locality uncertain.
EMENDED DIAGNOSIS. — The species differs from Ziphirostrum turniense in: a deeper, longer and wider prenarial basin, bordered by relatively thicker and higher strips of the maxillae posterolaterally curving towards the antorbital notches; a more posterior position of the top of the premaxillae on the rostrum, at one quarter of its total length. It differs from the morefragmentarily known Z. recurvus n. comb. in a relatively lower rostrum, wider than high in its anterior portion; the open mesorostral tunnel.
DESCRIPTION ( FIGS 2-10 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
General observations
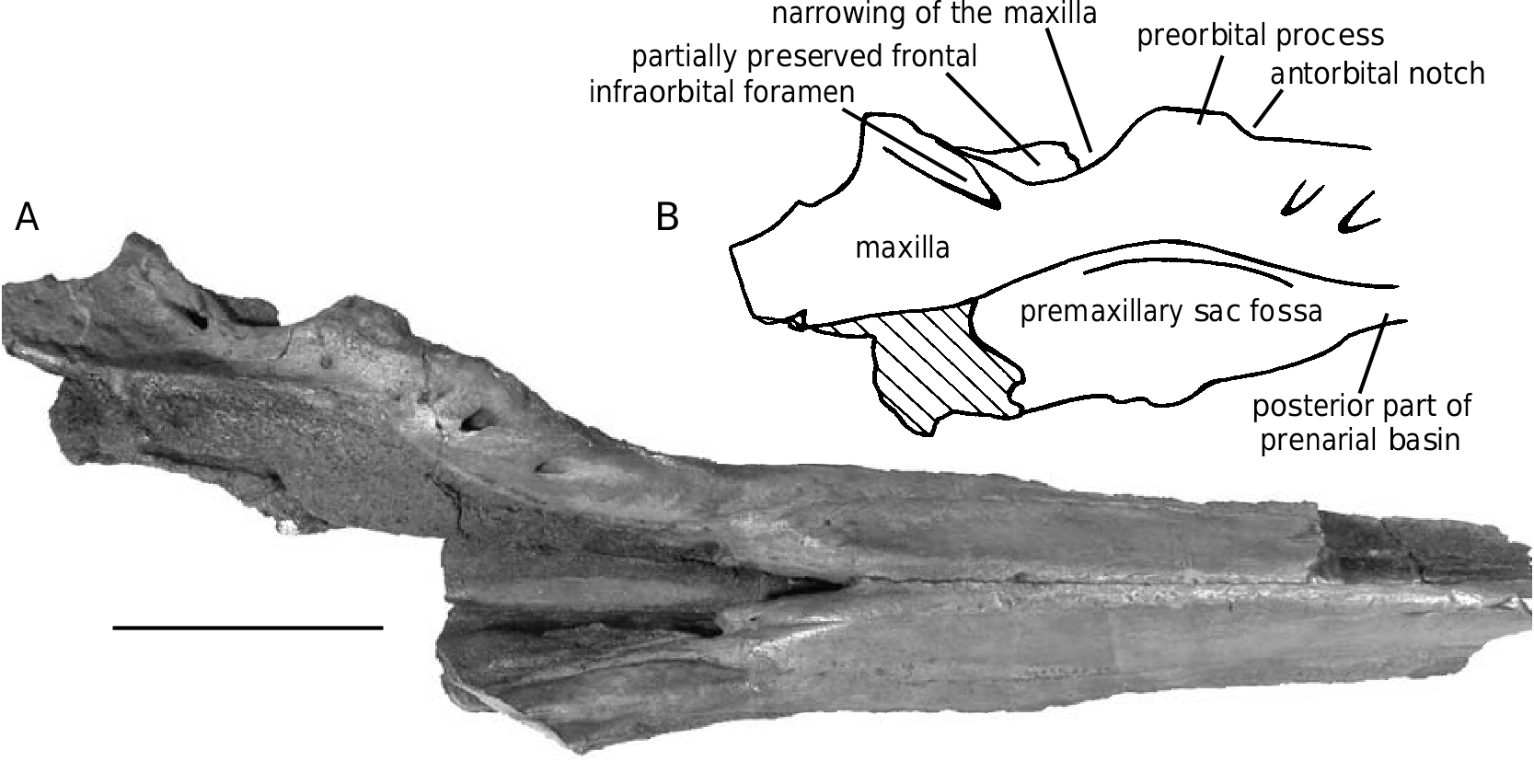
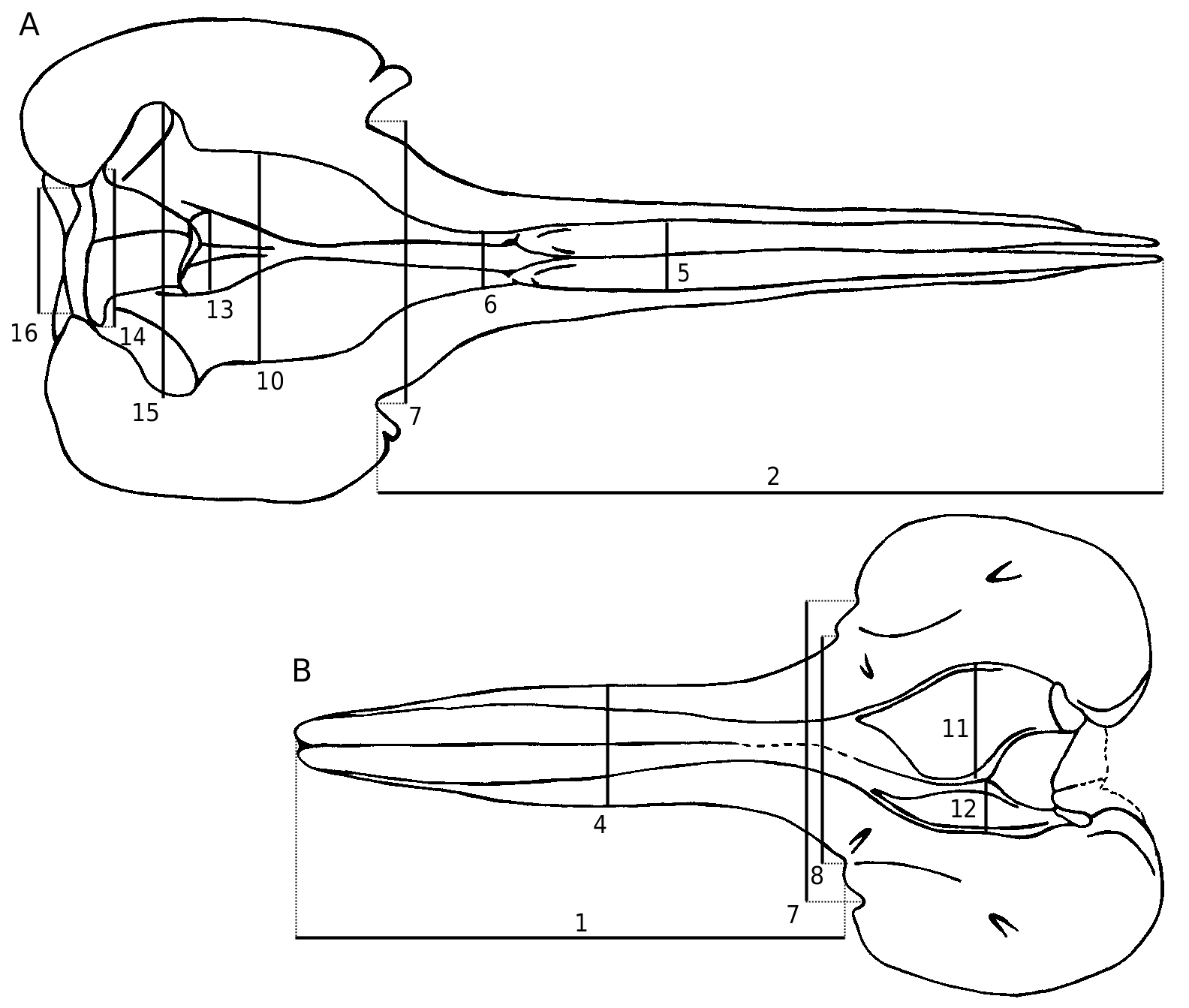
This species was previously only known by the rostrum and the anterior part of the cranium; the basicranium and supraoccipital are usually lacking, as well as the teeth and ear bones. No mandible can be referred to this species; the mandibles described by Abel (1905) were found isolated (see below). The preserved parts are always robust and heavy, particularly the pachyosteosclerotic rostrum with the mesorostral groove dorsally closed by the thickened premaxillae. The skull is medium-sized, with a supraorbital width (see Table 1, measurement 6) intermediate between the smaller Messapicetus and the larger Ziphius . The relatively long roughly cylindrical rostrum is shorter than in Messapicetus , but more elongated than in Ziphius , proportionally closer to Tasmacetus . A deep prenarial basin (sensu Heyning 1989a) precedes the asymmetrical premaxillary sac fossae. The vertex is wide and high.
