
Mylossoma duriventre ( Cuvier, 1818 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4387.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:366DF9BF-EA85-4DB4-9857-8E6D7B3B3E31 |
|
DOI |
https://doi.org/10.5281/zenodo.5965013 |
|
persistent identifier |
https://treatment.plazi.org/id/014587BD-9254-FFF5-FF2C-F980B42FF95C |
|
treatment provided by |
Plazi |
|
scientific name |
Mylossoma duriventre ( Cuvier, 1818 ) |
| status |
|
Mylossoma duriventre ( Cuvier, 1818) View in CoL
( Figs. 16 – 19 View FIGURE 16 View FIGURE 17 View FIGURE18 View FIGURE 19 , Table 4)
Myletes duriventris Cuvier, 1818: 451 View in CoL [original description; type-locality: " Brésil "].— Bertoni, 1939: 56 [ Paraguai]— Bertin, 1948: 28 [type-material listed].
Myletes orbignyanus Valenciennes, in Cuvier & Valenciennes, 1850: 220 View in CoL [original description; type-locality: "au-dessus de Corrientes, dans le Parana"].— Bertoni, 1939: 56 [Paraguai].— Bertin, 1948: 28 [catalog of type-material].
Mylossoma albiscopus View in CoL (non Cope)— Eigenmann & Kennedy, 1903: 530 [rio Paraguay and Laguna at Asuncion].
Mylosoma [ sic] albiscopus .— Eigenmann, 1915: 266 [ partim; redescription].
Mylosoma [ sic] ocellatum Eigenmann, 1915: 265 View in CoL [original description; type-locality: "Villa Hays, Paraguay"].— Henn, 1928: 69 [type-material listed].
Mylossoma duriventris View in CoL .— Norman, 1929: 813 [ partim; brief description].— López et al., 2003: 34 [occurrence in Argentina].— Menni, 2004: 79 [occurrence in Argentina].
Mylossoma paraguayensis Norman, 1929: 814 View in CoL [original description; type-locality: "Rio Paraguay "].— MacDonagh & Fuster, 1944: 355 [redescription].— Géry, 1976: 50 [listed].— Oldani, 1983: 89 [larvae morphology].— Miquelarena, 1986: 37 [description of dentition].— Sverlij et al., 1998: 25 [río Uruguay].— López et al., 2003: 34 [ Argentina].— Menni, 2004: 79 [ Argentina].— Britski et al., 2007: 81 [Pantanal].
Mylosoma [ sic] aureum (not Spix & Agassiz)— Bertoni, 1939: 56 [Paraguay].
Mylossoma duriventre View in CoL .— Géry, 1976: 50 [listed].— Gómez & Chebez, 1996: 52 [Misiones Province, Argentina].— Jégu, 2003: 187 [listed].— Menni, 2004: 79 [Argentina].— Graça & Pavanelli, 2007: 82 [upper rio Paraná].— Langeani et al., 2007: 185 [upper rio Paraná, listed].— Litz & Koerber, 2014: 9 [Uruguay].— Mirande & Koerber, 2015: 15 [Argentina].— Koerber et al., 2017: 22 [Paraguay].
Mylossoma duriventris orbignyanum View in CoL .— Géry et al., 1987: 443 [lectotype designation; Paraguay, taxonomic comments].— Gómez & Chebez, 1996: 52 [Misiones Province, Argentina].
Mylossoma ocellatum View in CoL .— Ibarra & Stewart, 1987: 58 [type-material listed].
Mylossoma orbignyanum View in CoL . — Britski et al., 2007: 81 [Pantanal]
Mylossoma orbignyanus View in CoL .— López et al., 2003: 34 [ Argentina].— Menni, 2004: 79 [ Argentina].
Diagnosis: Mylossoma duriventre differs from M. aureum by presenting the last abdominal spine reaching the anal-fin origin or almost so ( vs. last abdominal spine clearly separated from the anal-fin origin), 35–36 vertebrae ( vs. 38–39) and a conspicuous black blotch on opercle ( vs. black blotch absent or inconspicuous). It differs from M. albiscopum by presenting 26 to 32 branched rays on the anal fin ( vs. 31 to 38). Finally, it differs from M. unimaculatum by presenting 30 to 34 circumpeduncular scales ( vs. 34 to 40), 74 to 98 perforated scales on lateral line ( vs. 95 to 110) and 35–36 vertebrae ( vs. 37).