Several specimens – IRSNB 3845-M.536, IRSNB 3844-M.1874, IRSNB 3829, IRSNB 3841, and IRSNB 3842 – show rounded postmortem excavations of the surface of the bone with a more or less regular outline. Those depressions have a variable diameter: the small ones reach only 7 to 9 mm, while larger ones, more ellipsoidal, have a maximum diameter reaching 19-23 mm – with the largest reaching 33 mm. The depth is also variable: the depressions are sometimes very shallow (less than 1 mm) but some are deeper than 10 mm. The most excavat- ed area of the skulls is the dorsal part of the maxillae and the premaxillae on the cranium, indicating that the large mass of facial muscles was already detached at the moment of the drillings. Some fossae also excavate the lateral and ventral surfaces of the rostrum. More than 25 fossae are counted on IRSNB 3845-M.536 ( Fig. 2 View FIG ). Those shallow holes may be aborted drillings made by bivalves. Those molluscans tried to excavate the bone, but its high density and hardness might have precluded them to finish their work. Among the bivalves recorded in the upper Miocene of the region of Antwerp, species that are known to drill in harden substrates are Hiatella arctica (Linnaeus, 1758) and Gastrochoena dubia (Pennant, 1777) (R. Marquet pers. comm. 2004). Other organisms that could be responsible of those holes are bone-eating osteopeltid limpets, or even echinoids (R. E. Fordyce pers. comm. 2005).