Description: Morphometric data presented in Table 4. Body deep, compressed laterally. Dorsal profile slightly concave at posterior region of head and convex between head and dorsal fin. Ventral profile convex; slightly concave at isthmus and strongly convex from this point to end of anal fin. Highest body depth on vertical line passing through dorsal- and pelvic-fins origins. Caudal peduncle as deep as long.
Eyes lateral at middle of head; upper margin of eyes below longitudinal axis of lateral-line origin. Frontal and parietal fontanel broadly expanded laterally. Snout short and rounded on lateral view. Nostrils dorsolateraly positioned, above longitudinal axis through upper margin of eye.
Mouth terminal to slightly upturned, at same level of orbits. Premaxilla projected forward, with inner premaxillary row over or surpassing dentary teeth. Premaxillary teeth molariform with robust base; outer portion of anteriormost three teeth of outer row and posterior portion of remaining teeth pointed. Inner premaxillary row with two teeth, separated from outer row. Outer premaxillary row with five teeth. Dentary with four or five tricuspid, robust, molariform teeth; symphyseal dentary teeth present, with robust base and large conical cusp, behind main row ( Fig. 2 View FIGURE 2 ). Maxilla edentulous.
First gill arch with elongated and conical gill rakers; epibranchial gill-rakers 10(2), 12(20), 13(2), 14(23) or 15(2); one (50) gill raker at cartilage between epi- and ceratobranchial; ceratobranchial gill rakers 11(2), 13(4), 14(1), 15(22), 16(5), 17(9) or 18(6).
Body completely covered by small cycloid scales. Accessory scales covering intersections of major scales, mainly at anterior portion of body. Lateral line complete, with 74(1), 76(1), 80(2), 81(1), 82(2), 83(1), 84(1), 85(5), 86(7), 87(3), 88(1), 89(8), 90(9), 91(3), 92(5), 93(3), 94(2), 95(2), 96(2), 97(2) or 98(1) perforated scales, extending into caudal fin. Scales above lateral line 51(1), 54(3), 56(3), 57(5), 58(1), 59(1), 60(5), 61(4), 62(1), 63(4), 64(5), 65(3), 66(4), 67(3), 68(5), 69(5), 70(2) or 73(2). Scales below lateral line 41(1), 43(1), 44(1), 45(2), 46(1), 47(1), 48(1), 49(1), 50(2), 51(2), 52(6), 53(4), 54(7), 55(4), 56(6), 57(5), 58(2), 59(2), 60(1), 61(2) or 65(1). Circumpeduncular scales 30(9), 32(32), 33(4) or 34(8). Ventral keel with well-developed spines forming high serra. Prepelvic spines 26(2), 27(2), 28(6), 29(13), 30(10), 31(11), 32(7), 33(7), 34(3) or 36(2). Postpelvic spines 11(1), 12(7), 13(22), 14(19), 15(9), 16(3) or 17(3). Anal spines 5(10), 6(32), 7(16) or 8(4).
Dorsal fin not preceded by spine, its origin equidistant from tip of snout and end of hypural plate. First dorsal-fin ray much shorter than second ray; in some specimens covered by skin, in such cases discernible only if dissected or x-rayed. Branched dorsal-fin rays gradually decreasing in size; dorsal-fin rays 12(19), 13(37), 14(4) or 15(3). Adipose fin short, entirely covered by small scales. Pectoral fin falcate, with i(63) + 11(1), 12(1), 14(4), 15(19), 16(24), 17(11) or 18(2) rays. Pelvic fin small, with i(61) + 5(6) or 6(55) branched rays. Anal fin long, with convex edge, median and posterior rays longer than anterior ones, not lobed; sheath of scales covering at least two-thirds of anal-fin length; branched anal-fin rays 26(2), 27(4), 28(7), 29(10), 30(24), 31(16) or 32(2). Caudal fin bifurcated, lobes of similar size, with 16(3), 17(49), 18(5) or 19(1) rays. Vertebrae 35(2) or 36(11). Supraneurals 4(9).