Premaxilla
The premaxillae are thick and dense on the rostrum, making it strong and heavy. They dorsally close most of the mesorostral groove. On the nearly complete rostrum IRSNB 3847-M.537, the premaxillae are closely applied for 310 of the 550 preserved mm, with a dorsal median suture distinct as a thin groove ( Fig. 5A View FIG ). They are separated on the apical 140 mm, with a progressive forward opening, and on the prenarial basin. When dislocated, the surface of suture between the two premaxillae on the rostrum is planar, without vascular grooves, indicating a close contact. A central tunnel between the vomer and the premaxillae, remnant of the mesorostral groove, is retained. The premaxillae occupy a large part of the dorsal surface of the rostrum apically; they widen posteriorly, reaching their maximum width at 200-205 mm of the antorbital notch. Then their width slightly decreases until the prenarial basin. Along their descent in the basin, the premaxillae abruptly narrow, and nearly disappear from the dorsal view, the left one more distinctly than the right one, at the level of the premaxillary foramen. Seen in lateral view, the rostrum is roughly rectilinear. The premaxilla appears at the anterior limit of the prenarial basin; it thickens until a maximum height reached 60-80 mm forwards. Then, it progressively lowers, with a second less pronounced prominence, and tapers towards the apex.
On the skull IRSNB 3847-M.537, with a long portion of the rostrum preserved, the poorly marked lateral suture between maxilla and premaxilla is visible until the anterior end of the maxilla, 510 mm anterior to the antorbital notch and 45 mm before the apex of the incomplete rostrum (probably lacking some centimetres).
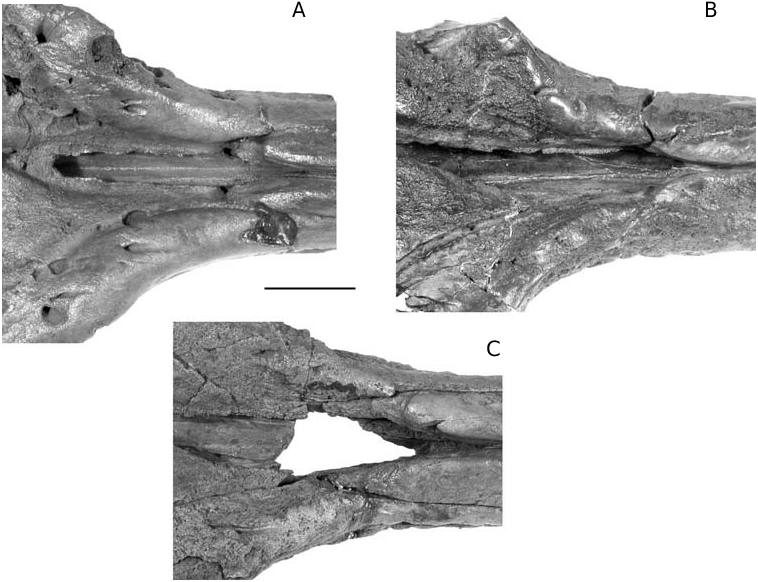
In Ziphirostrum marginatum , the prenarial basin is a deep and wide depression in the base of the rostrum, formed by nearly complete resorption of the premaxillae, and laterally limited by the maxillae. The basin extends from a level slightly anterior to the premaxillary foramina to the anterior of the premaxillary sac fossae ( Fig. 4A View FIG ). The floor of the basin is made of the vomer, which sends thin dorsomedial plates partially dorsally closing the mesorostral groove in the basin.
The premaxillary sac fossae are distinctly asymmetrical: the right one is longer and wider than the left. On IRSNB 3845-M.536, the right fossa reaches a maximum width of 68 mm (taken perpendicular to the longitudinal axis of the skull), and the left one 54 mm. The premaxillae nearly contact each other immediately anterior to the bony nares (6 mm of separation on IRSNB 3845- M.536). More anteriorly, they are joined on IRSNB 3845-M.536 by an irregular mass of bone, probably the mesethmoid, closing the mesorostral groove for a short distance. The posterodorsal elevation of the premaxilla is progressive, with an inclination of more or less 45° for the main part of the ascending process. At the level of the narrowest portion of the premaxilla, the slope distinctly increases; on the last centimetres, the flat anteriorly-facing surface of the bone becomes vertical, with a distinct twisting giving that surface a more medioanterior orientation. The wide and thick dorsal extremity of the premaxilla forms the transverse premaxillary crest, a lateral part of the vertex. The medial part of the crest is anterolaterally directed, while the anterolateral part is more laterally directed ( Fig. 5D View FIG ).
Maxilla
The prenarial basin is laterally margined by the thick, prominent and wide maxilla, sometimes slightly lower and flatter, rising towards the antorbital notch. This posterolaterally oriented thick strip of the maxilla at the base of the rostrum is pierced by a series of dorsal infraorbital foramina until the level of the antorbital notch. The maxilla reaches a maximum dorsal height just before the notch, with a variably acute dorsolateral edge (e.g., lower and more rounded on IRSNB 3845-M.536).