Color in alcohol. General body color light brown darker dorsally ( Fig. 17 View FIGURE 17 ). Fins pale yellow; often with bases of caudal and anal fins and distal margin of anal fin dark. Conspicuous black blotch on opercle. Juvenile specimens with several vertical pale brown bands on flanks and a round brown ocellar blotch below dorsal fin ( Fig. 18 View FIGURE18 ); the bands may persist, although lighter, in well-preserved adult specimens.
Color in life. General body color silvery, darker dorsally; yellowish pigmentation on ventral portion of chest and head; region around eyes orange. Presence of a black blotch on opercle. Anal fin yellow or orange, with a black band on distal margin; caudal fin with an orange band on distal margin; remaining fins hyaline; several vertical grey bands on flanks ( Fig. 19 View FIGURE 19 ).
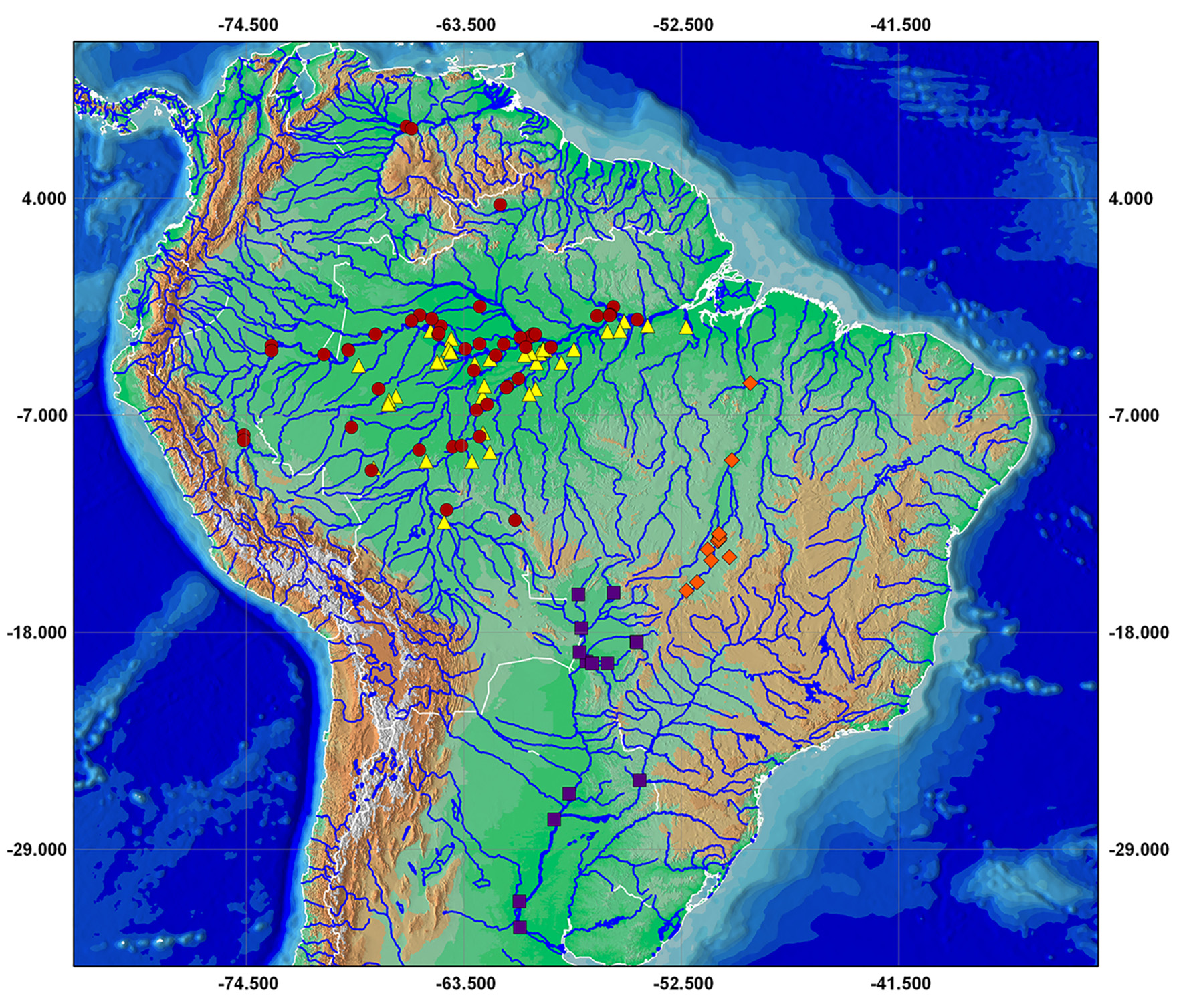
Geographic distribution. Mylossoma duriventre is known from the rio Paraguai, lower Paraná and lower Uruguai rivers basins ( Fig. 3 View FIGURE3 ).