In lateral view, a longitudinal lateral groove appears below the maxilla-premaxilla suture 100- 120 mm anterior to the antorbital notch; it quickly widens forward up to c. 15 mm width. The bottom of this groove is punctuated by a series of regularly spaced shallow depressions 6-7 mm in diameter, separated by transverse bridges 3-4 mm long ( Figs 3B View FIG ; 4B View FIG ). These shallow vestigial alveoli could probably not effectively hold teeth; alternatively, perhaps teeth were mainly supported by the gum. The groove and alveoli disappear some centimetres before the anterior point of the maxilla, and do not mark the smooth lateral surface of the premaxilla. Forty-two alveoli are counted on the complete left maxilla of the skull IRSNB 3847-M.537.
The median ventral sutures are poorly discernable and the acute keel seen on the holotype medially to the anterior sinus fossa is at least partially made of the vomer. From 40 mm anteriorly to the palatines, the two maxillae are separated by a thin anteriorly widening ventral exposition of the vomer, for more than 190 mm on the skull IRSNB 3847 View Materials - M.537. The maxillae are pierced along their median suture by pairs of palatine foramina opening forwards (more than three pairs on the incomplete IRSNB 3845 View Materials - M.536). The most posterior pair is located along the median margins of the palatines ( Figs 4B View FIG ; 5C View FIG ) .
The antorbital notch is deep and wide. The anterior margin of the maxilla on the preorbital process curves anteroventrally around the jugal, forming a half cylinder anterolaterally directed, with a lateral extremity well demarcated from the preorbital process of the frontal. The thick strip of the maxilla medial to the antorbital notch extends for a short distance on the preorbital process, forming a shallow dome. The supraorbital process of the maxilla is pierced on IRSNB 3845-M.536 by one dorsal infraorbital foramen on the left, and one major and one smaller foramen on the right side. A plate of the maxilla totally covers the posterolateral edge of the transverse premaxillary crest, slightly overhanging the rest of the maxilla. The suture between this plate and the transverse premaxillary crest is pierced by a series of small vertical foramina (five major ones are distinct on both sides on IRSNB 3845- M.536, with several smaller ones).
Nasal
Between the premaxillary crests, the triangular nasals, longer than wide, form an anterior round- ed protuberance overhanging the bony nares; this prominent anterior angle reaches the anterior level of the premaxillary crests. The posterior margin of the nasals is rectilinear; their median suture is, anteriorly, distinctly deviated to the left side, so that it lies to the left relative to the median sutures of the rostrum and the inner sagittal crest of the cranium ( Fig. 4A View FIG ). The joined dorsal surfaces of the nasals are slightly medially depressed. Each nasal contacts the premaxillary crest only at the level of its posterolateral corner. In some specimens, the nasal sometimes contacts the median plate of the maxilla, preventing a suture between premaxilla and frontal, but the sutures are never clear on this area, and the character might even be variable on opposite sides of the same skull.
Mesethmoid
The dorsal margin of the posterior plate of the mesethmoid reaches a level 30 mm under the dorsal surface of the nasals. The triple point between nasal, premaxilla and mesethmoid is pierced on IRSNB 3845-M.536 by a vertical foramen, wider on the right side, probably corresponding to the additional exit from the infraorbital complex mentioned by Rommel (1990: 36) in Tursiops , and observed in several odontocetes, including extant ziphiids. This foramen is not always present (e.g., absent on IRSNB 3847- M.537). On IRSNB 3845-M.536, the mesethmoid is pierced by a terminal nerve foramen (see Rommel 1990 in Tursiops ), at mid-height and on the two sides of the low and wide keel. This moderately large foramen is laterally prolonged by a short groove, and exits on the inner anterior face of the cranium cavity at the suture between frontal and mesethmoid.
Frontal
The lateral margin of the supraorbital process is horizontal and thin, with a moderate preorbital thickening. The postorbital process is slender, probably originally more than 27 mm long. The frontals are poorly preserved on the vertex, forming a short strip along the posterior margin of the nasals and the posterior tip of the premaxillae. The right frontal of IRSNB 3847-M.537 is complete with a length of 20 mm (see Fig. 5D View FIG ).
Palatine
The palatine is completely fused with the maxilla on IRSNB 3845-M.536; a slight bulging of the surface, going 165 mm anterior to the antorbital notch, probably corresponds to an area of muscle insertion. On the skull IRSNB 3847-M.537, the maxillopalatine suture is partly distinct, indicating a palatine reaching laterally close to the antorbital notch, and extending forward 135 mm anterior to the notch ( Fig. 5C View FIG ). Posteriorly, the lateral part of the bone goes beyond the anterior wall of the choana (incompletely preserved). The pterygoid is lost but a large portion of the palatine is excavated, either by the lost pterygoid, or by the pterygoid sinus fossa, only retaining a narrow unexcavated strip around the fossa. Grooves and ridges on the anterior margin of the fossa indicate the suture with the pterygoid. The remaining surface of the fossa is very smooth, the pterygoid being probably very thin or even absent there. The dorsolateral margin of the fossa is a low longitudinal ridge.
Lacrimal-jugal-optic groove
On the orbit roof, the lacrimal and the jugal are completely fused. The lacrimal-jugal complex is fused to the frontal and the anterior curved part of the maxilla. The optic groove is wide, with the anterior wall more elevated, separating it from the large infraorbital foramen. One or two small frontal foramina pierce the posterior wall of the groove. A foramen starting from the large infraorbital foramen in a posteromedial direction, presumably the sphenopalatine foramen, emerges on the lateral wall of the choana.
INTRASPECIFIC VARIATION
In Ziphius cavirostris View in CoL , which is the only recent ziphiid possessing a prenarial basin, this structure is considered as extremely sexually dimorphic, only well developed in adult males ( Heyning 1989a). While the prenarial basin roughly occupies the same position in Ziphirostrum marginatum , its morphology differs from Ziphius cavirostris View in CoL : in the former, the basin is not laterally margined by the elevated lateral portion of the premaxilla, but by the thickened maxilla. Furthermore, the basin is observed in all the preserved specimens of Ziphirostrum marginatum , large and smaller ones, with a nearly total resorption of the premaxillae anterior to the premaxillary sac fossae ( Fig. 6 View FIG ). The absence of resorption would give stronger skulls, more likely to be preserved; because this condition is not recorded in the collection of the IRSNB, the species Z. marginatum is probably characterized by the presence of a prenarial basin on specimens of both sexes and different ages. Width and length of the basin vary among the specimens – for instance, the distance between the posterior apex of the thickened premaxilla on the rostrum and the antorbital notch is greater on IRSNB 3845-M.536 relative to IRSNB 3847-M.537 (respectively 95 and 79 mm). However, it seems difficult to correlate this variation to ontogeny, contrary to Ziphius cavirostris View in CoL : the larger skull IRSNB 3846-M.1875 has a minimum width between the thickened lateral strips of maxillae that is smaller than on IRSNB 3 8 4 5-M.5 3 6 (respectively 2 7 and 34 mm). The premaxillary sac fossae of the skull IRSNB 3843-M.1876 are anteriorly longer in the basin than on IRSNB 3845-M.536, for a specimen similar in size. In Z. cavirostris View in CoL , the variably developed resorption of the premaxillae is related to the more or less complete filling of the mesorostral gutter ( Heyning 1989a), mainly by the vomer, while in Ziphirostrum marginatum the mesorostral gutter is not filled by the vomer, but is dorsally closed by the joined premaxillae on every specimen observed. The pattern of bones is therefore very different. Nevertheless, the filling of the prenarial basin with the enlarged right nasal plug, observed in the adult males of Ziphius cavirostris ( Heyning 1989a) View in CoL , is suggested in Ziphirostrum marginatum , but without distinction of sex and age.
The morphology of the premaxillae at the anteri- or limit of the prenarial basin is variable in Z. marginatum . On IRSNB 3845-M.536, the premaxillae are separated for a length of 30 mm. They are more closely appressed on the skull IRSNB 3847-M.537, but they present a narrow posterior projection towards the prenarial basin.
The skulls IRSNB 3842, IRSNB 3843-M.1876, IRSNB 3844-M.1874, and IRSNB 3846- M.1875 show more posteriorly diverging lateral margins of the prenarial basin, with more elevat- ed posterolateral crests compared to IRSNB 3845-M.536.
Height and width of the premaxillae anterior to the prenarial basin, are also variable (see Fig. 1A View FIG , Table 1, measurement 5 for the width). For instance, the premaxillae of IRSNB 3847-M.537 are wider and more elevated than in IRSNB 3845-M.536 and the large skull IRSNB 3846-M.1875. On IRSNB 3847-M.537, the lateral margins of the enlarged premaxillary dome nearly reach the lateral edges of the rostrum, and, at the level of the maximum elevation, the rostrum is higher than wide, contrary to IRSNB 3845-M.536 and IRSNB 3846-M.1875.
The narrowing of the premaxillae just before the transverse premaxillary crests is more pronounced on IRSNB 3847-M.537 than on IRSNB 3845- M.536, with minimum widths of the left premaxilla respectively of 30 and 38 mm.
Taking into account this intraspecific variability, several species of the genus Ziphirostrum described by du Bus (1868) should be referred to Z. marginatum . Each of these species was based on fragmentary rostral material, lacking the vertex. Hereafter, several characters of these nominal species are briefly discussed to examine the variability and the anatomical data they add to Z. marginatum .
ZIPHIROSTRUM LAEVIGATUM SENSU DU BUS, 1868 The species Ziphirostrum laevigatum sensu du Bus, 1868 was erected for the specimen IRSNB 3830-M.542, a partial skull figured by Van Beneden & Gervais (1880: pl. 27bis, fig. 5). The definition of the species was based on different proportions of the rostrum in general, on the thickness and the porosity of the maxillae, the size of the prenarial fossa, and the proportions and porosity of the premaxillae ( du Bus 1868). Some of these characters were later used to suggest affinities with Z. laevigatum for the specimen IRSNB M.1877, a more complete skull collected in non-reworked deposits ( Bosselaers et al. 2004). For these two skulls, the general morphology of the rostrum matches that described above, and the few differences observed – the anterior part of the rostrum wider than high, due to the poorly elevat- ed premaxillae, the lateral part of the premaxillae forming with the lateral maxillae a concave dorsolateral surface, and a longer separation between the premaxillae before the prenarial basin – are considered intraspecific variation. The density of the rostrum bones, used by du Bus (1868) to distinguish Z. laevigatum , will be discussed in another work. The degree of rostral mineralization is far from consistent in Z. marginatum , and should not be used as a diagnostic character.