Ecological notes. Mylossoma duriventre is an herbivore fish, highly dependent from the floodplains and presenting a great economic importance for both commercial and sport fisheries ( Resende et al. 1998; Sverlij et al. 1998; Chernoff et al. 2001).
Molecular data. DNA barcoding revealed that the genetic distance between M. duriventre ( M. duriventre "group 3" in Mateussi et al. 2016) and the remaining species analyzed ranged from 0.014 to 0.090 and that this species is genetically most similar to M. unimaculatum ( M. duriventre "group 4" in Mateussi et al. 2016).
Remarks. Cuvier (1818) in the original description cited the type-locality of Myletes duriventris as “ Brésil ” at the end of species description: " Mais ce même Brésil nous a fourni deux autres espéces de ce genre ". The label of the holotype indicate that it belonged to the so-called “Cabinet d’Ajuda”, a collection made by Alexandre Rodrigues Ferreira housed at Museu da Ajuda in Lisbon, Portugal, which was ransacked by Étienne Geoffroy Saint-Hillaire during the military expedition of the Napoleonic armies to Portugal, headed by the General Junot ( Myers 1950; Vanzolini 2004). Alexandre Ferreira made several journeys across the Brazilian Amazon and traveled for almost three years throughout the Paraguai basin. Vanzolini (2004) emphasized that there are doubts about the exact provenance for specimens collected by Ferreira, since the specimens’ origins were not indicated on his drawings nor recorded at the Museu d’Ajuda. Since the holotype of M. duriventris presents iii+29 anal-fin rays, which is within the range from specimens from the rio Paraguai basin, our results suggest that the name Mylossoma duriventre should be used for the only species of the genus occurring in this river basin. Thus, records of M. duriventre from the Amazonas, Orinoco and Tocantins-Araguaia basins are herein considered as incorrect and referable to M. albiscopum ( Amazonas and Orinoco) or M. unimaculatum (Tocantins-Araguaia) . The inability of previous authors in recognizing the distinction of these three species resulted in accounts mixing specimens of more than one species, as in Eigenmann (1915) and Norman (1929).
Géry et al. (1987) discussed the validity of the species of Mylossoma from the La Plata basin (cited on their paper as M. orbignyanum and M. paraguayensis ). These authors argue the diagnose proposed by Norman (1929) (i.e. number of dorsal and anal fins branched rays and the proportion between adipose and anal-fin length), considering that only dorsal-fin branched rays could be used, "at least provisionally", to differentiate M. orbignyanum at the subespecific level. However, during the present study, we concluded that this character is not in fact diagnostic. These authors discarded the possibility of anal-fin rays as a diagnostic character since the type of M. duriventre presents 29 branched rays and according to Norman (1929), M. duriventre should present 28 to 34 and M. paraguayensis 26 to 31 anal-fin branched rays. Also, Géry et al. (1987) countings on specimens from Paraguay basin reaches 33 branched rays at anal-fin. However, after examining about 80 Mylossoma specimens from the Paraná-Paraguai basin, we were unable to find specimens presenting more than 32 branched anal-fin rays. Machado- Allison & Castillo (1992) found the same overlap proposed as diagnosis by Géry et al. (1987) and, instead, proposed that the differences on color pattern between specimens from the La Plata basin (presence of vertical dark bands in adults) and Amazon basin (absence of such bands) as diagnostic between these populations. Both Géry et al. (1987) and Machado-Allison & Castillo (1992) considered M. paraguayensis as a synonym of M. orbignyanum , and the latter as a subspecies of M. duriventre . However, those authors considered that the type-locality of Mylossoma duriventre lies at the Amazon basin rather than the rio Paraguai basin. We herein confirmed that anal-fin rays counts and the color pattern are the best features to diagnose both species. Additionally, and as detailed below, Mateussi et al. (2016) provided DNA barcode evidence supporting the distinctness of both species. Thus, we herein consider M. orbignyanum and M. paraguayensis as both synonymies of M. duriventre and restrict the distribution of the latter species to the rio Paraguai basin, and lower portions of the rio Paraná and rio Uruguai basins.
Langeani et al. (2007) and Graça & Pavanelli (2007) listed Mylossoma duriventre as occurring at the upper rio Paraná basin, Brazil, based on the lot NUP 2158. However, this specimen was actually collected at the rio Cuiabá, upper rio Paraguai basin, Brazil. No records of M. duriventre were found upstream from the Sete Quedas Falls that, until 1982, consisted in a natural, ecological barrier splitting populations from the lower and upper rio Paraná ( Vitule et al. 2012). Since then, the faunistical barrier moved 200 km downstream at the Itaipu hydroelectric dam, which submerged the Sete Quedas Falls. We thus consider the purported occurrence of M. duriventre at the upper rio Paraná basin as incorrect.
Britski et al. (2007) listed both Mylossoma paraguayensis and M. orbignyanum as valid species for the Pantanal, rio Paraguai basin, Brazil, and remarked that those species could be diagnose by presenting distinct color patterns, M. paraguayensis presenting sharper vertical bands on flanks than M. orbygnyanum . However, our present analysis did not confirm this purported diagnosis between those two nominal species, and consequently we consider that a single Mylossoma species occurs at the La Plata basin.
Unfortunately, no specimens from the rio Uruguai basin were analyzed in the present study. Sverlij et al. (1998) presented a short description and a picture of Mylossoma duriventre from the rio Uruguai basin and it matches the diagnosis presented herein based on the number of anal-fin rays and the color pattern.
We were not able to examine the original description of Salmo trigintaradiatus by Larrañaga (1923). Our comments about this nominal taxon are based on Devincenzi (1925), who transcribed and commented the original descriptions by Larrañaga (1923). Salmo trigintaradiatus was poorly described, no drawings were presented or type specimens designated, which hinders any attempt at the definition of this nominal species. Devincenzi (1925) hypothesized that this species represented an unknown species or as a probable synonym of Myletes orbignyanus , although mentioning some disagreement between the description by Larrañaga (1923) and the latter species. Jégu (2003) followed Devincenzi (1925) in considering S. trigintaradiatus as a synonym of Mylossoma duriventre . In contrast, Lima et al. (2003) considered S. trigintaradiatus as species inquirendae in Characidae . Due to the lack of types, drawings and an adequate description that could help us to define this nominal species, and considering that the character " mandibula inferior longior " in the description is not present in any species of Mylossoma , we agree with Lima et al. (2003) and prefer to consider Salmo trigintaradiatus as a species inquirendae in Serrasalmidae .