A strange characteristic of the skull IRSNB 3830- M.542 is the morphology of the left maxilla posteriorly to the antorbital notch ( Fig. 7 View FIG ). Even if the notch is only partially preserved, the lateral margin of the maxilla, some centimetres posterior to the notch, extends no further laterally than just before the notch: the preorbital process forms a reduced lateral projection anterior to a strong narrowing of the maxilla with a deep inner curvature. The frontal being poorly preserved under the maxilla, this strange feature, not observed on other specimens of Z. marginatum , seems difficult to explain – very narrow supraorbital process or only resorption of the maxilla. The surface of the maxilla at that level is relatively smooth, and does not provide indications for a pathology derived from a trauma.
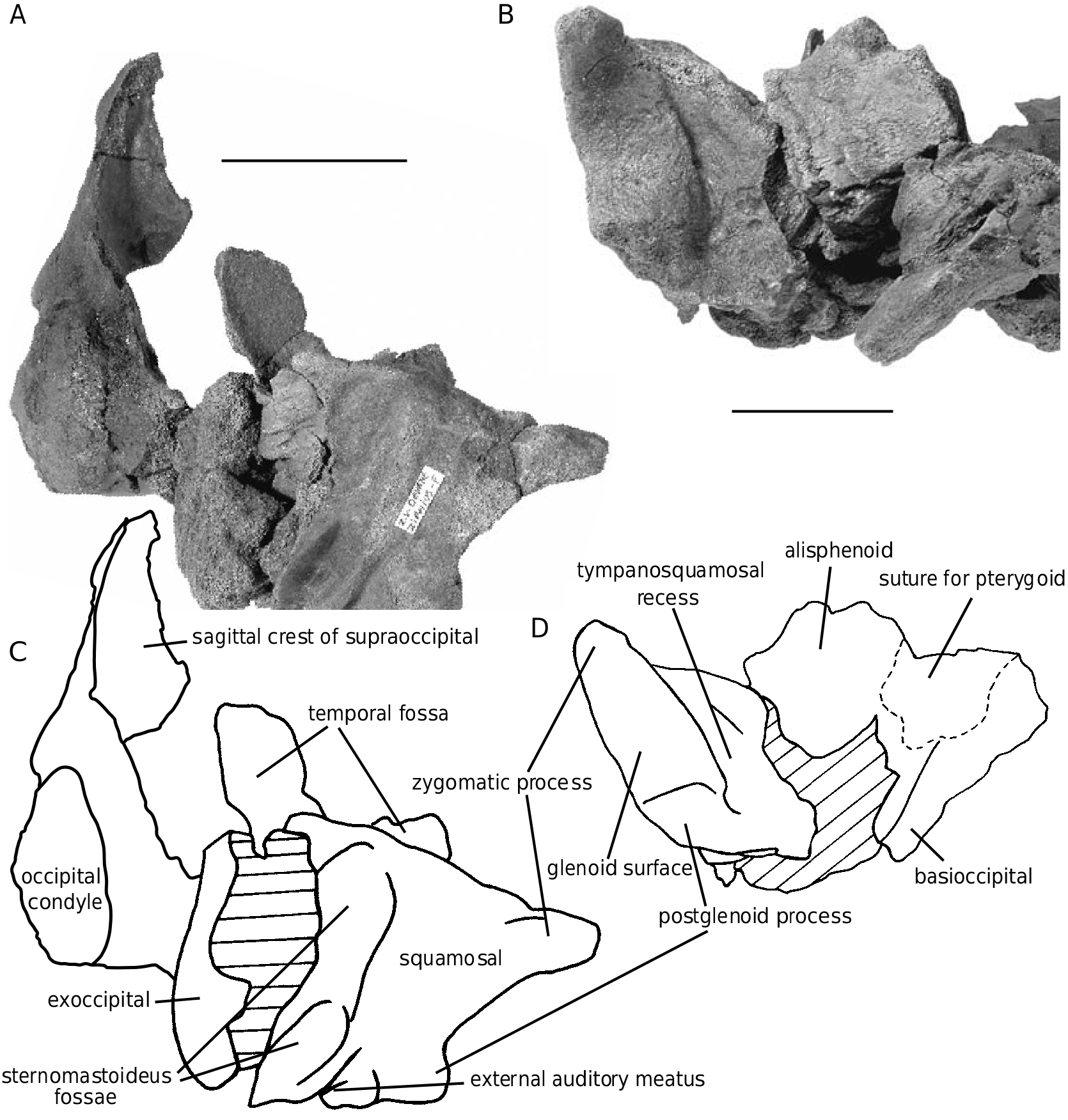
The specimen IRSNB M.1877, with more porous bone, has the left squamosal, fragments of the left exoccipital, the left occipital condyle, fragments of the basioccipital and supraoccipital, and four teeth preserved, parts never described for the species. In lateral view, the squamosal ( Fig. 8 View FIG ) shows a long and anterolaterally pointed zygomatic process, perpendicular to the wide and short postglenoid process. This articular part of the bone is separated from the post-tympanic process by a weakly marked oblique ridge, forming the anterior wall of the two sternomastoideus fossae; the lower fossa is deep and narrow and the upper one is wider. The glenoid surface occupies a large part of the ventral surface of the squamosal, laterally to a poorly excavated and narrow tympanosquamosal recess (compared to Ziphius for instance). The floor of the temporal fossa is a wide basin, more distinct than in Ziphius .