Material examined. Type material. MNHN A.9891, 1 (rd), 181 mm SL, Brasil. Holotype of Myletes duriventris . MNHN A.9863, 1 (rd), 230 mm SL, above Corrientes, río Paraná. Lectotype of Myletes orbignyanus (designated by Géry et al., 1987). FMNH 56816, 1 (rd), 44 mm SL, Villa Hays, Paraguay. Holotype of Mylosoma ocellatum . BMNH 1895.5.17.254–255, 2 (rd), rio Paraguai; BMNH 1910.5.26.47–48, 2 (rd), Puerto Asir, Paraguay; BMNH 1910.5.26.46, 1 (rd), San Salvador, Paraguay. Syntypes of Mylossoma paraguayensis . Non-type material. Argentina. Río Paraguay basin. Formosa: MLP 6466, 2, 18.7–23.2 mm SL, Formosa, lagoon adjacent to río Paraguay, 26°11'05"S 58°10'30"W (city coordinates), Dec 1954. MLP 6472, 1, 15.5 mm SL, Formosa, río Paraguay, 26°11'05"S 58°10'30"W (city coordinates), Nov 1957. Lower río Paraná basin. Chaco: MLP 7870, 7, 21.5–30.1 mm SL, Barranqueras, confluence río Negro y riachos, 27°28'56"S 58°56'00"W (city coordinates). Santa Fe: MLP 6803, 1, 77.6 mm SL, Rosario, río Paraná, 32°57'02"S 60°39'59"W (city coordinates), 12 Dec 1960. MLP 7646, 4, 75.1–82.9 mm SL, Santa Fe, Madrejón Don Felipe, 31°38'12"S 60°41'58"W (city coordinates), 31 May 1963. Brazil. Rio Paraguai basin. Mato Grosso: LBP 4256, 1, 174.7 mm SL, Cáceres, rio Cabaçal (trib. rio Paraguai), c. 15°40'S, 57°48'W, 0 1 Jun 2005, W. Troy. LBP 4257, 2 (rd), 172.1–185.2 mm SL, Cáceres, rio Paraguai, 16°04'36"S 57°40'56"W (city coordinates), 0 1 May 2006, W. Troy. LBP 4674, 1 (rd), 163.1 mm SL, Santo Antônio de Leverger, rio Aricá (trib. rio Paraguai), 15°59'15"S 55°55'42"W, 15 Jun 2007, W. Troy. LIRP 9628, 2, 121.8 mm SL, Poconé, rio Bigueirinho, 17°47'34"S 57°33'27"W, 30 Oct 2011, CEPTA team. NUP 1004, 4, 31.5–72.6 mm SL, Chapada dos Guimarães, Reservatório da Usina Hidrelétrica de Manso, 14°41'46"S 56°15'13"W, 14 Nov 2003, Nupélia team. NUP 1017, 5, 153.4– 166.3 mm SL, Chapada dos Guimarães, Reservatório da Usina Hidrelétrica de Manso, 14°41'46"S 56°15'13"W, 14 Nov 2003, Nupélia team. NUP 2054, 1, 170,4 mm SL, Barão de Melgaço, rio Cuiabá, 16°20'02"S 55°57'10"W, 24 May 2003, Nupélia team. NUP 2158, 3, 70.8–76.8 mm SL, Santo Antônio do Leverger, rio Cuiabá, 15°58'26"S 55°56'26"W, Mar 2000, Nupélia team. NUP 15258, 1, 79.8 mm SL, Barão de Melgaço, baía Sinhá Mariana, rio Cuiabá, 16°20'20"S 55°54'10"W, 25 Oct 2003, Nupélia team. NUP 15259, 1, 96.6 mm SL, Barão de Melgaço, baía Sinhá Mariana, rio Cuiabá, 16°20'20"S 55°54'10"W, 20 Sep 2003, Nupélia team. NUP 15275, 1, 170.4 mm SL, Barão de Melgaço, rio Cuiabá, 15°58'26"S 55°56'26"W, 24 Apr 2004, Nupélia team. Mato Grosso do Sul: DZSJRP 5479, 1, 204.7 mm SL, Morro do Azeite, rio Miranda, 19°29'02"S 57°17'49"W, V. Garutti. DZSJRP 7802, 1, 202.9 mm SL; DZSJRP 7803, 1, 193.0 mm SL, Miranda, Morro do Azeite, Fazenda Bodoquena, rio Miranda, 20°07'02"S 56°44'47"W, Nov 1989, V. Garutti. LBP 34, 1, 68.0 mm SL, Corumbá, rio Miranda, 19°34'S 57°01'W, 30 Jul 1996, C. Oliveira et al. LBP 1771, 3, 141.0– 183.1 mm SL, Coxim, rio Taquari, Dec 2002, D. C. Pompiani. LBP 3741, 6, 123.3– 146.6 mm SL, Aquidauana, rio Negro (trib. rio Paraguai), 19°34'33"S 56°14'49"W, 0 1 Aug 2006, C. Oliveira et al.. LBP 5148, 1, 157.9 mm SL, Coxim, rio Taquari, 18°28'33"S 54°46'38"W, 20 Aug 2007, W. Troy. LBP 9822, 15, 