wear facet from opposite tooth
The four teeth ( Fig. 9 View FIG ) were found detached near the skull IRSNB M.1877 on the field; whether upper or lower is uncertain. Their length ranges from 18 to 22 mm. All of them have a variously curved smoothly enamelled crown, slightly anteroposteriorly flattened, constituting less than half the total length of the tooth. The root is thick, wide (maximum diameter between 8 and 9 mm), nodulous, and rectilinear. The size of the root is too large for the shallow maxillary alveoli described on Ziphirostrum marginatum IRSNB 3847-M.537; this might suggest a dentary origin for those teeth. The apex of the root is hollowed by a circular pulp cavity variously filled (one of the teeth has a deep and narrow empty cavity, characteristic of younger individuals). Three of the teeth show an irregular apical wear of the crown. Additionally, one of them bears a smooth wear facet at the base of the crown, perfectly matching the shape of a crown. This feature probably indicates the presence of opposite functional teeth (lower and upper jaw teeth), a primitive character shared with the extant Tasmacetus , the Pliocene Ninoziphius platyrostris and the Miocene Messapicetus longirostris . On a specimen of Tasmacetus shepherdi, Oliver (1937) also noticed signs of wear on the crowns of the teeth, many truncated tips, and grooves where opposing teeth have worn against them; Oliver concluded that those teeth were functional. There is a great variation of size and shape for the teeth of extant ziphiids, partially due to ontogeny and sexual dimorphism (e.g., Mead 1989). The swelling of the root and the blunt proximal extremity in the f o s s i l t e e t h a r e r e m i n i s c e n t o f t h e t e e t h o f Hyperoodon ampullatus figured by Moore (1968: figs 4, 5) and Tasmacetus shepherdi figured by Oliver (1937: pl. 4, fig. 10). Similarly, Ninoziphius platyrostris has a wide tooth root, but the crown is too worn to be compared (see Muizon 1984). Furthermore, the teeth of N. platyrostris are larger, with a width of the roots reaching 12 mm. On the holotype of that species, the long roots are only partially inserted in the alveolar groove, a feature that could be partially due to a post-mortem shifting; it is, however, likely that a portion of the root was included in the gum.
ZIPHIROSTRUM MARGINATUM SENSU DU BUS, 1868 The holotype, IRSNB 3783-M.1878 ( Fig. 10 View FIG ), was originally the only specimen referred to Ziphirostrum marginatum by du Bus (1868). That species was defined by the relative width and shape of the premaxillae and maxillae in dorsal view of the rostrum, and the size of the prenarial fossa; the premaxillae are a little bit narrower on the rostrum and the maxillae flatter and wider with a more acute lateral margin than in most of the other specimens of Z. marginatum . From the premaxillary foramen, an anteromedial sulcus excavates the thickened premaxilla on a short distance; anteriorly, the sulcus divides in a series of narrow and shallow sulci spreading on the surface of the premaxilla. A deep (more than 8 mm) and wide posterior sulcus connects the premaxillary foramen to the anterior of the premaxillary sac fossa, where it divides in short posteromedial and posterolateral sulci, vanishing before the level of the antorbital notch. On the other specimens of Ziphirostrum marginatum , this fragile structure is either broken, or resorbed by deepening of the prenarial basin; the morphology described here indicates how the prenarial basin develops in Z. marginatum , by deepening of a sulcus consisting in the joined anterior portions of the posteromedial and posterolateral sulci.
The holotype shows interesting similarities with Messapicetus longirostris , from the late Miocene of Italy ( Bianucci et al. 1992), including the narrow and moderately high premaxillae anteriorly to the prenarial basin, median to wide and flat maxillae, and the poorly excavated prenarial basin with a pair of wide longitudinal sulci bordered medially by a prominent ridge of the premaxilla and laterally by the thick maxilla. However, Messapicetus has more distinct and ventrally oriented maxillary alveoli, and the rostrum is longer. Messapicetus longirostris might nevertheless be closer to Ziphirostrum marginatum than previously thought (see Bianucci et al. 1992, 1994, and phylogenetic discussion below).
ZIPHIROSTRUM GRACILE SENSU DU BUS, 1868
The fragmentary rostrum IRSNB 3827-M.1879 is the only specimen of Ziphirostrum gracile sensu du Bus, 1868 found in the collection. The two others cited by Abel (1905), including the one on which du Bus (1868) noticed several deeper alveoli, are lost. This specimen is composed of fused rostral portions of the premaxillae with applied fragments of maxillae ( Fig. 11 View FIG ). The dorsolateral surfaces of the rostrum are concave, as in IRSNB 3830-M.542; the premaxillae are posteriorly separated on more than 90 mm, a character also present on the specimens IRSNB 3820 and IRSNB 3828, and considered as an individual variation in the species Ziphirostrum marginatum . “More porous maxillae” is the only character used by du Bus (1868) to define Z. gracile , a character unreliable in Z. marginatum . The maximum width of the premaxillae on the rostrum (50 mm) matches the variability observed in Z. marginatum .
A
MANDIBLES PREVIOUSLY REFERRED TO MIOZIPHIUS BELGICUS
In 1905, Abel placed in his carryall species Mioziphius belgicus several isolated symphyses and partial rami of mandibles. The best-preserved specimen, IRSNB 3854-M.538 (named Synostodon sp. by du Bus [in Vanden Broeck 1874], and identified by Abel [1905: fig. 19] as a cotype of Mioziphius belgicus ), is mainly composed of a fused symphysis (177 of the 215 mm of the fragment), hollowed by two pairs of prominent alveoli for enlarged apical teeth ( Fig. 12 View FIG ). The first anterior pair is the largest, constituting the anterior rounded margin of the mandible. Those alveoli are slightly longitudinally elongat- ed, with a maximum length of 28 mm. The alveoli of the second pair, separated from the first by 15-20 mm, are distinctly narrower and have a length of 25 mm. A pointed anterodorsally directed peg is present at the centrum of the first and near the anterior wall of the second of these cylindrical depressions. The alveoli are inserted in a wide groove (8-9 mm wide between the first and second alveoli). 6-7 mm posteriorly to the second alveolus, the groove is filled by a continuous series of small rounded bony pegs. The external aspect of those pegs is reminiscent of the alveolar groove of the mandible of a fetus of Physeter macrocephalus (ZMA 12.810, total length of the body = 2540 mm), with the prominent but unerupted teeth still embedded in the bone of the alveolar groove. A lateral longitudinal groove is present on the posterior few centimetres. Ventrally, the joined acute medioventral edges of the rami progressively diverge, opening a long triangular depression which opens dorsally only after 60 mm. Five pairs of small foramina pierce the mandible on both sides of the ventral symphysis, on the apical 25 mm. Width and height of the mandible at the level of the posteri- or wall of the first alveolus are respectively of 44 and 37 mm, and at the posterior end of the symphysis of 53 and 36 mm.
Following the interpretation of Abel (1905), the pointed pegs on the enlarged alveoli are probably a center of fixation for the root of poorly-inserted large apical teeth. A reduction of the number of mandibular teeth and the development of a few pairs of apical enlarged teeth is one of the main characteristics of the ziphiids. Among extant ziphiids, the genera Hyperoodon , Ziphius and Mesoplodon only keep one pair of apical (or subapical) mandibular teeth, two pairs are present in Berardius , with some variation, and about 48 pairs occur in Tasmacetus (with one apical pair of slightly greater size) ( Moore 1968). The teeth of Indopacetus are not known. The shallow protruding alveoli of this specimen, partially filled with porous bone, can be compared to the platforms of dendritic or spongiform bone in Tasmacetus , or to the spongiform pads in Berardius ( Moore 1968) . They also fit the description made by True (1910) in Ziphius : “[…] in adult males the teeth are almost entirely protruded from the alveoli, which are filled with a coarse bony network”. In Ziphius , the filling of the alveoli forces the teeth up and causes them to erupt in adult males ( Heyning 1989b). Moore (1968) suggested that the same process happens in males of Hyperoodon but not in Mesoplodon . The mandible of Ninoziphius (early Pliocene, Peru) bears three apical and subapical pairs of “exostoses”, also suggested as the support of enlarged teeth ( Muizon 1984). The similarities of the smaller posterior pegs in the alveolar groove with the situation in fetal Physeter might provide an idea of the way the reduction of the number of teeth occurs in the ziphiids, by retention of a juvenile condition for a large part of the teeth in adults, and an “extra” growth of only a small number of them.
The more eroded symphyses IRSNB 3848 and IRSNB 3855, even if smaller, do not show strong differences with IRSNB 3854-M.538; they might belong to the same taxon. Nevertheless, all these specimens are isolated, and their size and characters might correspond equally to Ziphirostrum sp. , Choneziphius sp. or Aporotus sp. (see below). Accordingly, these symphyses are considered as Ziphiidae incertae sedis.
For two isolated fragments of left ramus IRSNB 3851 and IRSNB 3852 referred to Mioziphius belgicus by Abel (1905), few characters are comparable to the above symphyses; on both specimens, the preserved symphyseal portion does not seem to be tightly fused, and the dorsal groove is hollowed by shallow and poorly distinct alveoli. Because no clear ziphiid characters are apparent, these undiagnostic fragments are placed in Odontoceti incertae sedis.







| IRSNB |
Institut Royal des Sciences Naturelles de Belgique |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ziphirostrum marginatum du Bus, 1868
| Lambert O. 2005 |
Mioziphius belgicus
| ABEL O. 1905: 99 |
Ziphirostrum marginatum du Bus, 1868: 624
| DU BUS B. A. L. 1868: 624 |
Ziphirostrum laevigatum du Bus, 1868: 624
| DU BUS B. A. L. 1868: 624 |
Ziphirostrum gracile du Bus, 1868: 625
| DU BUS B. A. L. 1868: 625 |