15.3–35.2 mm SL, Miranda, rio Miranda, 19°34'58"S 57°01'18"W, 22 Nov 2009, C. Oliveira et al.. LBP 12623, 4, 142.5– 180.5 mm SL, Corumbá, rio Cuiabá, 19°00'29"S 57°39'05"W (city coordinates), 24 Oct 2010, R. Britzke et al. MZUEL 0 7415, 14 (rd), 137.8–175.7 mm SL, Corumbá, Passo do Lontra, rio Miranda, 19°34'37"S 57°00'42"W, O. A. Shibatta et al.. MZUEL 0 7416, 1 (rd), 155.9 mm SL, Corumbá, Passo do Lontra, rio Miranda, 19°34'37"S 57°00'42"W, 15 Jul 2005, O. A. Shibatta et al.. MZUSP 83762, 1, 188.0 mm SL, Coxim, rio Taquari, 18°30'42"S 54°45'34"W (city coordinates), 14 Dec 1976, CEPIPAM. NUP 12551, 1, 85.8 mm SL, Porto Murtinho, rio Paraguai, 21°42'07"S 57°53'33"W, 25 May 2010, Y. Súarez. NUP 14219, 1, 105.8 mm SL, Corumbá, lagoa Doze, rio Paraguai, 19°24'21"S 57°18'11"W, 26 Mar 2012, Nupélia. NUP 15260, 1, 164.9 mm SL, Corumbá, lagoa Albuquerque, rio Paraguai, 19°26'S 57°22'W, 27 Mar 2012, Nupélia team. NUP 17182, 2, 72.5–92.5 mm SL, Porto Murtinho, rio Paraguai, 21°41'00''S 57°45'00''W, 23 Mar 2010, Y. Súarez. ZUFMS 0 119, 2 (rd), 102.2–120.4 mm SL, Corumbá, Passo do Lontra, rio Miranda, 19°34'37"S 57°00'42"W, 29 Dec 1990, G. Soares. ZUFMS 1679, 1 (rd), 107.7 mm SL, Corumbá, Passo do Lontra, rio Miranda, 19°34'37"S 57°00'42"W, 0 1 Apr 1992, J. C. Louzan & E. Pereira. Lower rio Paraná basin. Paraná: MZUSP 43995, 2 (rd), 106.7–220.1 mm SL, Foz do Iguaçu, lower rio Paraná, downstream Reservatório de Itaipu, 25°25'59"S 54°34'59"W, 15 Jan 1986, Nupélia team.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mylossoma duriventre ( Cuvier, 1818 )
| Mateussi, Nadayca T. B., Oliveira, Claudio & Pavanelli, Carla S. 2018 |
Myletes duriventris
| Cuvier, 1818 : 451 |
| Bertoni, 1939 : 56 |
| Bertin, 1948 : 28 |
Myletes orbignyanus Valenciennes, in Cuvier & Valenciennes, 1850 : 220
| Cuvier & Valenciennes, 1850 : 220 |
| Bertoni, 1939 : 56 |
| Bertin, 1948 : 28 |
Mylossoma albiscopus
| Eigenmann & Kennedy, 1903 : 530 |
Mylosoma [ sic ] albiscopus
| Eigenmann, 1915 : 266 |
Mylosoma [ sic ] ocellatum
| Eigenmann, 1915 : 265 |
| Henn, 1928 : 69 |
Mylossoma duriventris
| Norman, 1929 : 813 |
| López et al., 2003 : 34 |
| Menni, 2004 : 79 |
Mylossoma paraguayensis
| Norman, 1929 : 814 |
| MacDonagh & Fuster, 1944 : 355 |
| Géry, 1976 : 50 |
| Oldani, 1983 : 89 |
| Miquelarena, 1986 : 37 |
| Sverlij et al., 1998 : 25 |
| López et al. , 2003 : 34 |
| Menni, 2004 : 79 |
| Britski et al., 2007 : 81 |
Mylosoma [ sic ] aureum
| Bertoni, 1939 : 56 |
Mylossoma duriventre
| Géry, 1976 : 50 |
| Gómez & Chebez, 1996 : 52 |
| Jégu, 2003 : 187 |
| Menni, 2004 : 79 |
| Graça & Pavanelli, 2007 : 82 |
| Langeani et al. , 2007 : 185 |
| Litz & Koerber, 2014 : 9 |
| Mirande & Koerber, 2015 : 15 |
| Koerber et al. , 2017 : 22 |
Mylossoma duriventris orbignyanum
| Géry et al. , 1987 : 443 |
| Gómez & Chebez, 1996 : 52 |
Mylossoma ocellatum
| Ibarra & Stewart, 1987 : 58 |
Mylossoma orbignyanum
| Britski et al. , 2007 : 81 |
Mylossoma orbignyanus
| López et al., 2003 : 34 |
| Menni, 2004 : 79 |